Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2013) Volume 4, Issue 1
The aim of this study was to discuss a common concept of population dynamics applicable to both Thrips imaginis (Thysanoptera) and the Pacific stock of the Japanese sardine (Sardinops melanostictus). First, I elucidate the mechanism that produces the false density-dependent effect on population changes of Thrips imaginis, I conducted simple deterministic simulations to discuss the qualitative viewpoints. Second, I conducted Monte Carlo simulations by using the average population size and standard deviation of the thrip data used by Davidson and Andrewartha for the quantitative discussion. In simple deterministic simulations, the resultant plots of population change against population size showed a decreasing trend for which the slope was statistically significant even though the true relationship between the population change and population size had no density-dependent effect. The results of the Monte Carlo simulations indicated that nearly 70% of the trials showed false density-dependent effects. The provability of the false decision, which was to recognize the existence of density-dependent effects, increased as the standard deviation in population size in a month i-1 increased. When the number of samples increased, the probability of the false decision greatly increased. The conclusion from these simulations was that the density-dependent effect that emerged in the population change of T. imaginis was artificially produced and invalid. Further, the thrip population size in month i was determined in proportion to that in month i-1; and environmental conditions in month i. This mechanism was completely same of that shown in the Japanese sardine (Sardinops melanostictus). Therefore, the fluctuations in populations of T. imaginis and Japanese sardine could be explained with the same concept of population dynamics shown here.
Keywords: Thrips imagines, Density-Dependent Effect, Population Change, Reproductive Success, Japanese Sardine
When discussing a fluctuation mechanism in biotic resources, one of the key issues is whether a density-dependent effect exists. In fisheries science, nearly all scientists, at one time, have believed in the existence of a density-dependent effect and have discussed management schemes under the assumption that a density-dependent effect plays an important role in controlling population fluctuations. In other words, if a density-dependent effect truly exists in the stock-recruitment relationship, which is the relationship between recruitment and spawning-stock biomass, or is a key factor in controlling population fluctuations, then an optimal population size that achieves the maximum sustainable yield (MSY) can be defined, which is one of the most important concepts in fisheries management. The basic idea of fisheries management is essentially constructed by this concept, which has been widely used all over the world [1,2]. However, if a densitydependent effect does not exist or is nearly negligible in explaining population fluctuations, then an MSY cannot be defined, and the optimal population size could never be determined biologically. Therefore, depending on whether a density-dependent effect exits or is negligible, the management procedure will differ. The concept of “regime shift,” which is large, abrupt, and persistent changes in the ocean ecosystem, has recently become popular [3]. By using the concept of regime shift, Wada and Jacobson [4] analyzed Japanese sardines and concluded that density-dependent effects existed in the favorable and unfavorable regimes, and the carrying capacities during the favorable regime were 75 times greater than those during the unfavorable regime. Wada and Jacobson’s [4] work has been cited in many paper [5-9] and is truly a key paper in the analysis of Sardinips population dynamics. However, Sakuramoto [10] reanalyzed the data used by Wada and Jacobson [4] and showed diametrically opposed results. That is, a false decreasing trend was produced in the regression line of the natural logarithm for reproductive success (recruitment divided by egg production) against the natural logarithm for egg production in response to the observation error, and thus, the density-dependent effect detected was artificial and invalid. Sakuramoto [10] concluded that the recruitment for the Japanese sardine was basically determined in proportion to the spawning stock biomass, then a proportional model was reasonable to accept as the optimal stock-recruitment relationship model.
Sakuramoto and Suzuki [11] showed the mechanism that the density-dependent effect was erroneously detected in response to the observed and/or process errors in the variables. In particular, the effect was large when the errors in the independent variables increased. Further, Sakuramoto and Suzuki [11] specifically proposed that an opposite result was not possible. That is, when the true model showed a density-dependent effect, such as that expressed by the Ricker [12] or Beverton and Holt model [13], the proportional model was seldom selected in response to process and/or observation errors. This suggests that if a proportional model is identified for the actual data, then it does not erroneously produce results in response to process and/or observation errors.
I was surprised to recently learn of the existence of a similar polemic on the density-dependent effect in the field of entomology, more than 50 years ago. The apple blossom thrips (Thrips imaginis Bagnall) is widely distributed in Australia, where is an indigenous species [1]. The females do not lay eggs in batches, but lay several eggs each day continuously throughout their adult life; and an adult may live for several months at the temperatures experienced during the spring [14]. The population at any time includes individuals of various stages and ages; the generation effects is therefore not likely to be pronounces [15]. During the years 1932-8 the number of T. imaginis in 20 roses in the garden at the White Institute was recorded throughout the year. Since 1938 samples have been taken only during the spring and early summer (September to December) [16].
By using these data, Davidson and Andrewartha [1,16] showed that the population size of T. imaginis can be predicted accurately based on the data regarding the temperature and precipitation, and they concluded that the population size could be explained by a succession of good and bad seasons and by the heterogeneity of places where they live [17]. That is, they denied the conventional understanding that a natural population can be regulated only by density-dependent factors; however, Frederick Smith [18] reanalyzed Davidson and Andrewartha’s data to test whether changes in the thrip population size also showed signs of density-dependent effects. That is, whether the changes in the thrip population size showed a negative trend against the population size. I believe that the polemic expressed regarding the thrip population size is essentially the same problem discussed in fisheries researches. The issue discussed in the field of entomology will be resolved using the same logic as that proposed by Sakuramoto [10]. The aim of this study is to elucidate the mechanism that produces the false densitydependent effect in population changes of T. imaginis and to show the mechanisms in population fluctuations of T. imaginis can be explained with essentially the same mechanisms in population fluctuations of Japanese sardine.
Simple deterministic simulations
First, in order to show the mechanism that produces the false density-dependent effect in population changes, simple deterministic simulations were conducted according to Sakuramoto [10] as follows: N1 and N2 denote the average thrip population size in months t1 and t2, respectively. For simplicity, I considered the case when N1 and N2 independently fluctuated in response to environmental conditions in month t1 and t2. Only 3 environmental conditions were assumed: bad, moderate, and good. When the condition was bad, the population deceased at half the average rate, and when the condition was good, the population increased by 2 times the average rate. When the condition was moderate, the population equaled the average size. When the number of thrips in t1 and t2 were independently determined by environmental conditions in the months studied, 9 pairs of population sizes were obtained (n1,i, n2,i) (i=1, 2,…9), where n1,i, n2,i denote the observed thrip population size in months t1 and t2, respectively. The population changes during these 2 months were defined by using the following logarithm: log(dni) = log(n2,i) - log(n1,i). That is, 9 pairs of log(n1,i) and log(dni) (i=1, 2,…9) were obtained. Table 1 shows the 3 cases of (N1, N2 ). That is, N1 was set at 1000 for all 3 cases; however, N2 was set at 2000 (increasing pattern), 1000 (no different pattern) and 500 (decreasing pattern) respectively.
| Deterministic simulation 1 N 2 = 2000 | |||||
|---|---|---|---|---|---|
| Increasing pattern | n 2,b | n2,m | n2,g | ||
| 1000 | 2000 | 4000 | |||
| N1=1000 | n1,b | 500 | (n1,dn1)=(500, 2) | (n1, dn1)=(500, 4) | (n1, dn1)=(500, 8) |
| n1,m | 1000 | (n1, dn1)=(1000, 1) | (n1, dn1)=(1000, 2) | (n1, dn1)=(1000, 4) | |
| n1,g | 2000 | (n1, dn1)=(2000, 0.5) | (n1, dn 1)=(2000, 1) | (n1, dn1)=(2000, 2) | |
| Deterministic simulation 2 N 2 = 1000 | |||||
| No different pattern | n2,b | n2,m | n 2,g | ||
| 500 | 1000 | 2000 | |||
| N1=1000 | n1,b | 500 | (n1, dn1)=(500, 1) | (n1, dn1)=(500, 2) | (n1, dn1)=(500, 4) |
| n1,m | 1000 | (n1 dn1)=(1000, 0.5) | (n1, dn1)=(1000, 1) | (n1, dn1)=(1000, 2) | |
| n1,g | 2000 | (n1, dn1)=(2000, 0.25) | (n1, dn1)=(2000, 0.5) | (n1, dn1)=(2000, 1) | |
| Deterministic simulation 3 N 2 = 500 | |||||
| Decreasing pattern | n 2,b | n 2,m | n2,g | ||
| 250 | 500 | 1000 | |||
| N1=1000 | n1,b | 500 | (n1, dn1)=(500, 0.5) | (n 1, dn 1)=(500, 1) | (n1, dn1)=(500, 2) |
| n1,m | 1000 | (n1, dn1)=(1000, 0.25) | (n 1, dn 1)=(1000, 0.5) | (n1, dn1)=(1000, 1) | |
| n1,g | 2000 | (n1, dn1)=(2000, 0.125) | (n 1, dn 1)=(2000, 0.25) | (n1, dn1)=(2000, 0.5) | |
Table 1: Population sizes in month t1 and t2 assumed in the simple deterministic simulations. (n1, dn1) denotes the observed pairs of population size and population change determined by each environmental condition. ni,b, ni,m, and ni,g, (i=1,2) denote bad, moderate and good environmental conditions, respectively.
Monte Carlo simulations
Second, in order to check the probability that produces the false density-dependent effect in population changes of T. imaginis, Monte Carlo simulations were conducted by using the data for thrip populations listed in Table 3 of Davidson and Andrewartha [1] (Table 2). In accordance with Smith [18], I first focused on the data in October and November. According to the period of data from 1932 to 1938, I randomly generated 7 pairs of artificial data, log(n1,i) and log(n2,i) from the normal distribution N(moct,sd2Oct) and N(mNov, sd2Nov) respectively, where mOct, sdOct and mNov, sdNov were the means and standard deviations of the observed logarithm transformed thrip data by October and November, respectively. Monte Carlo simulations were conducted 1000 times and the slopes of the regression lines of log(dni) were calculated against log(n1,i) (i = 1, 2,…,7). Then, I counted the number of trials in which the slope was significantly negative.
| 1932-33 | 1933-34 | 1934-35 | 1935-36 | 1936-37 | 1937-38 | 1938- | Mean | Standard deviation | |
|---|---|---|---|---|---|---|---|---|---|
| Apr. | 0.65 | 0.38 | 0.79 | 0.41 | 0.52 | 0.30 | 0.54 | 0.51 | 0.17 |
| May | 1.37 | 0.59 | 1.16 | 0.79 | 0.91 | 0.48 | 1.04 | 0.91 | 0.31 |
| Jun. | 1.25 | 0.72 | 1.43 | 1.22 | 1.11 | 0.68 | 0.75 | 1.02 | 0.30 |
| Jul. | 0.64 | 0.74 | 0.83 | -0.10 | 0.96 | 0.40 | 0.90 | 0.62 | 0.37 |
| Aug | 0.52 | 0.23 | 0.57 | 0.23 | 0.26 | 0.23 | 0.73 | 0.40 | 0.21 |
| Sep. | 1.53 | 0.45 | 0.77 | 0.74 | 0.63 | 0.66 | 1.71 | 0.93 | 0.49 |
| Oct. | 1.14 | 1.00 | 0.20 | 1.34 | 1.05 | 1.39 | 2.20 | 1.19 | 0.59 |
| Nov. | 2.13 | 1.89 | 1.56 | 2.13 | 1.61 | 2.49 | 2.76 | 2.08 | 0.44 |
| Dec. | 2.43 | 1.85 | 1.88 | 2.14 | 1.84 | 2.11 | 2.14 | 2.05 | 0.21 |
| Jan. | 1.58 | 1.19 | 1.16 | 1.20 | 0.77 | 1.19 | 1.18 | 0.26 | |
| Feb. | 0.99 | 0.71 | 0.89 | 0.76 | 0.46 | 0.79 | 0.77 | 0.18 | |
| Mar | 0.45 | 0.86 | 0.57 | 0.80 | 0.57 | 0.60 | 0.64 | 0.16 |
Table 2: The data used by Davidson and Andrewartha [1] transformed by logarithm.
Sensitivity tests
The sensitivity tests for the number of trials in which the slope was significantly negative were conducted when the standard deviations changed in log(n1,i) and/or log(n2,i). In addition, the tests were conducted when the number of observed samples changed from 7 to 14.
Reanalysis of the data by Davidson and Andrewartha [1]
In accordance with Smith [18], I first reanalyzed the October and November data by Davidson and Andrewartha [1]. Second, I analyzed the relationships between the population size in month i and that in month i-1 for 7 years (i=May, Jun,...,Sep., and Dec.) and 6 years (i=Dec,..., Feb.) respectively. I also analyzed the relationship using all the data from April 1932 to December 1938. I used 2 regression methods for plotting log(n2,i) against log(n1,i) (i.e., single regression analysis and Deming regression analysis [19]. A single regression analysis is problematic in that it assumes no observation error of the independent variable; therefore, parameter estimates derived from a single regression analysis exhibit bias [20]. When both independent and dependent variables have observational errors, the Deming regression analysis [19] is effective to remove the bias derived from a single regression analysis for the case when the sample size is enough large. The estimation using the Deming regression was conducted using the program developed by Aoki [21].
Comparison of the model proposed by Davidson and Andrewartha [1] and that proposed in this paper
If the population size in month i is determined in proportion to that in month i-1 and the effect of the environmental conditions in month i multiplicatively add to the population size in month i, the relationship of population size between the 2 months can be expressed as follows:
ni=a·ni-1·f(Xi), (1)
Where a and f(.) denote a proportional constant and functions that determine the effects in response to environmental conditions Xi-=[xi,1,...,xi,k] (k denotes the number of environmental factors), respectively. When n0 denotes the first month or reference month, ni can be written as follows:
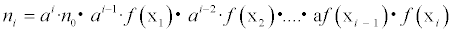 (2)
(2)
This indicates that the population size in month i can be expressed using the population size in reference to month 0 and the environmental conditions of the reference month over that of month i. That is, the difference of population size in month i from the reference month 0 can be explained only by the environmental factors, x1,...,xi. On the other hand, if equation (2) is correct, then the following relationship between month i and month i–1 can be obtained:
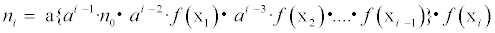 (3)
(3)
Davidson and Andrewartha [16] used the partial regression analysis to explain the population fluctuation in T. imaginis and noted in the summary of their paper as follows: The method of partial regression was used to measure the degree of association between the number of thrips present during the spring and the weather experienced during the preceding months. Therefore, their partial regression models can be summarized as follows:
log(ni) = log(n) + log h(X1) + log h(X2)+.... + log h(Xi), (4)
Where, log(n) and h(.) denote the logarithm of the geometric mean of population size in spring and functions that determine the effects in response to environmental conditions Xi=[xi,1,..., xi,q] (q denotes the number of environmental factors).
Results of simple deterministic simulations
Figure 1 shows the results of the simple deterministic simulations. In these Figure 1 simulations, although the values on the y-axis were different, all the patterns were the same. That is, I did not assume a density-dependent effect in the relationship between log(dni) and log(n1,i) in these simulations, even though the resultant plots showed decreasing trends in all cases. All the slopes of the regression lines of log(dni) against log(n1,i) were -1, and all were statistically significant. In Figures 1a, b, and c, the significant provability (p values) was 0.033 (<0.05); in Figure 1d, it was 0.022, which was smaller than those in Figures 1a, b, and c, because the number of data sets increased from 9 to 18.
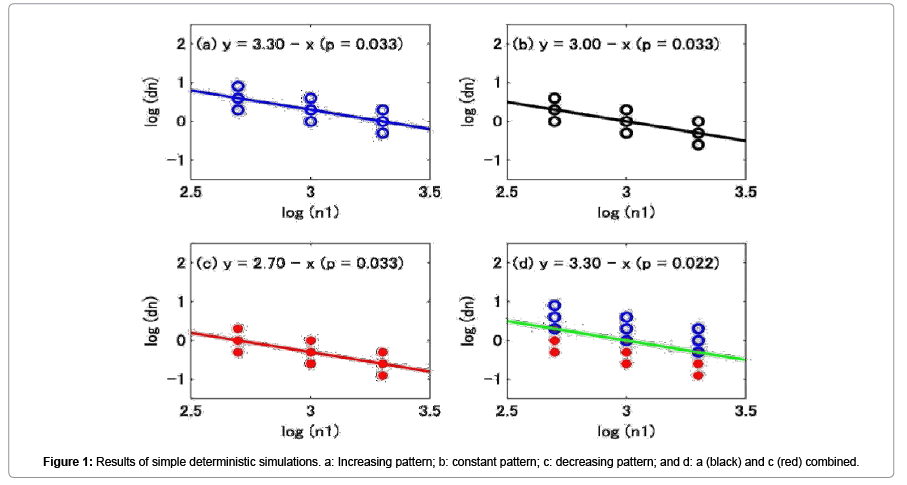
Figure 1: Results of simple deterministic simulations. a: Increasing pattern; b: constant pattern; c: decreasing pattern; and d: a (black) and c (red) combined.
Results of Monte Carlo simulations
Table 2 shows the logarithm transformed thrip data from Table 3 in Davidson and Andrewartha [1]. The averages and standard deviations, respectively, were 1.19 and 0.59 in October and 2.08 and 0.44 in November. By using these averages and standard deviations, Monte Carlo simulations were conducted. Figure 2 shows the histogram of the slopes for 1000 trails (Figure 2a) and that of the slopes that were significantly negative by a 5% significance level in the 1000 trails (Figure 2b). In this case, 687 out of 1000 trials showed significantly negative slopes with a 5% significance level. That is, 68.7% of the trials erroneously indicated the existence of density-dependent effects in the relationship between population change and population size.
| Number of samples = 7 | s.d. (Nov.) | |||
|---|---|---|---|---|
| 0.75 | 1.00 | 1.25 | ||
| s.d. (Oct.) | 0.75 | 687 | 493 | 346 |
| 1.00 | 849 | 687 | 529 | |
| 1.25 | 932 | 817 | 687 | |
Table 3: Results of sensitivity tests when the standard deviation was changed.s.d denotes the standard deviation. Figures show the number of trials in which the slope was significantly negative.
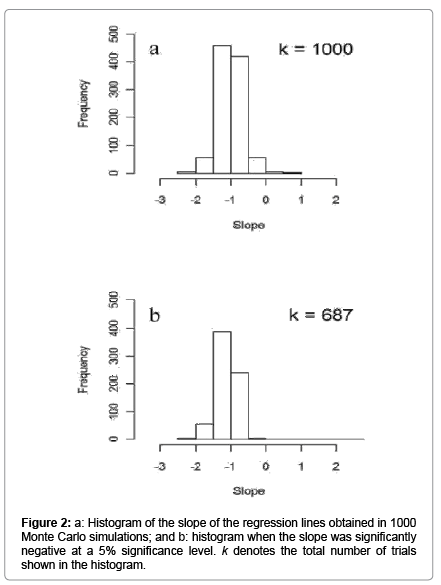
Figure 2: a: Histogram of the slope of the regression lines obtained in 1000 Monte Carlo simulations; and b: histogram when the slope was significantly negative at a 5% significance level. k denotes the total number of trials shown in the histogram.
Results of sensitivity tests
Table 3 shows the results of the sensitivity tests for the number of trials in which the slope was significantly negative when the standard deviation in the population size in October and/or November was changed. When the standard deviation in October (independent variable) increased by 1.25 times than that observed, the percentage of misjudges increased. On the other hand, when the standard deviation in November (dependent variable) decreased by 0.75 times than that observed, the percentage of misjudges increased. When the levels of decreasing and increasing standard deviations were the same in both October and November (e.g., 0.75 times in October and 0.75 times in November or 1.25 times in October and 1.25 times in November), the percentage of misjudges was the same at 68.7%. The standard deviation in October was large and that in November was small, the percentage of misjudges was the highest, and vice versa.
Table 4 shows the results of sensitivity tests for the number of trials in which the slope was significantly negative when the number of samples changed from 7 to 14. This effect was extremely large. In nearly all cases, the slopes of the regression lines indicated the false decreasing trends.
| Number of samples = 14 | s.d. (Nov.) | |||
|---|---|---|---|---|
| 0.75 | 1.00 | 1.25 | ||
| s.d. (Oct.) | 0.75 | 964 | 883 | 737 |
| 1.00 | 999 | 964 | 914 | |
| 1.25 | 1000 | 995 | 964 | |
Table 4: Results of sensitivity tests when the standard deviation was changed. Figures show the number of trials in which the slope was significantly negative.
Re-analysis results of the data from Davidson and Andrewartha [1]
Figure 3 shows the reanalysis results of the data from Davidson and Andrewartha [1]. Figure 3 (Top left) shows the trajectories of the log transformed thrip population size in October and November. The patterns of the trajectories were similar to each other. Figure 3 (Top right) shows the relationship between population change log(dn) and population size in October. As mentioned previously, the population change log(dn) was defined using log(nNov) - log(nOct). The regression line indicated the negative slope at -0.342 and showed a 10% statistical significance. Figure 3 (Bottom left) shows the regression line of the population size in November against that in October. The slope of the regression line estimated using a simple regression method was 0.656 (p=0.008), and the 95% confidence interval of the slope was [0.260, 1.051]. The slope of the regression line estimated using the Deming regression method was 0.713, and the 95% confidence interval of the slope was [0.529, 1.718].
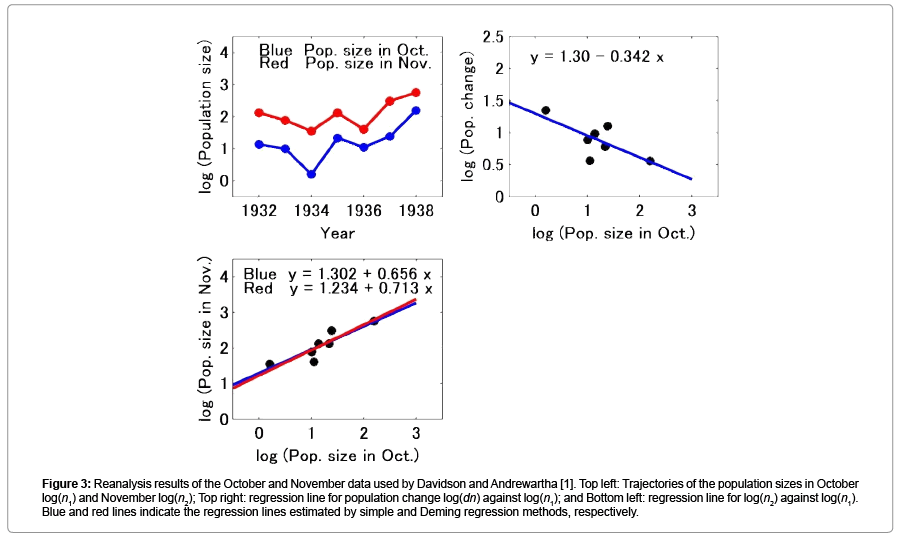
Figure 3: Reanalysis results of the October and November data used by Davidson and Andrewartha [1]. Top left: Trajectories of the population sizes in October log(n1) and November log(n2); Top right: regression line for population change log(dn) against log(n1); and Bottom left: regression line for log(n2) against log(n1). Blue and red lines indicate the regression lines estimated by simple and Deming regression methods, respectively.
Figure 4 shows the trajectories of the log transformed thrip population size in month i-1 and month i (i=May, Jun, …, Mar., except Oct.), respectively. Figure 5 shows the relationship between population size in month i against that in month i-1, (i=May, Jun, …, Mar., except Oct.), and shows the regression lines estimated by simple method (blue line) and Deming regression (red line), respectively. The slopes estimated are also shown in Table 5. In the simple regression, in only 3 cases (Oct.-Nov., Dec.- Jan., and Jan.-Feb.), the slopes were not significantly different from unity. For other 2 cases (Apr.-May, and Aug.-Sep.), the 95% confidence interval of the slopes were extremely large however they were not significantly less than unity. Figure 6 shows the results when all data from April 1932 to December 1938 were combined. Then the number of samples was 74. In this case the slope of the regression line estimated using the simple regression method was 0.597 (p=1.897 × 10-8), and the 95% confidence interval of the slope was [0.409, 0.786]; The slope estimated by the Deming regression was 1.002 and the 95% confidence interval was [0.814, 1.338], then the slope was not significantly different from unity.
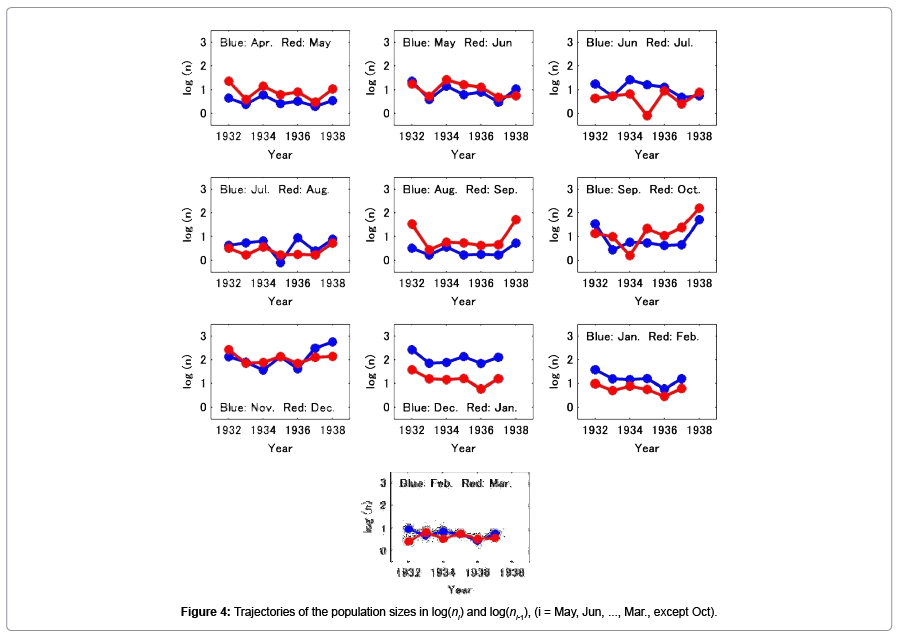
Figure 4: Trajectories of the population sizes in log(ni) and log(ni-1), (i = May, Jun, ..., Mar., except Oct).
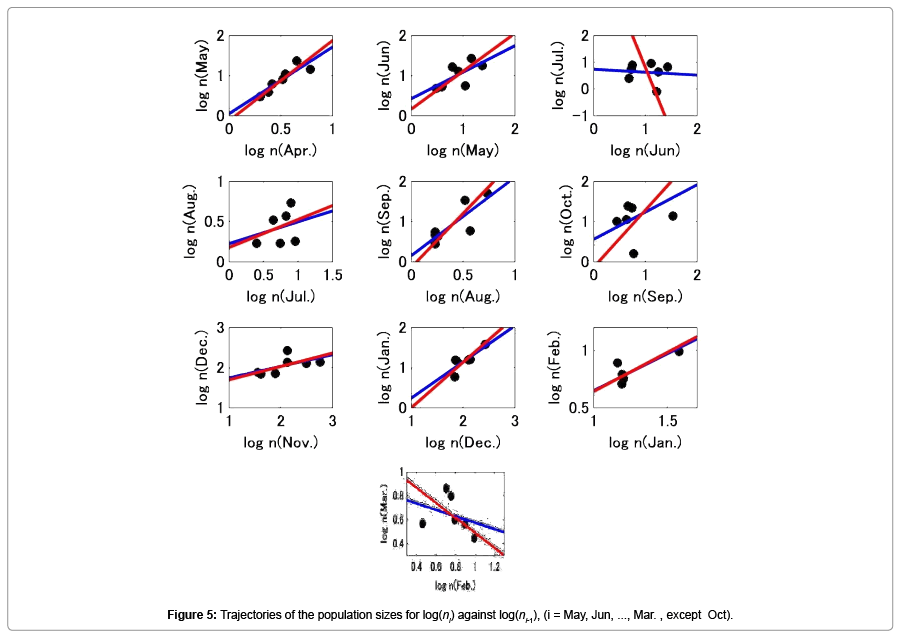
Figure 5: Trajectories of the population sizes for log(ni) and log(ni-1), (i = May, Jun, ..., Mar., except Oct).
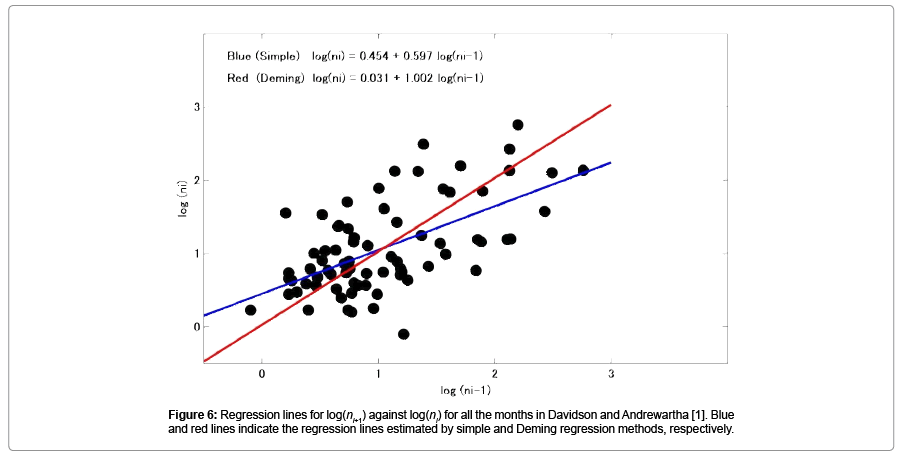
Figure 6: Regression lines for log(ni-1) against log(ni) for all the months in Davidson and Andrewartha [1]. Blue and red lines indicate the regression lines estimated by simple and Deming regression methods, respectively.
| Simple regression | ||||
|---|---|---|---|---|
| ni-1 | ni | Slope | 95% confidence intarval | p-value |
| Apr. | May | 1.661 | (0.680, 2.64) | 0.00733 |
| May | Jun | 0.661 | ( -0.142, 1.463,) | 0.0880 |
| Jun | Jul. | -0.109 | ( -1.510, 1.292) | 0.850 |
| Jul. | Aug. | 0.271 | (-0.294, 0.836) | 0.272 |
| Aug. | Sep. | 1.950 | (0.449, 3.451) | 0.0206 |
| Sep. | Oct. | 0.679 | (-0.487, 1.845) | 0.195 |
| Oct. | Nov. | 0.656 | (0.260, 1.051) | 0.00802 |
| Nov. | Dec. | 0.288 | (-0.166, 0.741) | 0.164 |
| Dec. | Jan. | 0.901 | (0.00776, 1.793) | 0.0488 |
| Jan. | Feb. | 0.645 | (0.253, 1.036) | 0.0103 |
| Feb. | Mar. | -0.271 | (-1.409, 0.868) | 0.546 |
| All data | 0.597 | (0.409, 0.786) | 1.897 (10-8) | |
| Deming regression | ||||
| ni-1 | ni | Slope | 95% confidence intarval | |
| Apr. | May | 1.998 | ( 1.715, 2.735) | |
| May | Jun | 0.944 | (-2.114, 1.686) | |
| Jun | Jul. | -4.775 | -5.581, 0.603 | |
| Jul. | Aug. | 0.348 | ( - , - ) | |
| Aug. | Sep. | 2.684 | (2.137, - ) | |
| Sep. | Oct. | 1.424 | (0.664, 11.978) | |
| Oct. | Nov. | 0.713 | (0.528, 1.718) | |
| Nov. | Dec. | 0.335 | (0.133, 0.841) | |
| Dec. | Jan. | 1.133 | (0.279, 1.356) | |
| Jan. | Feb. | 0.682 | (-2.450, 0.738) | |
| Feb. | Mar. | -0.635 | (-2.004, 0.600) | |
| All data | 1.002 | (0.814, 1.338) | ||
Table 5: Intervals and p values estimated using a simple or the Deming regression.
The results of the simulations showed that even when a densitydependent effect did not exist, an apparent density-dependent effect was erroneously detected from the regression line plotted for population change against population size. Therefore, when such negative slopes of the regression lines are obtained, we should not judge that a densitydependent effect truly exists.
In this analysis, I defined population change by log(n2) - log(n1) instead of log[(n2-n1)/n1], because I wanted to analyze the change when n2 is smaller than n1. The population size in November was greater than that in October; therefore, when the population size in October is used as n1 and that in November is used as n2, the definition log[(n2 - n1)/n1] could be used for the population change. However, the results obtained were nearly same as that by using the former definition. The slope of the regression line was -0.393, and the significant probability was 0.0808. The analyses for each month, because the sample sizes were limited, then only in 5 cases, the slope of the regression lines between population size in month i against that in month i-1 were significant with 5% significant level. In there, only for 3 cases (Oct.-Nov., Dec.- Jan., and Jan.-Feb.), the slopes were not significantly different from unity. For other 2 cases (Apr.-May and Aug.-Sep.), the 95% confidence interval of the slopes were extremely large, however, the interval did not show less than unity. This means that there was no case that showed the negative density-dependent effect between the population sizes in the successive two months. When the regression line of log(ni) against log(ni-1) for all data was estimated using the simple regression method, the slope was less than unity. However, as Sakuramoto and Suzuki [11] showed, the regression line less than unity was considered to be produced by process and/or observed errors. When the Deming regression was applied to the data, the slope was not statistically different from unity. Then, the relationship can be expressed using log(nNov)=log(a) + log(nOct) or nNov = a•nOct. This indicates that the population size in month i could be determined in proportion to the population size in month i-1, i.e., the relationship between the populations sizes in month i and that in month i-1 can be expressed using a simple proportional model. The validity of a proportional model is not explicitly noted, however, the fluctuations of the catches in the species such as snow crab [22], sandfish [23,24] and walleye pollock [25] are already explained without assuming a density-dependent effect in the fluctuation mechanism in catches. The population fluctuation in the Peruvian anchoveta [26] is explained well using the same concept of the fluctuation mechanism proposed in this paper. That is, my concept for the mechanism in population fluctuation coincides well with that by Davidson and Andrewartha [16] and Andrewartha and Birch [27] in that the densitydependent effect is not a key factor or a negligible factor in controlling the population size.
However, an important difference in the concept between Andrewartha and Birch [27] and that proposed by me is that they believed that the essentially important factors in controlling population size were the succession of good and bad seasons and the heterogeneity of places where they live [17], and they did not refer to the proportional relationships of population size between each month. On the contrary, my proposed concept is that the population size in month i, is proportional to that in month i-1 and environmental conditions in month i multiplicatively add to the population size in month i. This simple mechanism can also explain the concept of the fluctuation mechanism proposed by Davidson and Andrewartha [16] and Andrewartha and Birch [27] as shown in equations (2) and (4). This seems to be the same mechanism that Sakuramoto [10,28] proposed for the stock-recruitment relationship in Japanese sardine. That is, recruitment Rt is essentially determined in proportion to spawning stock biomass St, and then environmental factors in year t further change the recruitments. The subscripts showing years are all t, however, there are time lags. St, and Rt denote the biomass in January and the number of recruitment over March to May, respectively. The environmental factors are sea surface temperature over the southern area of the Kuroshio Extension in February and Arctic oscillation in February. Therefore, the fluctuations mechanism for Japanese sardine, which lives in the sea, and that for T. imaginis, which lives on the land, could be explained by the same concept of fluctuation mechanism; population size in time i is essentially determined in proportion to that in time i-1, which is further changed in response to environmental conditions in time i [29].
I thank to the great efforts expended by Davidson, Andrewartha and their groups in collecting the detailed data. I also respect their contributed works. I thank to Drs A. Ohno, N. Suzuki, H. Sugiyama and S. Hasegawa for their useful comments. I thank two anonymous reviewers for their many useful suggestions that led to improvements in the manuscript. Editage, a division of Cactus Communications, for their assistance with translating and editing this manuscript.