Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2014) Volume 2, Issue 2
The present study was pertaining to study the mechanism and signaling of BRs under high temperature stress in Brassica juncea L. plants. Results advised that high temperature stress reduced the shoot length, root length, fresh weight of the plants but 28-homobrassinosteroids (28-homoBL) treatment reduced the toxic effect of high temperature stress by improving the same of B. juncea L. plants. To ascertain these free radical scavenging enzymes (superoxide dismutase (SOD), Catalase (CAT), Peroxidase (POD), Ascorbate peroxidase (APOX), Polyphenol oxidase (PPO) and Indole acetic acid oxidase (IAAO) and lipid oxidation (MDA) were assayed along with photosynthetic pigments, total carbohydrates, reducing sugars and non-reducing sugars. All studied scavenging enzymes activities were ameliorated by the homoBL treatments. The pigments decreased under temperature stress were also protected by homoBL treatments. Proteins profiling by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDSPAGE) were also studied to know the behavior of proteins under temperature stress as well as with the treatment of homoBL. Protein bands disappeared in temperature stress while in the presence of homoBL treatment existing proteins displayed more accumulation leading to the appearance of new bands. Native-PAGE of SOD, CAT and POD suggested the presence of in isoenzymic forms in cells. Application of different concentration of homoBL increased intensities of these isoenzymes under temperature and normal condition as compared to control untreated seedlings. The gene expression of SOD and CAT also suggested that synthesis these enzymes were decreased under high temperature stress but ameliorated with the treatment of homoBL. Present study demonstrated that 28-homoBL have stress-ameliorative properties in B. juncea seedlings exposed to high temperature stress by improving seedling growth and enhancing protein content as well as activities of SOD, CAT, APOX, PPO and IAA Oxidase. Further, it was suggested by decreasing the MDA content after the treatments of 28-homoBL. This study demonstrate the culmination of BR’s as an anti-stressor for protection of plant exposed to high temperature stress but extensive studies are still needed to know the various aspects related to stress and role of brassinosteroids in regulating them at molecular and signaling level.
Keywords: 28-hmobrassinolide; Brassica juncea L; Reactive oxygen species; High temperature stress
It is predicted that increases in “greenhouse gases” concentration will result in the rise in mean temperatures of about 2°C by the middle of the next century. Although yields of temperate crops increase with enhanced CO2 concentrations, this may be offset by the negative effects of warmer temperatures in determinate crops. The duration of grain filling in crop plants are determined principally by temperature. In winter crop, high temperatures (“31°C) after anthesis can decrease the rate of grain-filling, whilst high temperatures imposed before anthesis can also decrease yield of crop plants (In winter crop, high temperature episodes occurring near to anthesis can reduce the number of grains per ear and the subsequent rate of increase in harvest index, resulting in smaller grain yields. High temperature stress also leads to over production of reactive oxygen species (ROS) in plants which are highly reactive and can cause damage to proteins, lipids, carbohydrates, RNA and DNA ultimately results in death of cell [1]. Plants have evolved mechanisms on morphological, physiological and molecular level to survive with modulated environmental conditions [2].
Various strategies have been employed to enhance plant growth and productivity under environmental stress. Use of phytohormones is one of the simpler approaches to make crops resistible to negative environmental conditions and perform well by acclimatizing at morphological, physiological, biochemical and molecular level. The use of plant growth regulators (PGRS) in agriculture to promote plant growth production and quality is becoming more common [3- 6]. Brassinosteroids is a sixth class of phyto-hormone which plays an advantageous role in enhancing crop productivity under adverse conditions [7,8]. BRs are very peculiar in regulating the activities of antioxidant defense system at biochemical and also regulate the compatible solutes under stress condition in plants. Although studies related to the role of brassinosteroids in controlling the metabolic activities of antioxidant defense system enzymes at biochemical and molecular level are available, a comprehensive study is required to gain a deeper insight of the above mechanism. Therefore present study was pertaining to study the mechanism and signaling of BRs under high temperature stress in B. juncea plants.
28-Homobrassinolide (28-homoBL) was purchased from Sigma-Aldrich Chemicals Pvt. Ltd, India. The stock solution (10-4 M) of 28-homoBL was prepared by dissolving required quantity of 28-homoBL in 5 ml ethanol, in a 100 ml volumetric flask. The desired concentrations (10-6, 10-8 and 10-10 M) of 28-homoBL were prepared by the dilution of stock solution. Seeds of B. juncea L. cv. RLM-619 were procured from the Department of Plant Breeding, Punjab Agriculture University, Ludhiana, India. Healthy seeds of uniform size were surfacesterilized with 0.05% (w/v) Sodium hypochlorite solution followed by repeated washing with deionised water. The surface sterilized seeds were sown in Petri-dish grown in seed germinator with control conditions (16 h Dark, 200 μmol PAR m-2 s-1, 25°C and 80% relative humidity) for 7 days. 5 h temperature treatment (35, 40 and 45°C) was given to 7 days old seedlings for 3 consecutive days and on 10th day seedlings were harvested for various physiological, biochemical and molecular analysis. Morphological data in terms of shoot, root length and fresh weight was measured on 10th days after harvesting.
Determination of carbohydrate, sugar, reducing and reducing sugar content
Carbohydrate content was estimated by the method of Dubois et al. [9]. Reaction mixture contains phenol reagents, H2SO4 and samples were incubated at room temperature for 30 minutes. Absorbance was taken at 485 nm and amount of carbohydrate was expressed as mg g-1 dry weight. Total sugar was estimated following the method of Loewus 1952. Reaction mixture contains 0.05 ml of ethanol extract added 0.95 ml of distilled water and then added 3ml cold anthrone reagent with constant shaking. Reaction mixture was heated for 10 min in boiling water bath and cooled rapidly at room temperature. Absorbance was measured at 630 nm. Amount of total sugars content was calculated and expressed as mg g-1 dry weight. Reducing sugar content was estimated by the method of Nelson [10]. Absorbance was measured at 620 nm expressed as mgg-1 fresh weight. Non reducing sugar was calculated by subtracting the reducing sugars content from the total sugar and expressed as mg g-1 dry weight.
Determination of pigments (Chl a, b and carotenoids)
Total chlorophyll (Chl), chlorophyll a, b and carotenoid (Car) content were quantified by following the method Lichtenthaler et al. Fresh leaves were homogenized in 3.0 ml of 70% acetone followed by its centrifugation at 12000X g for 15 min. Absorbance of supernatant was observed at 645, 663 and 470 nm and 70% acetone was taken as blank. The contents of total chlorophyll, chlorophyll a, chlorophyll b and carotenoid were calculated and presented in terms of mg g-1 fresh weight (Table 1).
| 1. | 28-homoBL | 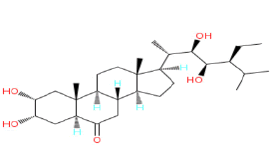 |
478.704 | Sigma Aldrich, USA |
Table 1: Structure, molecular weight of plant steroids.
Determination of vitamin C and proline content
Known weight of fresh seedlings was homogenized in metaphosphoric citrate buffer for quantifying vitamin C using the method of Chinoy et al. [11]. Homogenized extract was centrifuged at 3000X g for 5 min using cooling centrifuge and to the 6 ml supernatant 2 ml of 2, 6-dichlorophenol indophenols was added as dyeing agent. Absorbance was taken at 530 nm immediately after adding dye and amount of vitamin C was calculated using standard growth values in mg g-1 fresh weight. Proline content was estimated spectrometricaly following the method of Bates et al. [12]. 1g fresh leaf tissue was homogenized in 3% sulphosalicylic acid, centrifuged at 12000X g for 10 min. 2 ml supernatant, 2 ml acid ninhydrin and 2 ml glacial acetic acid was added in test tube and reaction was made at 100 C for 1 h. Absorbance of red colored praline-ninhydrin product was measured at 520 nm.
Determination of lipid peroxidation
Lipid Peroxidation was determined by measuring the content of malondialdehyde (MDA) using the method of Heath and Peacker [13]. The sample were homogenized in 10% (w/v) tri-chloroacetic acid and centrifuged at 1000X g for 10 min. supernatants were mixed with equal volume of 10% TCA containing 0.6% (w/v) TBA. Absorbance was taken at 450, 532 and 600 nm after centrifuged at 1000X g for 10 min again.
Determination of oxidative enzymes
Superoxide dismutase (SOD, EC 1.15.1.1) activity was assayed by measuring the ability of the enzymes extract to inhibit the photochemical reduction of nitrobluetetrazolium (NBT) (Kono 1978). For total SOD assay, 3 ml reaction mixture was prepared containing 50 mM sodium carbonate (pH 10.2), 24.1 M NBT, 0.1 mM EDTA, 1 mM hydroxylamine, 0.03% (v/v) Triton X-100 and 70 μl enzymes extract. Absorbance was recorded at 560 nm for 2 min. One unit of SOD activity was defined as the amount of enzyme required that caused 50% of NBT reduction. Activity of ascorbate peroxidase (APOX, EC 1.11.1.11) was determined spectophotometricaly as described by Nakano and Asada [14]. Catalase (CAT, EC 1.11.1.6) activity was assayed by measuring the initial rate of H2O2 disappearance using the method of Aebi [15]. Monodehydroascorbate reductase (MDHAR, EC 1.6.5.4) activity was assayed using the method of Hossain et al. [16]. 3 ml reaction mixture was prepared by using 50 mM Tris-HCl (pH 7.6) contain 2.5 mM Ascorbic acid, 0.1 mM reduced nicotinamide adenine dinucleotide (NADH), 0.14 units of ascorbic acid oxidase and 100 μl of enzyme extract. The enzyme activity was measured by following the decrease in absorbance at 340 nm using extinction coefficient of 6.2 mM- 1cm-1. Dehydroascorbate reductase (DHAR, EC 1.8.5.1) activity was measured following the method given by Dalton et al. [17]. Guaiacol peroxidase (POD, EC 1.11.1.7) activity was assayed using the method of Putter J 1971 with some modifications. 3 ml reaction mixture was made by 50 mM Potassium phosphate buffer (pH 7.0), 20 mM guaiacol, 12.3 mM H2O2 and 100 μl enzyme extract. POD activity was determined by measuring the absorbance at 436 nm and using an extinction coefficient of 26.6 mM-1.cm-1. One unit of POD activity represents the amount of enzyme catalyzing the oxidation of 1 mol of guaiacol in 1 min. PPO (E.C. 1.10.3.1) activity was measured using the method of Bastin and Unleur [18]. The reaction mixture contained 60 mM Phosphate buffer (pH 7.0) and 0.01 M chlorogenic acid and incubated at 30 ± 2°C for one hour. Absorbance was taken at 430 nm and enzyme activity was expressed as mol-1.min-1.mg-1. protein. IAAO (EC. 1.11.1.7) activity was measured according the method of Gordon and Weber [19].
Determination of protein content
Protein estimation was done following the method following the method of Lowry et al. [20].
Native PAGE and enzyme activity staining
Native polyacrylamide gel electrophoresis (PAGE) was performed at 4°C, 180 V, following Laemmli [21]. The enzyme solutions were subjected to native PAGE with 10% polyacrylamide gel SOD for POD and CAT. SDS was omitted from the PAGE. Activity stain for each enzyme was carried out as follows. POD, gel was incubated in 25 mM potassium phosphate buffer (pH 7.0) and then the gel was submerged again in the same buffer (fresh amount) containing 18 mM guaiacol and 25 mM H2O2, until the POD activity-containing band was visualized carefully according to Fielding and Hall [22]. For CAT, gel was incubated in 3.27 mM H2O2 solution for 25 min and then the gel was washed in distilled water and submerged in a solution composed of potassium ferricyanide 1% and ferric chloride 1% for 4 min according to Woodbury et al. [23].
Statistical analysis
All analyses were done on a completely randomized design. All data obtained was subjected to one-way analysis of variance (ANOVA) and the mean differences were compared by Turkey’s test. Each data was the mean of three replicates (n=3) except for shoot lengths of B. juncea L. seedlings (n=10) and comparisons with P-values ≤ 0.05 were considered significantly different. In all the figures, the spread of values is shown as error bars representing standard errors of the mean.
Brassinosteroids stimulate plant growth and development under normal conditions as well as under inadequate conditions of temperature [24]. Present study revealed that temperature stress reduced plant shoot and root growth. Presoaking treatment of BRs made the seedlings more tolerant to abide high temperature shock treatments. Supplementation of BRs ameliorates both shoot and root growth under temperature stress (Figures 1a and 1b). Asthir and Deep [25] also observed similar results in wheat seedlings root and shoot growth. It was found to be inhibited due to heat exposure but applications of homoBL ameliorate growth by mitigating the toxic effect of heat stress. This enhancement was found due to the increased concentrations of ca2+ ions by BRs treatment [26]. It may be possible that there is some cross talk between ca2+ signaling and brassinosteroids at cellular level. ca2+ concentrations in growing parts of the control plants the required signaling cascade under stress as well as with BRs treatments. In present results high temperature stress reduced plant shoot and root growth in B. juncea seedlings.
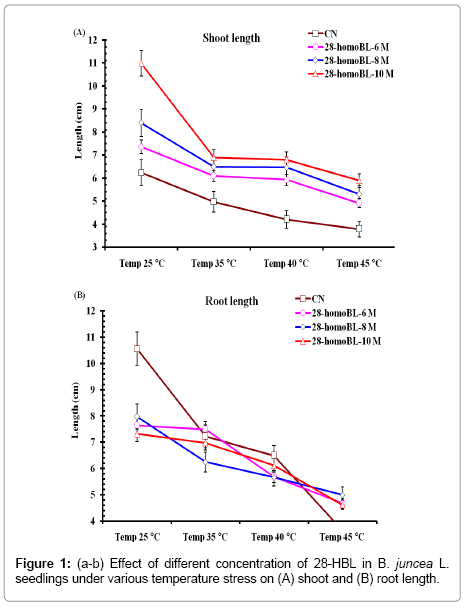
Figure 1: (a-b) Effect of different concentration of 28-HBL in B. juncea L.seedlings under various temperature stress on (A) shoot and (B) root length.
High temperature treatments degraded photosynthetic pigments and BRs treatments provide the protection to the photosynthetic pigments. Among different concentrations of 28-homoBL, 10-8 M homoBL (0.095 ± 0.00029 mg.g-1 fresh weight) was the best for Chlorophyll a at normal room temperature and also in high temperature stress as compared CN untreated seedlings revealed 0.05 ± 0.00023 mg.g-1 fresh weight (Figures 2a-d). Detrimental effect of high temperature on Chl a content 0.01 ± 0.00001 mg.g-1 fresh weights was observed at 45°C. But it was measured 0.053 ± 0.00004 mg.g-1 fresh weight with treatment of homoBL at same temperature. Similarly chl b content decrease 31.76% at 45°C 0.58 ± 0.0026 mg.g-1 fresh weight as compared to CN seedlings (0.85 ± 0.0056 mg.g-1 fresh weights). Opposing decrease in photosynthetic pigments in rape leaves when exposed to low temperature. Present result of seed priming with 28-homoBL help in alleviating the effect of high temperature by protecting the photosynthetic pigments.
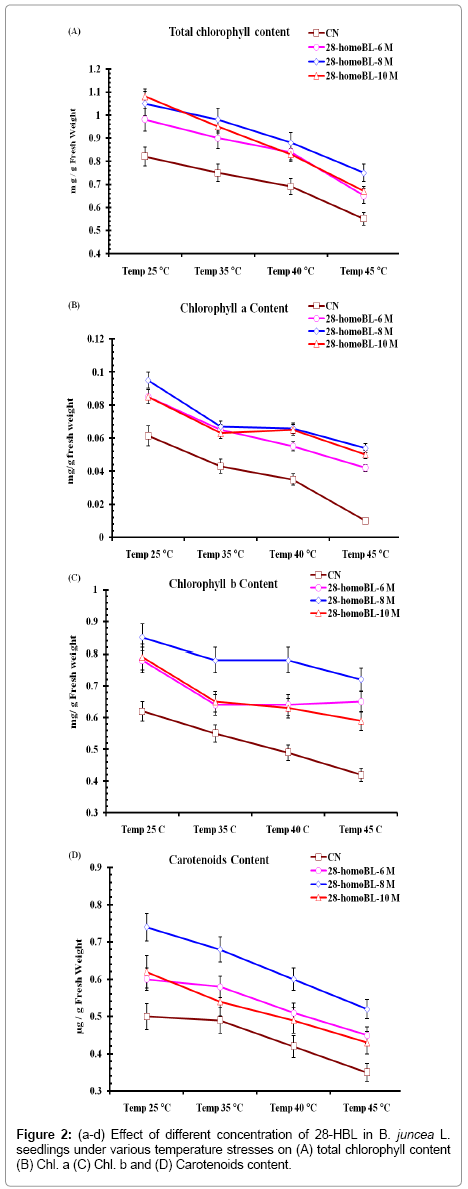
Figure 2: (a-d) Effect of different concentration of 28-HBL in B. juncea L. seedlings under various temperature stresses on (A) total chlorophyll content (B) Chl. a (C) Chl. b and (D) Carotenoids content.
In present study total carbohydrates, total soluble sugar; reducing and non-reducing sugar ameliorated under extreme condition of temperature (Figure 3a-d). Results suggested that with increase in temperature from 35 to 45°C, total carbohydrates accumulated in BRs treated and untreated CN seedlings. It was further ameliorated in the seedlings supplemented with different concentration of homoBL. It was 21.64% more in 10-8M homoBL treatment as compared to CN untreated seedlings exposed to 40°C temperatures stress. Present results also revealed that the seeds primed with different concentrations of homoBL and then exposed at to high temperature showed more accumulation of carbon compounds although photosynthetic degradation was still at higher rate. Carbohydrates, total soluble sugar, reducing and nonreducing sugars are primary source of energy and carbohydrates also accumulates under stress full conditions in plants to act as osmoprotectent [27]. It also due to loss of chlorophyll and carotenoids and due oxidation under high temperature of these pigments which decreases the metabolic rate leads to reduced carbon metabolism. Reported that BR27 decreased sugar content in winter oilseed rape callus.
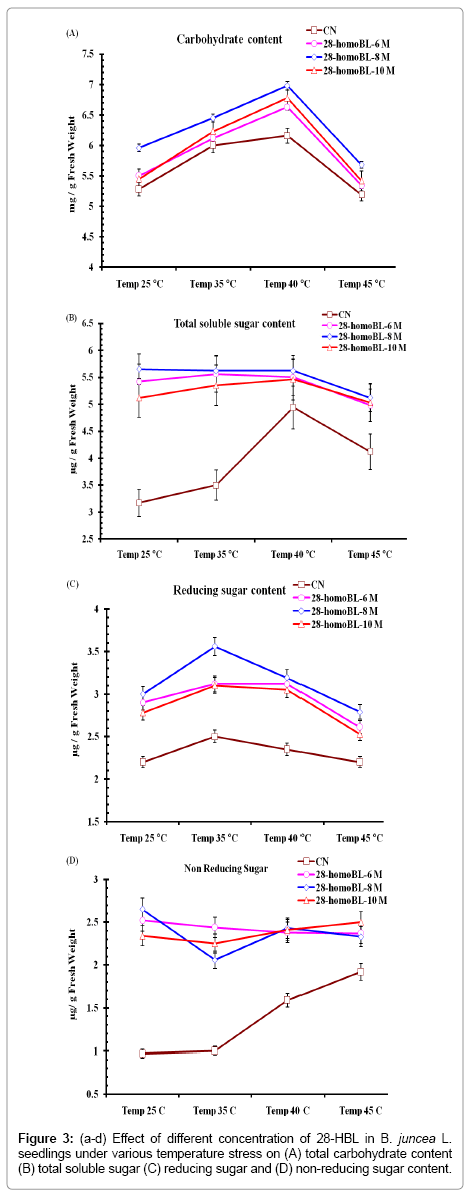
Figure 3: (a-d) Effect of different concentration of 28-HBL in B. juncea L.seedlings under various temperature stress on (A) total carbohydrate content (B) total soluble sugar (C) reducing sugar and (D) non-reducing sugar content.
Ascorbic acid content also increase significantly in 10-10 M concentration of homoBL treated seedlings as compared to control seedlings and after the high temperature treatment it was further increase with homo BL treatments (Figure 4a). This was 40.50% more than respective control untreated seedlings. 28-homoBL also proved to be beneficial in ameliorating AsA at all temperatures. In homoBL treated seedlings, 10-8 M concentration was best for AsA (13.51 ± 2.74 μg g-1 FW) content at 40°C. Whereas CN seedlings had 5.5 ± 0.99 μg.g-1 FW AsA content. This amelioration was 59.28% more than the CN untreated seedlings[28]. In their experiment on Lycopersicom esculentum L. reputed exogenous application of BR27 alters the ascorbic acid under normal conditions. Significant effect of temperature stress and 28-homoBL was observed on proline accumulation in B. juncea L. seedlings. The amelioration of proline content was 61.68% high than CN (Figure 4b). Proline content in higher plants increases under different environmental stress as observes in H2O2, drought, high salinity [29] high light and UV radiation [30], heavy metals [31] and oxidative stress [32]. Present studies suggested that under high temperature proline content increased to assertively which further ameliorated when seedlings grown with 28-homoBL primed seeds [33]. Indicated the ameliorative effect of 24-epiBL and 28-homoBL in Cd-stress alleviation which was associated with enhanced levels of proline. Under high temperature stress accumulation of proline strongly protects B. juncea seedlings from toxic effect of ROS produces in plant cells. Under high temperature which may represent a secondary defensive mechanism against oxidative stress of high temperature.
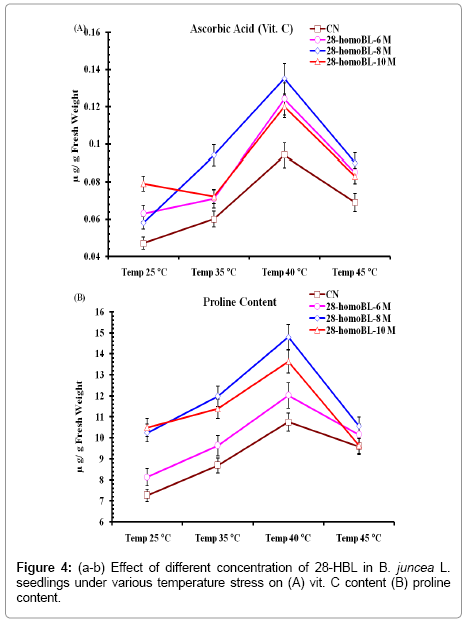
Figure 4: (a-b) Effect of different concentration of 28-HBL in B. juncea L. seedlings under various temperature stress on (A) vit. C content (B) proline content.
Total protein is the most sensitive component of structure and storage start degradation during in adequate conditions. Total protein under present study significantly decreased under present high temperature stress. Seed primed with 28-homoBL when exposed to high temperature found to be more tolerant in terms of their protein degradation rate. Seedlings having only 28-homoBL pre-sowing treatment grow under normal temperature conditions decisively increased their protein content (Figure 5a). While studying Brassinosteroids functions to protect the translational machinery and heat shock protein synthesis in B. napus L. following thermal stress. It found that treatment of B. napus L. seedlings with 24-epiBL leads to a significant increase in their basic growth and thermo-tolerance properties. EBR treatment resulted higher accumulation of four major classes of heat shock protein (Hsps) as compared to untreated seedlings.
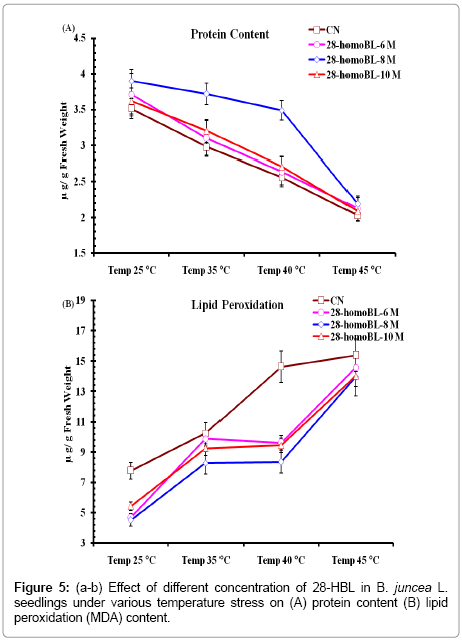
Figure 5: (a-b) Effect of different concentration of 28-HBL in B. juncea L. seedlings under various temperature stress on (A) protein content (B) lipid peroxidation (MDA) content.
Thermo-tolerance in plants is mediated via accumulation of osmoprotectant like proline and vitamins along with phenolic compounds and auxins also found to involved in such tolerance mechanisms. However higher temperature stress generates high level of ROS which causes oxidative damage leading to membrane destruction which in terms affects the levels MDA content and antioxidant enzymes [7,8]. Over production of MDA content leads to oxidative degradation of phospholipids of membranes, ensuing in disruption of the permeability of cell membrane. However, it is feasible that treatment of brassinosteroids contribute in inhibition of lipid degradation as well as stimulation of activities of antioxidant enzymes under stress. In present study 28-HBL, enhances thermo-tolerance generated by high temperature exposure has been observed. This aligns with the prior reports of Sirhindi et al. [34] in B. juncea under various temperature stresses. Alteration of MDA content may suggest that 28-HBL treated seeds were less affected by thermal stress than the untreated control (7.8 ± 0.26) in B. junceaseedlings. The maximum increase in MDA content was observed at 45C (15.14 ± 1.53) and this increase in MDA content was reduced to significant low level by 28-HBL seed priming (Figure 5b). The maximum reduction in MDA content was recorded in seedlings treated with 10-8 M HBL under 40°C temperature tress (8.29 ± 1.35). It was also observed that seedlings treated with 28-HBL alone could lower MDA level then control untreated seedlings at normal temperature (7.8 ± 0.26).
Thermal stress induces production of phenolic compounds, such as flavonoids and phenyl propanoids [33,35,36]. Synergistic effect of high temperature and 28-HBL was found in Brassica juncea seedlings Polyphenol oxidase activities (PPO). It was found that seedlings having pre-sowing soaking treatment of 28-homoBL increase their PPO activity to further enhanced levels then control (7.54 ± 1.62). The maximum increase in PPO activity was observed at (14.83 ± 2.31) and this increased in PPO was further enhanced significantly by 28-homoBL seed priming treatment. The maximum activity of PPO was observed in seedlings raised from seeds primed with 10-8 28-HBL at 40°C. It was also observed that 28-HBL alone under normal temperature help in ameliorating PPO activity in treated seedlings than control untreated seedlings (7.54 ± 1.62)) (Figure 6a) [37] and [38] reported the activity PPO increases in response to deferent types of biotic and abiotic stress [39]. Reported accumulation of soluble phenolic compound but lower activities of Peroxidase and PPO activity in tomato and watermelon plants, which indicated that thermal stress induces accumulation of phenolic in plants by activating their biosynthesis as well as inhibiting their oxidation. Related PPO activity to the appearance of physiological injuries caused by thermal stress in Mahoniarepens plants. The specific activity of IAA oxidase in seedlings of B. juncea L. exposed to high temperatures was observed in present study. This Increase was further ameliorated in seedlings raised under normal temperature after seeds treated with different concentration of 28-homoBL. 10-8 M homoBL was found to be best in present case study.
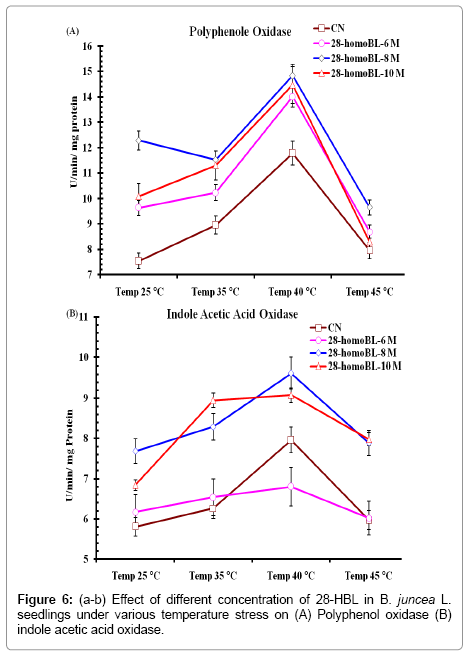
Figure 6: (a-b) Effect of different concentration of 28-HBL in B. juncea L. seedlings under various temperature stress on (A) Polyphenol oxidase (B) indole acetic acid oxidase.
The enzymatic activity sharply increased during temperature exposure raised from 35 to 40°C, and its activity but detrimental decrease when temperature rose to 45°C (Figure 6b). 28-HBL treatment further ameliorated IAAO activity under normal and thermal stressed seedlings concluded that there must be some cross talk at signaling level between IAAO and HBL and 28-HBL might be regulated IAAO activity for various growth and developmental mechanisms under normal temperature and further enhanced under thermal stress conditions [40] and [7] studied IAAO activity in some forest tree and B. juncea, respectively under biotic stress and found that IAAO activity increased under such stress conditions. These results confirmed our previous results of 24-eBL regulation thermal stress in B. juncea L. [8].
The studies carried out on antioxidant enzyme activities in B. juncea L. seedlings exposed to different high temperature conditions revealed the considerable effect of 28-homoBL treatments. In present study activity of SOD increased significantly under temperature stress and maximum activity was observed at 40°C (34.88 ± 3.56). However, when temperature increased to 45°C, SOD activity decreased to lowest level (19.22 ± 1.23). Seed priming with different concentration of 28-HBL alone significantly alleviate the levels of SOD activity under normal temperature conditions. Our previous reports also showed that exogenous application of BRs modulate antioxidant enzyme activity in normal and stressed Brassica juncea plants [36,40]. Maximum activity of SOD was reported in Indian mustard seedlings treated with 10-8M 28-HBL exposed to 40°C temperature shock (34.88 ± 3.56) as compared to only 10-8 M 28-HBL or temperature (Figure 7a).
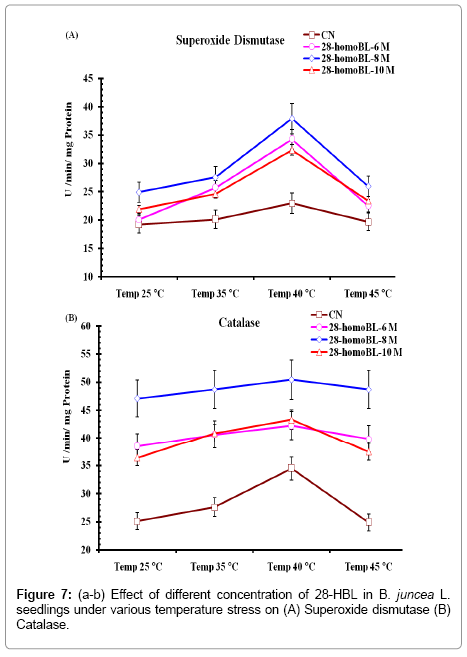
Figure 7: (a-b) Effect of different concentration of 28-HBL in B. juncea L. seedlings under various temperature stress on (A) Superoxide dismutase (B) Catalase.
Activity of CAT showed sharp increase in its activity in seedlings exposed to high temperatures and maximum activity was found at 40°C in distilled water seedlings. However, in seedlings raised from seeds primed with different concentrations showed significant high level of CAT enzyme activity under normal and high temperature conditions. Maximum CAT activity was observed at 10-8 M 28-HBL treated seedlings at 40°C (50.51 ± 4.56). Very smooth increase in CAT enzyme activity was observed in seedlings treated with 10-8 M homoBL with increase in temperatures from 25-45°C which otherwise sharp and dynamic in other treated seedlings at different temperature (Figure 7b). Lipid metabolism in peroxisomes produced H2O2 on which CAT acts to scavenge it into o2 and H2o. Under stress conditions this ROS (H2O2) produced at high rate which has to be detoxified and for this CAT enzyme activity increase. In present study it was observed that under normal conditions of growth and development presence of 28- HBL decisively increase CAT activity. This might be due to the reason that 28-HBL increases lipid metabolism which ultimately leads to more productivity of vegetative oil contents. Activities of APOX and POD were increased in seedlings exposed to high temperature alone. Maximum APOX (value) and POD (value) activity were observed at 40°C when seedlings were raised in distilled water only. Supplementation of 28- HBL at pre-sowing seed level enhanced activities of APOX and POD decisive level. Maximum APOX activity was found in 10-8 M HBL treated seeds at 40°C (26.12 ± 2.34). Similarly POD activity was also maximum in 10-8 M HBL treated seeds at 40°C (17.77 ± 1.89) (Figure 8a-b). [7] Reported in radish seedlings increased activity of APOX in 28-HBL treated seeds under normal and metal stress conditions. In some work they reported decrease in POD activity in presence of 28-HBL alone or supplemented with Cr stress. Both APOX and Pod are involved in removal of H2O2 produced during metabolism in the cytoplasm using different substrate to detoxify H2O2 produced during metabolism in the cytoplasm using different substrates. APOX use ascorbic acid as substrate to detoxify H2O2 by reduced itself to dehydroascorbate and then monodehydroascorbate, which ultimately manage and able to maintained the altered redox status of cells under normal conditions [41]. Established that ascorbic acid in its reduced form in essential for ROS scavenging and its cellular pool is maintained by DHAR and MDAR using NADPH as reducing power. From present findings we try to establish that 28-HBL possibly maintained the altered redox status of cells b regulating activities of antioxidant enzymes viz SOD, CAT, APOX, GR, DHAR, MDHAR etc. 28-HBL might be able to re-established redox potential by reducing lipid peroxidation of phospholipids in cell membrane and accumulation of certain osmoprotectants like proline.
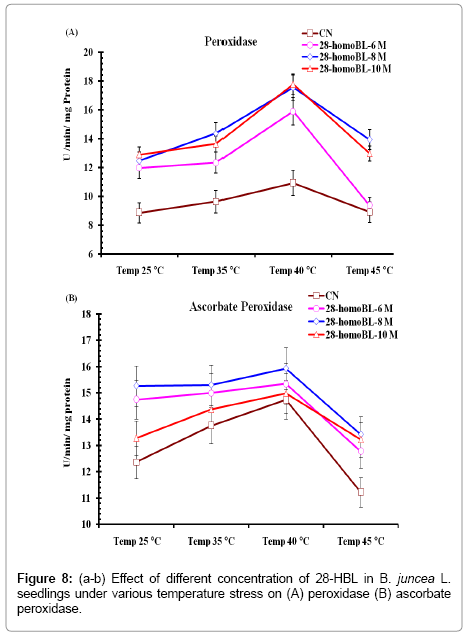
Figure 8: (a-b) Effect of different concentration of 28-HBL in B. juncea L. seedlings under various temperature stress on (A) peroxidase (B) ascorbate peroxidase.
Activities of DHAR and MDHAR decisively increased from (DHAR value) and (MDHAR value) respectively under control untreated seedlings to (DHAR) and value (MDHAR) in 28-HBL treated seedlings. Under temperature treatments DHAR and MDHAR activities eventually increased and maximum DHAR activity was observed in 10-8 M HBL treated seedlings at 40°C (Figure 9a-b).
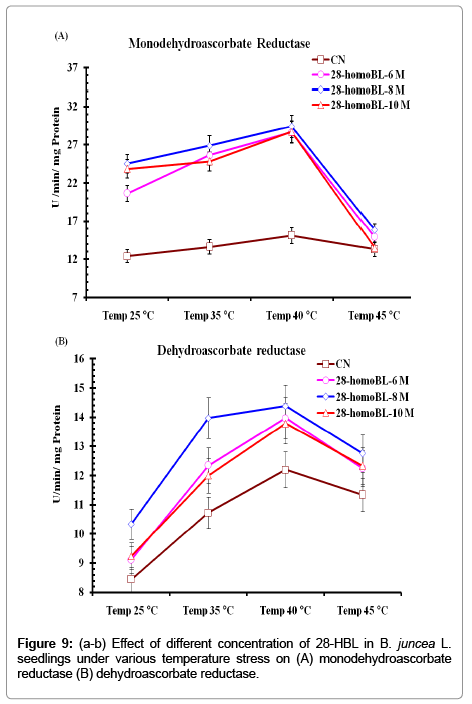
Figure 9: (a-b) Effect of different concentration of 28-HBL in B. juncea L. seedlings under various temperature stress on (A) monodehydroascorbate reductase (B) dehydroascorbate reductase.
Overproduction of ROS during thermal stress is responsible for oxidative degradation of cell membrane which is confirmed from higher values of MDA content in seedlings exposed to high temperature. Seed priming treatments with 28-HBL helped in reducing this lipid peroxidation to significantly low levels as compared to untreated seedlings under various temperature treatments. It might be due to the reason that BRs which are modified sterols can able to modulate the activity of proteins and other antioxidant enzymes within the membrane by affecting either protein confirmation (functionally) or protein activity by direct protein -sterol interactions under thermal stress.
To analyses of rates of SOD, POD, APOX, CAT, DHAR and MDHAR revealed that temperature stress ameliorate internal immune metabolism of plant to produce certain level of high resistance. Seed priming treatment with 28-homoBL enhanced this thermo-tolerance level to further heights. High temperature resulted in high production of ROS, the toxic effect of which was mitigated up to certain level by antioxidant enzyme system which scavenge these free radicals. When seeds prior to the temperature exposure were primed with different concentrations of 28-homoBL, scavenging activity of these antioxidant increased. Under temperature stress the cellular homeostasis disturb due to high rate of ROS production than their mitigation by various antioxidant enzymes. High radicals (o2 -), hydrogen peroxide (H2O2), hydroxyl radicals (HO-), peroxy radical (ROO), etc. [42]. These reactive species are symptoms of cellular injury due to high temperature [43]. High production of ROS causes autocatalytic peroxidation of membrane lipids and pigments thus leading to the loss of membrane semi-permeability and modifying its functions [43]. Thus, under the influence of enhanced ROS production, plant can maintain its ROSscavenging antioxidant enzymes such as SOD, CAT, APOX, POD, DHAR and MDHAR etc. to higher levels to detoxify the ROS toxicity [41]. Under thermal stress well regulated more efficient antioxidant system was studied by member of workers [44,45] Recently, studies available showed that different molecular signaling such as BR, abscisic acid (ABA) ca2+ calmodulin (CaM), hydrogen peroxide (H2O2)mitogenactivated protein kinase (MAPK) cascade and nitric oxide (NO) are involved in regulation of plant responses to various environmental stresses including heat stress [42,46] (Figure 10).
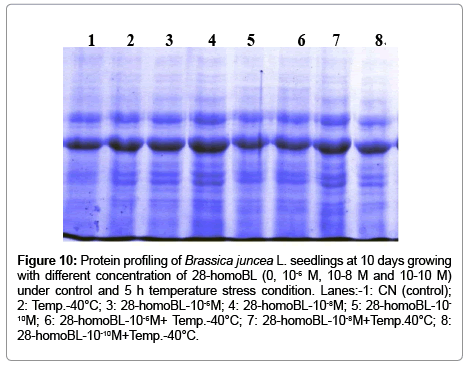
Figure 10: Protein profiling of Brassica juncea L. seedlings at 10 days growing with different concentration of 28-homoBL (0, 10-6M, 10-8 M and 10-10 M) under control and 5 h temperature stress condition. Lanes:-1: CN (control); 2: Temp.-40°C; 3: 28-homoBL-10-6M; 4: 28-homoBL-10-8M; 5: 28-homoBL-10-10M; 6: 28-homoBL-10-6M+ Temp.-40°C; 7: 28-homoBL-10-8M+Temp.40°C; 8: 28-homoBL-10-10M+Temp.-40°C.
In part of the native polyacrylamide gel electrophorasis (PAGE) analysis, three isozymes bands of SOD i.e. Mn-SOD, Fe-SOD and Cu/ Zn-SOD were detected (Figure 11). But the intensities of these bands were increased under temperature stress as well as with brassinosteroids treatments which is again confirmation of biochemical results of total SOD activities. In addition, three bands were also detected for CAT isozymes, which showed higher intensity under high temperature in brassinosteroids treated seedlings (Figure 12). Native polyacrylamide gel electrophoresis (PAGE) analysis for Peroxidase (POD) enzyme showed three isozymes which also showed increase in their intensities under temperature treatments supplemented with BRs (Figure 13). Similar result was observed by [28] in Lycopersicon esculentum under drought stress. SDS-PAGE also revealed that protein was degraded under temperature stress and ameliorated by brassinosteroids treatments under normal as well as under temperature stress condition in B. juncea L. seedlings.
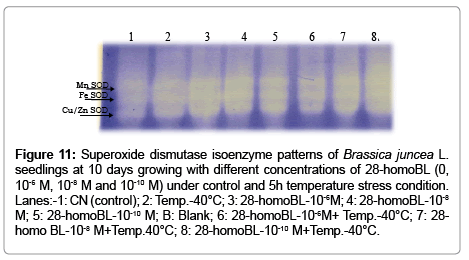
Figure 11: Superoxide dismutase isoenzyme patterns of Brassica juncea L. seedlings at 10 days growing with different concentrations of 28-homoBL (0, 10-6 M, 10-8 M and 10-10 M) under control and 5h temperature stress condition. Lanes:-1: CN (control); 2: Temp.-40°C; 3: 28-homoBL-10-6M; 4: 28-homoBL-10-8 M; 5: 28-homoBL-10-10 M; B: Blank; 6: 28-homoBL-10-6M+ Temp.-40°C; 7: 28- homo BL-10-8 M+Temp.40°C; 8: 28-homoBL-10-10 M+Temp.-40°C.
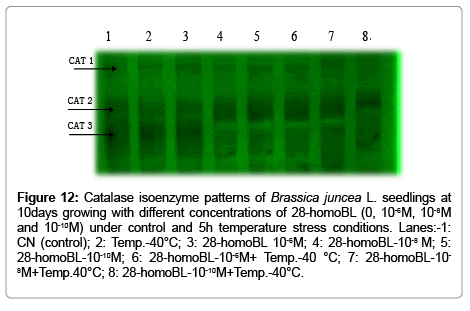
Figure 12: Catalase isoenzyme patterns of Brassica juncea L. seedlings at 10days growing with different concentrations of 28-homoBL (0, 10-6M, 10-8M and 10-10M) under control and 5h temperature stress conditions. Lanes:-1: CN (control); 2: Temp.-40°C; 3: 28-homoBL 10-6M; 4: 28-homoBL-10-8 M; 5: 28-homoBL-10-10M; 6: 28-homoBL-10-6M+ Temp.-40 °C; 7: 28-homoBL-10-8M+Temp.40°C; 8: 28-homoBL-10-10M+Temp.-40°C.

Figure 13: Peroxidase isoenzyme patterns of Brassica juncea L. seedlings at 10 days growing with different concentrations of 28-homoBL (0, 10-6M, 10- 8M and 10-10M) under control and 5h temperature stress conditions. Lanes:-1: CN (control); 2: Temp.-40°C; 3: 28-homoBL 10-6M; 4: 28-homoBL-10-8M; 5: 28-homoBL-10-10M; 6: 28-homoBL-10-6M+ Temp.-40°C; 7: 28-homoBL-10- 8M+Temp.40°C; 8: 28-homoBL-10-10M+Temp.-40°C.
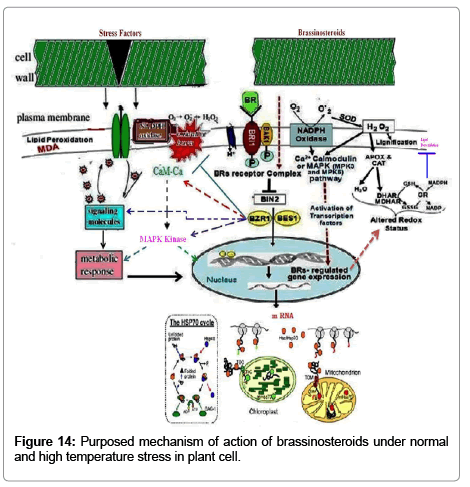
Figure 14: Purposed mechanism of action of brassinosteroids under normal and high temperature stress in plant cell.
High temperature stress indicated detrimental effects on several key physiological processes such as photosynthetic pigments, carbon metabolism, lipid peroxidation and antioxidant defense system. The main toxic effect of high temperature appeared to be on photosynthesis, degradation of proteins, increased lipid peroxidation. Supplementations of 28-homoBL treatment improve seedling/plant growth by protecting the proteins from thermal degradation by ameliorating the free radical scavenging system. BRs are present in plants accountable for pleiotropic effects in plants under normal and stress environment. A probable mechanism has been proposed based on the information of BRs mechanism available in literature and results obtained in present study. This study explores another role of BRs as an anti-stressor for protection of plant from thermal stress by modulating the metabolic machinery of plants.
Financial assistance from University Grants Commission, New Delhi is duly acknowledged.