Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2015) Volume 6, Issue 1
The efficacy and effect of tamoxifen and letrozole on sex reversal in common carp and Nile tilapia was studied using free swimming fry of Cyprinus carpio and tilapia Oreochromis niloticus. Treatment of letrozole in the dose of 100 mg kg-1 feed brought about 79.39 ± 1.09% masculinisation in C. carpio and 87.91 ± 1.39% masculinisation in O. niloticus, while larger dose (200 mg kg-1 feed) produced 98.47 ± 1.34% male C. carpio and 99.65 ± 0.72% male O. niloticus. In the control group, there was 48.28% male and 50.78% female C. carpio while 46.38% male and 53.616% female O. niloticus. Letrozole significantly increased serum testosterone (T) level suppressing 17β-estradiol (E2) production and the androgenizing action was more potent when compared with tamoxifen treatment. The histological examination of letrozole treated C. carpio and O. niloticus revealed increased number of irregularly spread spermatids and there was hardly any difference in control testes and letrozole treated gonads. The results showed potent and complete action of letrozole (Cyp19) on sex reversal for monosex male fish production delineating its commercial application in aquaculture.
Keywords: Masculinisation; Sex steroids; Tamoxifen; Letrozole; Nile tilapia; Common carp
Early maturation and frequent spawning are management challenges when working with tilapia and common carp. Sex control for aquaculture and research purpose has been attempted in many fish species by administration of exogenous hormones such as several androgens and estrogens for production of all-male or all-female populations [1-3]. At the same time, use of aromatase inhibitors (AIs) for sex reversal have also been attempted in mammals [4], birds (chickens) [5,6], amphibians [7], reptiles [8] and fishes [9]. However, the efficacy of aromatase inhibitors (AIs) is yet to be evaluated in important aquaculture species such as tilapia and common carp for effective sex reversal and monosex production. It is important to mention that male tilapia is preferred for culture because of faster growth while reverse is true for common carp [10]. The common carp C. carpio L. has a XX/XY type chromosomal genetic sex determination [1,11] and inverted XX males, produced by endocrine sex reversal technique, has been used for mating with normal females (XX) so as to achieve all-female population [1,11] which have a faster growth rate than mixed populations, and prevent uncontrolled reproduction.
It has been hypothesized that male and female sex differentiation is determined by aromatase activity in fish as Cyp19 gene is highly conserved in fish [12]. Estrogen biosynthesis is mediated by the steroidogenic enzyme Cyp19, which converts androgens to estrogens [12,13]. The key role of aromatase in ovarian differentiation is supported by the fact that the inhibition of estrogen synthesis in the differentiating ovaries can trigger partial or complete masculinisation in fish [12]. Tamoxifen and letrozole are known to influence the production of estradiol in mammals [14,15] as well as in fishes [9,16-18]. The mode of action of tamoxifen has been described to exert its main anti-estrogenic effect by competing with estradiol for its own receptor [9,14-15]. A likely mechanism of tamoxifen action has also been reported through highly reactive Cyp19-mediated tamoxifen metabolites [14,15]. However, letrozole has been reported as a highly potent inhibitor of aromatase (Cyp19) in vitro and in vivo in fishes [12], and in humans [14]. Tamoxifen is a known selective estrogen receptor modulator (SERM) and works through blockage of estrogen receptors [16] while letrozole is supposed to effectively down regulate Cyp19a1a or Cyp19a1b genes acting directly on aromatase production [12]. In this study, we have investigated the comparative performance of tamoxifen and letrozole on sex determination in a gonochorist cichlid O. niloticus and a cyprinid Cyprinus carpio hypothesizing that in these fish the sex determination is Cyp19 gene dependent and they will respond similarly for the two drugs. The present investigation was designed to better understand the aromatase based sex determination in fish using two functional treatments of tamoxifen (11β-hydroxyandrostenedione) an estrogen receptor blocker and letrozole (Cyp19) which is a known aromatase inhibitor.
Six hundred free swimming fry of 30 days post fertilized (30 dpf) C. carpio and O. niloticus (15 dpf) (length, 0.68 ± 0.21 cm and weight, 0.37 ± 0.08 g) were procured and equally divided into six glass aquaria (150 L). The reason of procuring different age groups of C. carpio and O. niloticus was simply because both were sexually undifferentiated and were able to actively accept external feeding. The C. carpio as well as O. niloticus were acclimatized to laboratory condition for one week maintaining in separate aquaria. Water temperature in the aquaria was maintained with the help of thermo-controlled heaters (300 W Risheng, Made in China) with filters at 25 ± 1°C. Tamoxifen (SIGMA Chem. Co. USA) and letrozole (USP, Evalet™, India) were dissolved in minimal volume of 95% absolute alcohol and the required volume of these chemicals was sprayed over a commercial pellet feed (Tiayo Pvt. Ltd.) to make them in the doses of 100 and 200 mg kg-1 feed and then dried in an oven. The doses of tamoxifen and letrozole were decided in consultation with available literature. This tamoxifen and letrozole mixed feed [Tamoxifen{100 mg kg-1 (T1) and 200 mg kg-1 (T2)} as well as Letrozole {100 mg kg-1 (L1) and 200 mg kg-1 (L2)] were given to the fingerlings of C. carpio and O. niloticus daily twice a day up to satiation for sixty days. The commercial pellet diet was consisting of crude protein (32%), crude fat (4%), crude fiber (5%) and moisture (10%). The control group of fingerlings was given tamoxifen or letrozole free similar diet. Since there was no circulatory water system, water of all experimental aquaria was completely changed every third day to eliminate the metabolites, possibility of accumulation of any steroid and to maintain the water quality.
At the end of 60 days, fishes were anesthetized with tricaine methanesulfonate (MS-222) (Merck, Germany) dissolved in water (100 mg L-1), buffered with sodium bicarbonate (100 mg L-1). Blood of anesthetized fish was drawn from caudal vein and pooled for 5 live specimens of different treatment groups as well as control separately since blood drawn from individual fish was insufficient for hormonal examination. About 2 ml blood was taken and centrifuged at 3000 rpm using an Eppendorff Centrifuge for 5 min to obtain serum which was stored at -80°C until used for hormonal assay. Serum 17β- estradiol and testosterone were estimated using enzyme linked immune absorbent assay (ELISA) technique by using kit provided by Enzo-Life Sciences, India. The optical density (OD) was read on ELISA reader and the observation were plotted using logit- log paper. The limit of detection for testosterone assay was 5.67 pg mL-1 while it was 14.0 pg mL-1 for E2 .
For all experimental groups, sex differentiation was studied using histological technique was used while sex frequency and sex change was assessed using acetocarmine squash method [19]. For histological examination, fishes were cut ventrally using a scalpel, from the genital papilla to the base of the pectoral fin. A window on the lateral side was opened and the viscera were removed, leaving gonads, swim bladder and kidneys in place. The pair of gonad was identified based on the morphology and location of the gonadal tissue. After macroscopic examination, the anterior and posterior ligaments were cut, and both gonads were removed using a forceps and fixed in 10% natural buffer formalin solution for histological examinations. Fixed tissue was dehydrated in different grades (30%, 70%, 90% and absolute) of ethanol and embedded in paraffin wax. Sections (5-7 μm) of gonadal tissues were cut, de-waxed and dehydrated with different grades of ethanol, stained with Haematoxylin and Eosin, dehydrated and mounted in Canada balsam (Merck, Darmstadt, Germany). Sections were examined under a compound microscope (Olympus Co., Japan).
All data were calculated and presented as mean ± standard deviation (X ± SD). Student’s t- test was calculated for observing the comparison between control and treatment groups. The coefficient of variance was also calculated so as to know the efficacy of the treatment groups. Further, ‘Chi square’ (χ2) test was done for calculating the variation in sex ratio in the treatment groups from the expected Mendelian sex ratio of 1:1. Data were analyzed by one way analysis of variance (ANOVA) using a computer programmed software, statistical package for Social science (SPSS).
Experimental fishes of control, tamoxifen and letrozole treated groups remained healthy and mortality ranged from 8.5% to 14.5% throughout the observation period of 60 days. Higher mortality was mainly in the tamoxifen treated fish particularly at the end of 60 days. Sex ratio of the experimental fishes was found to respond well with tamoxifen and letrozole treatments. There was significantly enhanced masculinisation both in C. carpio and O. niloticus. Low dose tamoxifen treated (T1) C. carpio showed 63.21% male and 36.79% female while in O. niloticus, there were 78.36% male and 35.80% female (Figure 1). Larger dose of tamoxifen (T2) brought about 84.56% male and 15.44% female in C. carpio and 90.01% male and 9.73% female populations of O. niloticus (Figures 1 and 2). Low dose of letrozole treated group showed 79.39% male and 20.61% female C. carpio; 87.91% male and 11.86 % female O. niloticus. Larger dose of letrozole (L2) brought about 98.47% male and 1.53% female C. carpio and 99.65% male and 0.52% female O. niloticus. Percentage of masculinisation was comparatively more in letrozole treated fish as compared to tamoxifen treatment. In the control group, there was 48.28% male and 50.78% female C. carpio and 46.38% male and 53.616% female O. niloticus (Figures 1 and 2). Some intersex fish was also observed in low dose of tamoxifen (T1) and letrozole treatments (L1) to fish. In C. carpio, tamoxifen treatment (T1) showed 0.30 % and letrozole treatment (L1) showed 0.40 % intersex (Figures 1 and 2). In O. niloticus, low dose of tamoxifen (T1) showed 1.0% intersex and low dose of letrozole (L1) showed 0.85% intersex. The statistical coefficient of variance (CV) for male was 20.56 and 8.76 for female after tamoxifen treatment to tilapia and it was 7.33 for male and 18.73 for female in letrozole treated O. niloticus. The calculated χ2 value for sex ratio was 17.64 which was close to degree of freedom (df) 15.51 showing that the deviation in sex ratio from the expected Mendelian sex ratio was highly significant (P<0.001). Similarly, the statistical CV for male was 7.97 and 3.83 for female common carp after tamoxifen treatment. In letrozole treated C. carpio, the CV was 4.07 for male and 18.27 for female. The calculated χ2 value was 33.65 which was much less than the chi-square Table value and was 26.13. Hence, there was highly significant (P<0.001) variation in sex ratio from the expected Mendelian sex ratio of 1:1.
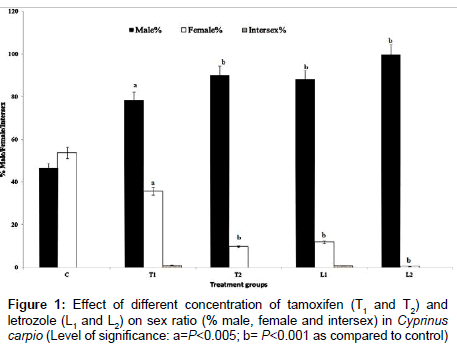
Figure 1: Effect of different concentration of tamoxifen (T1 and T2) and letrozole (L1 and L2) on sex ratio (% male, female and intersex) in Cyprinus carpio (Level of significance: a=P<0.005; b= P<0.001 as compared to control)
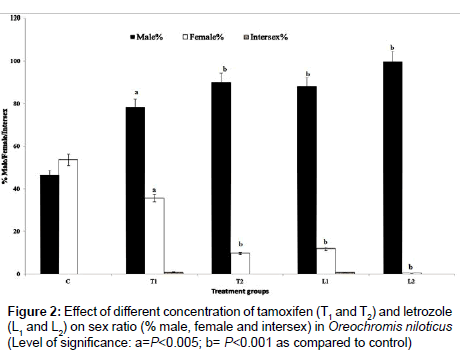
Figure 2: Effect of different concentration of tamoxifen (T1 and T2) and letrozole (L1 and L2) on sex ratio (% male, female and intersex) in Oreochromis niloticus (Level of significance: a=P<0.005; b= P<0.001 as compared to control)
In control Nile tilapia and common carp, the level of 17β-estradiol (E2) was 162.66 pg mL-1 and 196.5 pg mL-1 and testosterone (T) level was 610.23 pg mL-1 and 578.56 pg mL-1 respectively. The 17β-estradiol and testosterone (E2/T) ratio was 0.266 for tilapia and 0.339 for common carp. However, E2/T declined significantly after tamoxifen and letrozole treatments (Figures 3 and 4). Larger dose of letrozole treatment (L2) to fish brought about more pronounced decline in E2 level as compared to tamoxifen treated fish consequently declining the E2:T ratio remarkably which was 0.076 for tilapia and 0.085 for common carp (Figures 3 and 4). However, it was observed that some E2 level was present even after suppressing the aomatase activity by both tamoxifen and letrzole. The lowest value of E2 was 85.83 pg mL-1 in tilapia while it was 56.0 pg mL-1 for common carp. The coefficient of variation (CV) for E2 in tamoxifen treated O. niloticus was 16.0 while for testosterone, it was 10.44. The CV for E2 in letrozole treated O. niloticus was 8.67 and it was 11.18 for testosterone (T) level. In case of common carp, the coefficient of variance for E2 was 8.81 in tamoxifen treated C. carpio and 6.64 for letrozole treatment. The CV for T for tamoxifen treated C. carpio was 15.9 and 11.34 for letrozole treatment.
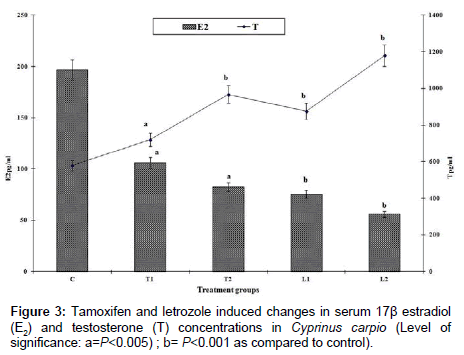
Figure 3: Tamoxifen and letrozole induced changes in serum 17β estradiol (E2) and testosterone (T) concentrations in Cyprinus carpio (Level of significance: a=P<0.005) ; b= P<0.001 as compared to control).
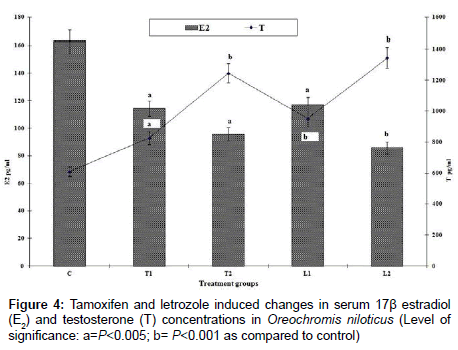
Figure 4: Tamoxifen and letrozole induced changes in serum 17β estradiol (E2) and testosterone (T) concentrations in Oreochromis niloticus (Level of significance: a=P<0.005; b= P<0.001 as compared to control)
Histological examination revealed that there was no difference in gonadal tissues between control males and letrozole treated C. carpio except localized presence of spermatozoa (Figure 5a). The paired testis of O. niloticus was elongated milky white organ covered by a thin connective tissue capsule (tunica albuginea). The histological sections, testis of O. niloticus showed organized branching lobules of the unrestricted spermatogonial type (Figure 5b). The ovarian cavity, oocytes and perinuclear oocytes were clearly seen in the ovary of female C. carpio and O. niloticus. Transitional phase of intersex gonad had arrested oocytes and spermatogenic germ cells at the late stages of spermatogenesis. In masculinising fish after tamoxifen treatment to C. carpio, testis contained spermatozoa accumulated in the seminiferous tubules. Histological examination of gonads revealed that few specimens exhibited intersex (ovotestis) i.e. presence of ovarian as well as testicular structures together in fishes treated with 100 mg kg-1 of letrozole and tamoxifen (Figure 5c). In the intersex fish induced by tamoxifen treatment, some spermatids were similar to those observed in the control males along with numerous arrested perinuclear oocytes in the same gonad. The intersex fish was partly masculinized and was having male as well as female cells in the same gonad (Figure 5c).
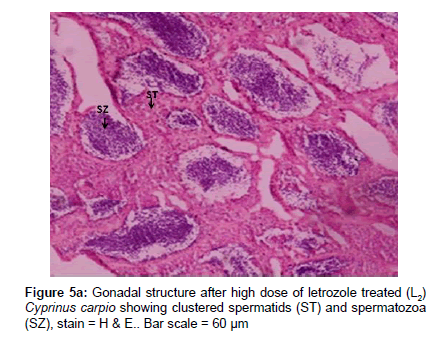
Figure 5a: Gonadal structure after high dose of letrozole treated (L2) Cyprinus carpio showing clustered spermatids (ST) and spermatozoa (SZ), stain = H & E.. Bar scale = 60 μm
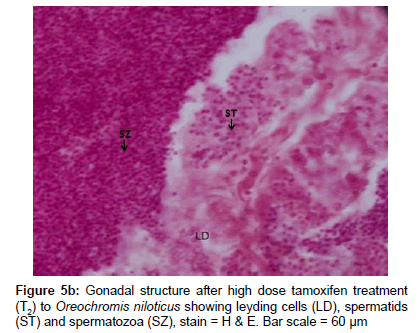
Figure 5b: Gonadal structure after high dose tamoxifen treatment (T2) to Oreochromis niloticus showing leyding cells (LD), spermatids (ST) and spermatozoa (SZ), stain = H & E. Bar scale = 60 μm
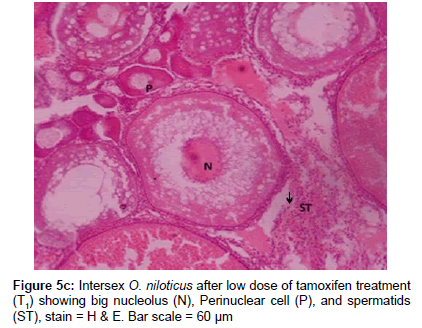
Figure 5c: Intersex O. niloticus after low dose of tamoxifen treatment (T1) showing big nucleolus (N), Perinuclear cell (P), and spermatids (ST), stain = H & E. Bar scale = 60 μm
The results of this study revealed that tamoxifen and letrozole in the tested doses had potent action on sex reversal and gonadal differentiation bringing about significantly high production of male C. carpio as well as O. niloticus. However, level of masulinization was found higher in the larger dose of letrozole which was more pronounced in case of O. niloticus as compared to common carp where almost all male tilapia was obtained. In our earlier study, we obtained 90% male tilapia when treated with tamoxifen in the dose of 200 mg kg-1 without any significant mortality [9]. Reports on other fish species have reported 100% male production by the use of AI treatment in the dose of 100 and 1000 mg kg-1 in the zebra fish Danio rerio [20]; and 500 mg kg-1 in the golden rabbit fish Siganus guttatus [21]. Our findings corroborates with earlier the report [22] where fadrozole in the doses of 200 mg kg-1 to 400 mg kg-1 produced approximately 97% male population of C. carpio. However, it has been reported [23,24] that tamoxifen or 17α-methyltestosterone (MT) masculinized 45- 50% C. carpio at 100 mg kg-1 experimental feed which has much lower level of sex reversal % than the obtained value in this study. Effect of tamoxifen on sex differentiation has been reported in Zebra fish, Danio rerio [25], sea scallop, Placopecten magellanicus [26], and tilapia [9]. It has also been reported that oral administration of tamoxifen (200 ppm) effectively acts for sex reversal in the bagrid catfish during the labile period of sex differentiation [27] and in Tinca tinca [28]. Tamoxifen, as an anti-estrogen analogue has also been evaluated as a feed additive to regulate the sex ratio in rainbow trout Oncorhynchus mykkis [29], O. niloticus (Hines and Watts 1995), and Japanese medaka Paralichthys olivaceus [30].
The analysis of estradiol (E2) levels in the blood serum showed declined values after letrozole and tamoxifen treatments in comparison with the controls, whereas significantly high serum testosterone (T) level was detected in all treatment groups. However, there was some estradiol level left in the circulation even after aromatase inhibition which could be due to two reasons; firstly the blood sample was pooled where specimens of both the genetic sexes might have been present and alternately some estradiol level may be needed for maintenance of testicular activity since Cyp19 gene is highly conserved in fish [12]. Our findings support the earlier findings of [31] where significant differences in concentrations of E2 in the blood plasma and hypothalami between fadrozole (5, 50 or 500 μg kg-1) treatment groups of goldfish and C. carpio has been reported. Fadrozole in the doses of 0, 100, 500 and 1000 μg/fish has been reported to decrease the plasma E2 in honeycomb grouper Epinephelus merra and Nile tilapia [32-34] which further supports our findings. Decreased serum E2 level has also been reported after the treatment of fadrozole (10.0 mg kg-1) to red-spotted grouper E. akaara [35].
Complete masculinisation in O. niloticus and C. carpio significantly accounted for suppressed E2 level which consequently elevated T level suggesting that aromatase based sex determination occurs in Nile tilapia and common carp. Our findings strongly supported that letrozole directly down regulate Cyp19 gene thereby inhibiting the aromatase production [12]. However, the masculinisation level with tamoxifen was bit different because anti-estrogenic action [36] and selective estrogen receptor modulating (SERM) action through blockage of estrogen receptors [9,16]. Since the masculinisation level was potent with letrozole treatment, our findings suggested that masculinising with commercially available letrozole (USP, Evalet™, India) could be a cheap method of inducing sex reversal in tilapia and common carp. This drug is non-steroidal aromatase inhibitor; inhibitor of estrogen biosynthesis and anti-tumour agent available in market for breast cancer treatment.
Histological examinations of the tamoxifen and letrozole treated gonads showed well differentiated testicular cells showing spermatozoa in seminiferous tubules suggesting that the sex reversal was male hormone (testosterone) based since letrozole blocked the aromatization process [12,37] while tamoxifen inhibited receptor binding. In this study, there was a notable, dose-dependent enlargement of the seminiferous tubules accompanied by an abundant accumulation of spermatozoa in the lumina after letrozole treatment. The marked accumulation of spermatozoa in testes after latrozole treatment could be due to enhanced sperm production related to the increase in plasma testosterone [38]. However, presence of intersex gonads could be a result of low dose of tamoxifen or letrozole treatment. Further, results of this study suggested that 60 days exposure of AIs in the dose of 200 mg kg-1 produced complete sex reversal in tilapia and common carp.
We conclude that letrozole at a dose 200 mg kg-1 significantly inhibit estrogen production thereby increasing level of circulating testosterone via suppressed action of Cyp19 binding which significantly influenced sex differentiation for masculinisation in C. carpio and O. niloticus. The masculinised fish had well-developed testes containing accumulated spermatozoa in the seminiferous tubules. The putative action of tamoxifen and letrozole was found more or less similar for their masculinizing effects; however, the effect of letrozole was more potent suggesting that the chemical could be used in reproductive management and sex reversal of such important aquaculture species. It is to mention here that the letrozole exposed fishes retained the sex reversed masculinity until six months of observation even after withdrawal of the treatment suggesting that gonochoristic fish particularly common carp and tilapia maintain their sexual plasticity until adulthood [39]. There is hardly any report to document that exogenous AI treated fishes are not safe for consumption [40,41]. Rather most potent estrogen blockers and aromatase inhibitors are reported to occur naturally as Chrysin (5’7-Dihydroxyisoflavone) a flavonoid from blue passion flower and White Button Mushrooms with great real life results having therapeutic values [42]. It is therefore, the results of this study to produce monosex tilapia and common carp using AI has strong potential having avenues in aquaculture industry.
Authors are grateful to Dr J.K.Jena, Director NBFGR for extending all support and encouragements. We thankfully acknowledge the help received by the Institute of Toxicological Research (IITR), Lucknow for reading out our samples over ELISA reader. We also thankfully acknowledge JVD Pharma Pvt.Ltd., New Delhi for providing us letrozole tablets (EvaletTM) for our experimental use.