Pancreatic Disorders & Therapy
Open Access
ISSN: 2165-7092
ISSN: 2165-7092
Research Article - (2013) Volume 0, Issue 0
Pancreatic cancer (adenocarcinoma) remains a deadly untreatable cancer with no effective early detection procedure. Little is known concerning the factors involved in the development of pancreatic malignancy, which impedes advancements in its treatment and detection. Altered cellular zinc has been implicated in several cancers. Recent studies provide evidence that zinc and zinc transporters are important factors in pancreatic cancer. This review discusses the current information relating to the status of zinc and zinc transporters in human pancreatic adenocarcinoma. Relationships of the physiology and biochemistry of zinc in mammalian cells are presented, which should be applied to the conduct, interpretation, and translational application of human studies and experimental models. Evidence from human pancreatic tissue studies supports a new concept of the role of zinc in the development of pancreatic adenocarcinoma. The zinc level of the normal ductal and acinar epithelium is markedly decreased in the development of the malignant cells and the premalignant PanIN cells. ZIP3 is identified as the likely zinc uptake transporter, which is down regulated concurrently with the loss of zinc. Ras responsive binding protein (RREB1) is identified as the possible transcription factor involved in the silencing of ZIP3 expression. The evidence supports the current views of transdifferentiation of PanIN epithelium to ductal adenocarcinoma, and the possibility that acinar epithelial dedifferentiation might be a source of premalignant cells. These zinc-associated events occur early in oncogenesis to protect the malignant cells from the cytotoxic effects of zinc levels that exist in the normal cells. Hopefully, this presentation will stimulate interest in and support for much needed research into the implications of zinc and zinc transporters as important events in pancreatic carcinogenesis. The potential exists for the RREB1-ZIP3- zinc concept and/or other implications of zinc as new approaches for the development of effective treatment and for diagnostic biomarkers for pancreatic cancer.
<Keywords: Pancreatic cancer, Ductal adenocarcinoma, Zinc, ZIP3, Zinc transporter, RREB1, PanIN
Pancreatic cancer (adenocarcinoma) is expected to result in ~44,000 new cases/year in the U.S., of which ~38,000 deaths will occur [1]. The 5-year survival rate has remained at ~6% or less for the past 35 years. These statistics demonstrate the absence of and need for effective treatment of pancreatic cancer, and for effective biomarkers for early detection. This lack of significant progress in dealing with pancreatic cancer is due to the poor understanding and resolution of the factors and events involved in the development of pancreatic malignancy and its progression to advanced stages.
Zinc and zinc transporters have been implicated in several cancers; although the specific relationships are largely undefined in most cancers [2-4]. The exception is prostate cancer [5], in which the role of zinc, specific zinc transporters, and the clinical implications represent the most studied and understood of all cancers; although many important issues remain unresolved.
In contrast, the possible involvement of zinc in human pancreatic cancer has received very little attention, and is just beginning to evolve. There now exists compelling initial evidence (described below) for the implication of zinc and zinc transporters in the development and progression of pancreatic cancer. However, divergent views currently exist as to the specific relationships of zinc in human pancreatic cancer. Some of these issues arise from the lack of understanding of the physiology and biochemistry associated with the status of zinc in mammalian systems; and the questionable application and translational interpretation of molecular genetics and proteomics relating to the identification and functional role of zinc transporters in cells.
This critical review is intended to provide an assessment of the current information relating to the status of the role of zinc and zinc transporters in relation to the development and progression of human pancreatic adenocarcinoma. The divergent views and issues regarding the role of zinc and specific zinc transporters that currently exist are discussed with reasons and explanations that contribute to such conflicting conclusions. A new concept is presented of the role of zinc and ZIP3 zinc uptake transporter in the development of pancreatic adenocarcinoma.
Hopefully, this presentation will be informative to the biomedical and clinical research community about the potential important implications of zinc relationships in pancreatic cancer; and will enhance investigations and support for needed research to establish the role of zinc in pancreatic cancer. For those who will enter this field of research, the presentation might be helpful in their employment of experimental and clinical design and conditions that are representative of the physiological and biochemical relationships of zinc in mammalian systems; so as to provide appropriate translational interpretations. Under such conditions, the zinc relationships will lead to new approaches for the development of reliable biomarkers for identification of early development and perhaps at-risk subjects for pancreatic cancer; and for the development of efficacious therapy and prevention of pancreatic cancer.
Some important zinc and zinc transporter relationships
Many contemporary reports of zinc relationships in cancer and other clinical conditions have lead to questionable and inappropriate conclusions and misleading translational interpretations regarding the role of zinc and the involvement of zinc transporters. For those interested in zinc relationships in pancreatic cancer, an understanding of the status of zinc in mammalian systems, and its transport and trafficking in cells is essential for the conduct of experiments and for the analysis, interpretation, and clinical application of the results. Since an extensive description of these zinc relationships is not possible, we refer the reader to our recent review of the trafficking, transport, and reactivity of zinc in mammalian cells [6], and other excellent reviews [7-10]. The following will highlight some important zinc relationships that are relevant to the specific issues that are the focus of this presentation.
The first important relationship is the cellular and extracellular (especially the plasma and interstitial fluid compartments) zinc composition. The common use of the designation of intracellular and extracellular zinc as “zinc ion”, or as “free Zn++ ion” is an inaccurate and misrepresentation of the physiological/biochemical status of zinc. As Vallee and Falchuk [11] described, “In biological systems, very little, if any, zinc is free in solution.” The total normal concentration of zinc in plasma is ~15 uM; of which free Zn++ ion concentration is ~0.1- 1.0 nM. The intracellular total zinc concentration is ~200-500 uM in most mammalian cells; of which the free Zn++ ion concentration is in the pM range. Therefore, essentially all of the zinc in these biological systems is bound to organic ligands. For purposes of reactivity and trafficking, the organic zinc ligands can be characterized as being a) ligands which tightly bind zinc as a pool of immobile, unexchangeable, unreactive Zn; and b) ligands which loosely or moderately bind zinc as a pool of mobile, exchangeable, reactive zinc. The former generally constitutes ~95% of the total zinc; and the latter constitutes ~5%, including the negligible free Zn++ ions. Consequently, cellular trafficking, zinc reactivity, and zinc transporter activities are dependent upon the mobile reactive pools of zinc.
The next important consideration is the role of zinc transporters in cellular accumulation and distribution of zinc. The source of cellular zinc is the extracellular environment, which most typically is the interstitial fluid (ISF) derived from blood plasma. A reasonable estimate of the ISF concentration of zinc donors for transport of zinc into cells is ~6 uM; which includes Zn-Albumin ~5 μM; Zn-organic acid ~0.3 μM; free Zn2+ ions ~0.5 nM. The cellular uptake of zinc from this zinc donor pool results from the presence of ZIP-family (Slc39A) zinc uptake transporters located at the plasma membrane. Once within the cell, the zinc is distributed among the cytosol, cytoplasmic organelles, and nucleus; or exported from the cell to the extracellular fluid by ZnT-family transporters (Slc30A). ZnT-mediated transport, as with ZIP transport, involves the transport of the mobile, exchangeable pool of Zn Ligands. For example, the cytosolic concentration of the mobile, exchangeable pool of zinc is estimated to be in the range of ~1- 20 uM; and includes metallothioneins and organic acids such as amino acids and citrate, with free Zn++ ion being <1 nM.
Zinc homeostasis of normal vs. malignant cells
Zinc is an indispensible component of all cells, including normal cells and malignant cells. All cells require the maintenance of their appropriate cellular concentration, forms, and distribution of zinc that is necessary to support their growth, proliferation, metabolism, and functional activities. The normal cells evolved with homeostatic mechanisms that maintain their normal status of zinc in their natural in situ environment. Under such appropriate conditions, zinc is not cytotoxic. However, if the normal cellular status of zinc is disturbed by influences that impose compromising conditions of either increased or decreased zinc, pathophysiological cytotoxic consequences can result.
The same relationship applies to malignant cells. However, the status of zinc (such as the cellular concentration of zinc) for the activities of the malignant cell is not the same as the required zinc status of the normal cell. This is revealed by the evidence that in several cancers [2-4] the zinc concentration of malignant tissue is markedly decreased compared to the corresponding normal or benign tissue; which is further supported by the cytotoxic effects of zinc accumulation on the malignant cells in contrast to the normal cells. Therefore, the malignant cells in situ do not retain the same protective mechanisms against cytotoxic effects of zinc that exist in the normal cells. Instead, the malignant cells evolved with other in situ protective mechanisms, which include the decrease in their cellular accumulation of zinc to lower concentrations that are not cytotoxic; but are sufficient to maintain and support their growth, proliferation, metabolism and malignant activities. Involved in this adapted protective mechanism by malignant cells is the alteration of expression and abundance of zinc transporters so as to decrease zinc accumulation and to sequester intracellular zinc to minimize potential cytotoxicity.
It is also important to bring attention to a related issue that exists in many studies of zinc effects on cell growth and proliferation. It has become a common experimental procedure to deplete cells of zinc by pretreatment with cell permeable zinc chelators such as TPEN (N,N,N′,N-tetrakis(2-pyridylmethyl ethylenediamine), in order to demonstrate a positive effect of zinc treatment on promoting cell proliferation. An assumption is made that treatment with a low concentration of TPEN (formation constant of log Kf~15) for zinc binding will deplete and tightly bind only an equivalent low amount of the total cellular zinc. Such pretreatment will first tightly bind and remove the pool of exchangeable reactive zinc; and will also deplete a significant amount of the total zinc concentration of the cell. The zinc-depleted cell will not exhibit the growth, proliferation, and activities of the normal cell; and can exhibit cytotoxic effects of zinc deficiency. Consequently, a positive effect of zinc treatment under such conditions likely involves the restoration of the required cellular zinc that is essential to restore the normal activities of the cell. This does not represent the effect of exposure to zinc on a cell that exists with its normal physiological composition of zinc.
Integration of molecular genetics and proteomics with zinc transporter function
The expression, abundance, and functional activity of ZIP and ZnT transporters play a major role in the status of zinc in all cells. Consequently, the identification of specific functional zinc transporters in cells in relation to the maintenance and changes in the zinc status of the cell is an important relationship in cancer. Since a cell’s concentration of zinc is first dependent upon its uptake of zinc from its extracellular fluid environment, the plasma membrane ZIP transporters play an important role. While there exist many studies of the relative gene expression of ZIP (and ZnT) transporters, very few studies of the functional role of the specific transporter in a specific cell exists. Unfortunately, descriptions and conclusions of the specific role of transporters in specific cells have been based solely on gene expression results; which have resulted in questionable and erroneous information. To establish the functional involvement of a ZIP transporter, the abundance and localization of the transporter at the plasma membrane of the cell should be demonstrated with in situ analysis (e.g. immunohistochemistry) of the cell in its natural environment [12,13]. This also includes, when necessary, that the plasma membrane localization be further identified as being apical, basilar, or both. When these conditions are established, studies with corresponding isolated cell preparations or appropriate cell lines should be conducted to determine the kinetics of zinc uptake transport under physiological conditions; and the effects of upregulation and silencing of the transporter on zinc uptake by the cells.
The above combined studies will then permit a conclusion of the likely involvement of a specific ZIP transporter as an important functional zinc uptake transporter in the cell. It is important to caution that the identification of the transporter and its activity in cell lines, in the absence of the in vivo identification of the transporter as described above, cannot be employed to establish the status and role of the transporter in the cell in its natural environment. This is well exemplified in our prostate studies, in which we identified that ZIP1 transporter is constitutively expressed and results in zinc uptake and accumulation in malignant prostate cell lines. However, we subsequently identified that in prostate cancer, ZIP1 expression is silenced and zinc accumulation is suppressed in the malignant cells in vivo [5,14]. We observed a similar relationship regarding ZIP3 in the PANC1 cell line that was derived from pancreatic ductal adenocarcinoma [15].
The preceding descriptions hopefully provide a fundamental basic awareness and understanding of zinc and zinc transporter relationships that should be considered and applied to any and all issues that involve the implications of zinc in cells. With this background, we will proceed with the specific issue of the role of zinc in pancreatic adenocarcinoma.
The status of zinc concentration in normal human pancreas versus carcinoma
Danielsen and Steinnes in a 1970 preliminary report [16] that involved only one pancreatic cancer tissue sample suggested that a decrease in zinc level might be associated with pancreatic adenocarcinoma. To the best of our knowledge, that remained the only report of measured zinc levels in human pancreatic cancer tissue versus normal or benign pancreas, until the report of Costello et al. [15]. Given the fact that many reports of important implications of zinc in other cancers had been published during this period of forty years, it is surprising that no apparent interest or consideration had been given to a possible involvement of zinc in pancreatic cancer. This is especially so in view of the lack of identification and understanding of important factors and conditions involved in the development and progression of pancreatic cancer.
Li et al. [17] in studies with with human pancreatic cancer and normal tissue analyses reported that ZIP4 transporter is upregulated in ductal adenocarcinoma; and ZIP4 is localized at the plasma membrane. The authors presented the conclusion that, “The overexpressed ZIP4 may provide an increased zinc supply to the fast growing tumor cells, in which the zinc availability is limited”. They further concluded from experimental genetically altered malignant cells and mouse xenograft studies that “...increased cell proliferation and higher zinc content were also observed in the (mouse) tumor tissues that overexpressed ZIP4.” However, the authors provided no measurements or data relating to zinc levels in the human normal/benign versus adenocarcinoma tissue samples; and no reference to any other reported studies that provided human pancreatic exocrine zinc levels. Their subsequent reports [18-20], as best that we could determine, did not provide any determination of zinc levels in human normal and cancer tissues; yet they concluded In their most recent report in 2013 [20], “...that ZIP4 might be the major zinc transporter that is responsible to the increased cellular zinc concentration in pancreatic cancer cells, and promotes pancreatic cancer cell proliferation and tumor growth, which strongly suggests that ZIP4 plays a central and decisive role in pancreatic cancer progression...” Seemingly, their conclusion that zinc is increased in human pancreatic ductal adenocarcinoma, is derived from their experimental studies with cell lines and tumorigenic animal models; and is not confirmed by direct measurements of zinc in human pancreatic tissues.
Pursuant to the initial reports of Li et al. Costello et al. determined the relative zinc levels in human normal/benign pancreatic tissue versus adenocarcinoma [15,21]. The studies employed in situ zinc staining with DTZ (dithizone) of human pancreatic tissues to determine if apparent changes exist in the zinc levels of normal/benign pancreas versus carcinoma; and if so, which specific cells are associated with the changes. This approach is advisable due to the complexity of the pancreas being comprised of an endocrine component and an exocrine component; and the fact that the islet cells (especially the beta cells) contain uniquely high zinc levels. Under such conditions, assay of zinc levels in pancreatic tissue extract preparations will not be specific for the exocrine normal epithelium and malignant components of the pancreas. DTZ was employed for the in situ determination of relative zinc levels in cells because it has a high Zn-binding affinity (formation constant log Kf~15) that provides an estimate of total zinc (i.e. exchangeable zinc plus strongly bound immobile zinc).
The following description provides representative results of the Costello et al. studies, and the reader is referred to their reports [15,21] for the original extensive and specific data and discussions. The initial study [15] was conducted with pancreatic tissue microarray slides obtained from U.S. Biomax and included a total of duplicate cores of Grade 1 (17 cases); Grade 2 (18 cases); Grade 3 (12 cases); and 10 normal cases. Figure 1 shows that the ductal and acinar epithelium in normal pancreas exhibit relatively abundant DTZ-detectable zinc. In contrast, zinc is uniformly markedly decreased in the ductal and acinar epithelium of well differentiated (Grade 1) malignancy; and this loss of zinc persists through advancing malignancy. We did not observe any case in which the zinc level in adenocarcinoma was increased in comparison to the normal pancreatic ductal and acinar epithelium. Zinquin (formation constant log Kf~11) staining confirms the loss of zinc, and demonstrates the exchangeable reactive pool of zinc is significantly decreased in the malignant cells compared to the normal epithelium. Also, the high zinc levels associated with the islet cells was retained and unchanged in the normal versus cancer tissues (not shown); so that the loss of zinc is specific for the adenocarcinoma.
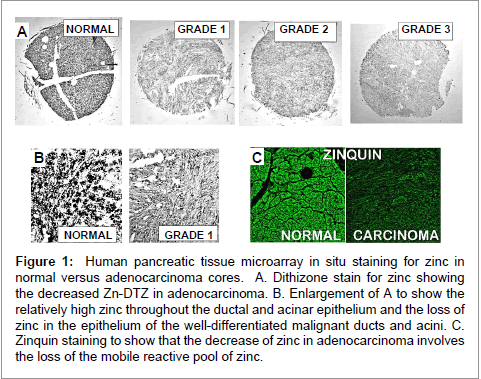
Figure 1: Human pancreatic tissue microarray in situ staining for zinc in normal versus adenocarcinoma cores. A. Dithizone stain for zinc showing the decreased Zn-DTZ in adenocarcinoma. B. Enlargement of A to show the relatively high zinc throughout the ductal and acinar epithelium and the loss of zinc in the epithelium of the well-differentiated malignant ducts and acini. C. Zinquin staining to show that the decrease of zinc in adenocarcinoma involves the loss of the mobile reactive pool of zinc.
The pursuant Costello et al. report [21] with archived human tissues in addition to TMAs focused on the early development of pancreatic malignancy. The results as represented in figure 2 further corroborated the consistent marked loss of zinc that exists in early malignancy. It is evident that the epithelium of the well-differentiated ducts and acini exhibit the loss of zinc. In addition, the low zinc level in adenocarcinoma is also evident in the epithelium of low grade and high grade PanINs. It is notable that the low zinc exists in PanIN lesions throughout the transdifferentiation of the early PanIN epithelial cells to the development of the putative malignant ducts. The consistent results of these studies lead to our conclusion that zinc is decreased in the development and progression of malignancy in human pancreatic adenocarcinoma.

Figure 2: Dithizone staining for relative zinc levels in human benign versus adenocarcinoma tissue sections. A and B show the loss of zinc in the ductal and acinar epithelium in Grade 1 well-differentiated adenocarcinoma. C shows that the columnar epithelium of low grade PanINs contains low zinc as does the surrounding ductal adenocarcinoma. D and E show the H&E and DTZ staining of high grade PanIN3 to demonstrate that zinc is low throughout the transdifferentiation of the epithelial cells to the formation of malignant ducts.
The status of relevant zinc transporters in normal human pancreas versus carcinoma
Keeping in mind that gene expression information, in itself, does not establish the status and functional relationship of transporters in cells, there exists little information regarding the status of ZIP and ZnT transporters in human normal and adenocarcinoma pancreatic exocrine tissues. Relative to ZIP family zinc uptake transporters, Li et al. [17] published the first report that identified the presence of a ZIP transporter (ZIP4) in human pancreatic exocrine cells (i.e. in ductal adenocarcinoma) and this was corroborated by Wang et al. [20]. They employed RT-PCR analysis of RNA extracts of human adenocarcinoma and adjacent normal/benign tissues to identify an increase in ZIP4 expression in adenocarcinoma; which they confirmed with ZIP4 IHC to demonstrate an increase in abundance of plasma membrane associated ZIP4 transporter in ductal adenocarcinoma. As described above, an association of the increased abundance of plasma membrane ZIP4 with a corresponding increase in zinc accumulation (or any zinc determinations) in the human pancreatic tissues, was not provided or demonstrated. Instead, they used the results of ZIP4 genetic engineered pancreatic cells and mouse xenograft tumors to reach their conclusion that “the overexpressed ZIP4 absorbed more zinc, and an increased amount of zinc is necessary for pancreatic cancer progression.”
Wang et al. [20] reported the results of the analyses of relative gene expressions of ZIP and ZnT members in RNA extracts of pancreatic tumor and surrounding benign tissues; which lead them to conclude “... that all 13 ZIPs other than ZIP4 were down-regulated in the pancreatic tumors compared with the surrounding benign tissues... ZIP4 is the only zinc transporter that is significantly up-regulated in human pancreatic cancer and might be the major zinc transporter that plays an important role in pancreatic cancer growth.” No description is presented of methods employed to minimize the heterogeneity of the cell populations in the resected tissue samples, which, if not done, results in a mixture of RNA’s from different cells. Does one expect that a cell requires or employs the functional activity of 13 ZIP transporters to achieve its uptake of zinc? Also, why should one expect that an increase of zinc uptake due to ZIP4 upregulation would negate the downregulation effects of 13 other (or any number) functional ZIP transporters, if they were all associated with the same cells? Unfortunately, these issues raise question regarding the approach and the conclusions represented in that report.
To identify the possible ZIP transporter(s) associated with the observed decrease in zinc in adenocarcinoma, Costello et al. [15,21] employed IHC analyses of the human pancreatic TMAs and archived human tissue sections described in the zinc studies above. Figure 3 shows that ZIP3 transporter is abundant in the normal ductal/ acini epithelium. Most importantly, the ZIP3 transporter is localized predominantly with the basal plasma membrane of the normal ductal/ acinar epithelium. In contrast, detectable ZIP3 transporter is virtually absent in early well-differentiated malignancy and in the advancing malignancy. In addition, it became evident that the islet beta cells also exhibit ZIP3 transporter; the abundance of which did not change in normal versus adenocarcinoma tissue. This demonstrates that the IHC loss of ZIP3 is specific for exocrine carcinoma. In addition to the IHC analyses, Costello et al. also showed [15] with in situ RT-PCR that ZIP3 gene expression that exists in normal ductal/acinar epithelium is down-regulated in adenocarcinoma; which is the likely cause of the loss of transporter.
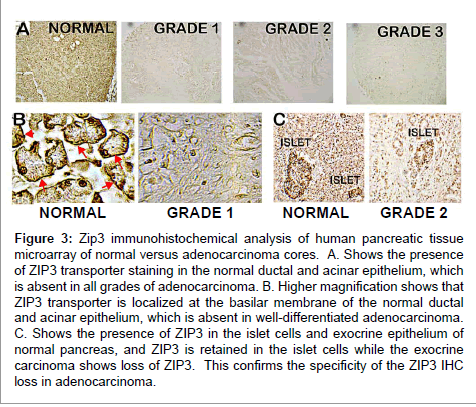
Figure 3: Zip3 immunohistochemical analysis of human pancreatic tissue microarray of normal versus adenocarcinoma cores. A. Shows the presence of ZIP3 transporter staining in the normal ductal and acinar epithelium, which is absent in all grades of adenocarcinoma. B. Higher magnification shows that ZIP3 transporter is localized at the basilar membrane of the normal ductal and acinar epithelium, which is absent in well-differentiated adenocarcinoma. C. Shows the presence of ZIP3 in the islet cells and exocrine epithelium of normal pancreas, and ZIP3 is retained in the islet cells while the exocrine carcinoma shows loss of ZIP3. This confirms the specificity of the ZIP3 IHC loss in adenocarcinoma.
Supporting evidence is provided by the results obtained with archived tissue sections that focused on the early development of malignancy [21]. Figure 4 corroborate the abundance of ZIP3 at the basilar membrane of the normal ductal/acinar epithelium; which is absent in well-differentiated and advanced adenocarcinoma, and in the epithelium of early and advanced PanINs. It is especially notable that the same high grade PanIN 3 lesion that contained loss of zinc throughout the transdifferentiation to malignant ducts (Figure 2), also shows concurrent absence of ZIP3. The localization of ZIP3 at the plasma membrane of the normal epithelium and the consistent loss of ZIP3 concurrently with the loss of zinc provides strong evidence that ZIP3 is the likely functional zinc uptake transporter; which, when down-regulated, results in loss of zinc in the premalignant and malignant cells in pancreatic adenocarcinoma.
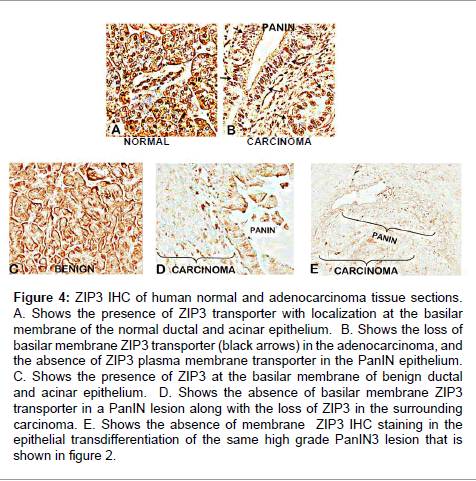
Figure 4: ZIP3 IHC of human normal and adenocarcinoma tissue sections. A. Shows the presence of ZIP3 transporter with localization at the basilar membrane of the normal ductal and acinar epithelium. B. Shows the loss of basilar membrane ZIP3 transporter (black arrows) in the adenocarcinoma, and the absence of ZIP3 plasma membrane transporter in the PanIN epithelium. C. Shows the presence of ZIP3 at the basilar membrane of benign ductal and acinar epithelium. D. Shows the absence of basilar membrane ZIP3 transporter in a PanIN lesion along with the loss of ZIP3 in the surrounding carcinoma. E. Shows the absence of membrane ZIP3 IHC staining in the epithelial transdifferentiation of the same high grade PanIN3 lesion that is shown in figure 2.
Costello et al. [15] also conducted IHC analysis of ZIP1 and ZIP2; which showed that both transporters exist in normal ductal/acinar epithelium; and both are decreased in adenocarcinoma. However, unlike ZIP3, there was no demonstrable association of ZIP1 or ZIP2 with the ductal/acinar epithelial plasma membrane. This indicates that they are not functional transporters for cellular zinc uptake by the normal ductal/acinar epithelium; and are not likely associated with the loss of zinc in adenocarcinoma. In addition this also demonstrates that RT-PCR gene expression studies alone cannot identify functional transporter; and can result in misinterpretation of the relationship of the transporter.
In prostate cancer, the decrease in zinc is due to the downregulation of ZIP1gene expression, which is regulated by RREB1 [22,23]. Therefore, Costello et al. [21] determined the status of RREB1 in relation to ZIP3 in TMAs and archived tissues of human normal pancreas versus carcinoma. They found that RREB1 is abundant in the normal ductal and acinar epithelium; and is lost, along with ZIP3, in well-differentiated and advanced carcinoma, and in the epithelium of the PanINs (Figure 5). This is suggestive of RREB1 regulation of ZIP3 expression, and that down-regulation of RREB1 might be the cause of silencing of ZIP3 gene expression in the development of pancreatic carcinoma. However, in prostate cancer, RREB1 is a negative modulator of ZIP1 expression; whereas RREB1 appears to be a positive modulator in pancreas. It is known that RREB1 regulation of gene expression as a positive or negative modulator is dependent on the specific cell type and the specific gene [24]. Further studies are required to establish that RREB1 downregulation leads to ZIP3 silencing in pancreatic cells.
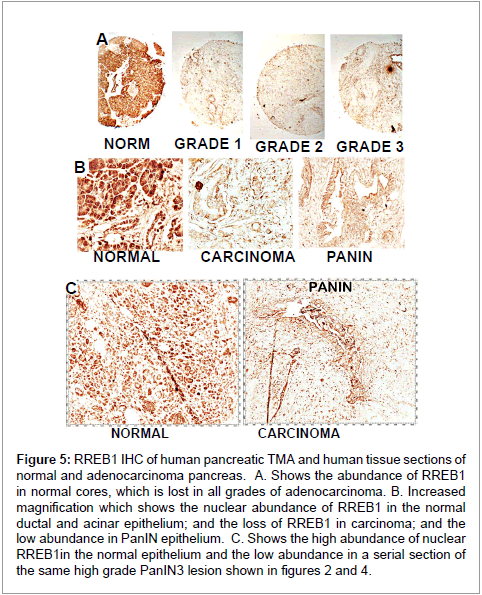
Figure 5: RREB1 IHC of human pancreatic TMA and human tissue sections of normal and adenocarcinoma pancreas. A. Shows the abundance of RREB1 in normal cores, which is lost in all grades of adenocarcinoma. B. Increased magnification which shows the nuclear abundance of RREB1 in the normal ductal and acinar epithelium; and the loss of RREB1 in carcinoma; and the low abundance in PanIN epithelium. C. Shows the high abundance of nuclear RREB1in the normal epithelium and the low abundance in a serial section of the same high grade PanIN3 lesion shown in figures 2 and 4.
The effects and role of zinc in pancreatic carcinoma and other cancers
The preceding description seemingly presents two opposing views of the effects and role of zinc in the development and progression of pancreatic carcinoma. Although they do not demonstrate increased zinc in human pancreatic adenocarcinoma, Li et al. [17-20] present the view that that zinc is increased in pancreatic carcinoma in order to promote the growth and proliferation of the malignant cells. Their conclusion is derived from experimentally engineered malignant pancreatic cell lines and mouse xenograft studies to show that increased zinc was associated with the promotion of tumor growth. They also reported that zinc treatment of zinc-depleted (TPEN pretreatment) malignant cells with overexpressed ZIP4 increased zinc accumulation, which increased cell proliferation. However, the adverse effects of TPEN zinc depletion on cell growth and the positive effect of required zinc restoration are not considered. They express the general conclusion [20] that “Overexpression of zinc transporters in cancers may regulate the expression of many other genes or transcription factors by providing sufficient zinc ions to the fast growing tumor cells.”
The opposing view from the studies of Costello et al. is that the decreased zinc in human pancreatic adenocarcinoma exists to prevent its potential in situ cytotoxic effects on the development and progression of the malignant cells. Reports of others [25,26] have shown that zinc treatment of malignant pancreatic cells inhibits cell proliferation. Also, Costello et al. [15] showed that PANC1 cells (a cell line derived from pancreatic ductal adenocarcinoma) exhibit constitutive plasma membrane ZIP3 transporter (Figure 6). Exposure of the cells to physiological concentrations of zinc results in significant inhibition of cell growth. Moreover, most cancers for which studies are available exhibit a decrease in zinc level, a cytotoxic/growthinhibitory effect of zinc on the malignant cells, and a down-regulation of corresponding ZIP zinc uptake transporters. This applies to prostate cancer, hepatocellular cancer, ovarian cancer, choriocarcinoma, colon cancer, and others [2-4,27-30]. The exception is breast cancer in which zinc is increased [31]. However, evidence exists that the cytoplasmic zinc is sequestered into vesicles by ZnT2, which prevents the cytotoxic effects in the malignant breast cells [31,32]. Nevertheless, contrary to the conclusion expressed by Li et al. the more appropriate generalization is that zinc is decreased in most cancers; and this occurs to prevent the potential cytotoxic effects of zinc on the malignant cells.
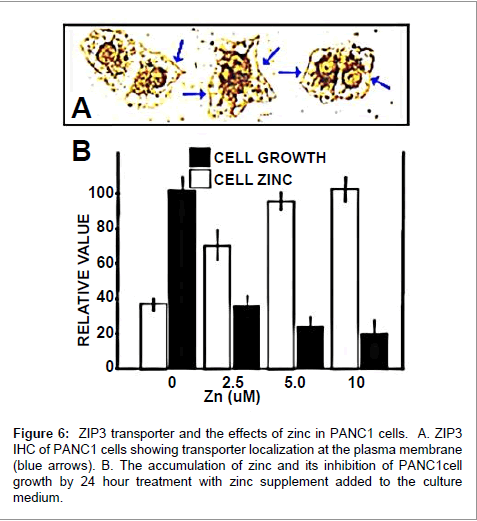
Figure 6 : ZIP3 transporter and the effects of zinc in PANC1 cells. A. ZIP3 IHC of PANC1 cells showing transporter localization at the plasma membrane (blue arrows). B. The accumulation of zinc and its inhibition of PANC1cell growth by 24 hour treatment with zinc supplement added to the culture medium.
Proposed concepts of the implications of zinc in pancreatic carcinogenesis
Presently, the origin of pancreatic ductal adenocarcinoma is unknown; especially the transformation of the normal cells that give rise to the premalignant cells leading to the malignant cells. A prevailing view is that PanINs, especially PanIN 2 and PanIN 3, are precancer lesions that give rise to ductal adenocarcinoma [33-35]; and the transdifferentiation of PanIN epithelium to ductal adenocarcinoma as described by Scarlett et al. [35] is shown in figure 7. Costello et al. [15,21] showed that the concurrent decrease in RREB1, ZIP3, and zinc is evident in all stages of PanIN development through welldifferentiated and advanced stage malignancy (Figure 7). This zinc relationship would be supportive of the close relationship between PanIN and the development of ductal adenocarcinoma.
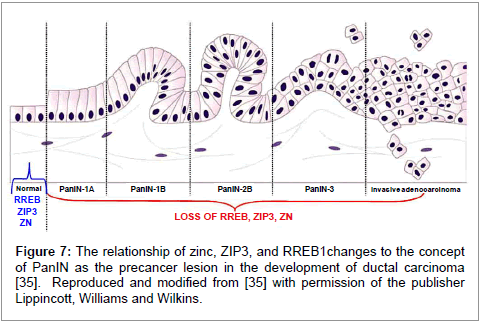
Figure 7: The relationship of zinc, ZIP3, and RREB1changes to the concept of PanIN as the precancer lesion in the development of ductal carcinoma [35]. Reproduced and modified from [35] with permission of the publisher Lippincott, Williams and Wilkins.
It is also notable that the acini epithelial cells also exhibit the concurrent loss of RREB1, ZIP3, and Zn along with the ductal epithelium in well-differentiated ductal adenocarcinoma, and with PanIN. This would seem to demonstrate a close association between the acinar cells and the development of ductal malignancy. Evidence from animal models indicates that the acini can give rise to ductal neoplasia; and the differentiated acinar cells might give rise to lowgrade PanINs [36-38]. The zinc events would support the concept that the acini epithelial cells might be the “cells of origin” leading to PanIN development and subsequent ductal carcinoma; or directly leading to ductal neoplasms. Although such relationships at this time are highly speculative, the zinc relationship provides new insight into the origin of ductal adenocarcinoma.
The consistent decrease in zinc that is apparent in the PanIN lesions, well-differentiated malignancy, and progressing malignancy leads to the likely conclusion that cellular zinc levels that exist in the normal ductal/acinar epithelium are potentially cytotoxic in malignant cells. Consequently, the pancreatic malignant cells evolved with specialized mechanisms that prevent the accumulation of cytotoxic levels of zinc. One such mechanism is the down-regulation of the functional zinc uptake transporter, ZIP3; which decreases the uptake and accumulation of zinc. This is supported by the marked decrease in ZIP3 transporter which occurs as a result of silencing of ZIP3 gene expression concurrent with the loss of zinc; and which is evident as an early event in the premalignant and malignant cells. We believe that this concept is supported by the consistent loss of ZIP3 and zinc in PanIN and malignant cells; which would suggest a generalization that malignant pancreatic cells that retain the increased zinc levels as found in the normal epithelial cells do not exist. This relationship parallels the well established role of zinc that has been demonstrated in prostate cancer, in which ZIP1 is the functional zinc uptake transporter that is down-regulated in order to decrease zinc accumulation and its cytotoxic effects in the malignant cells; and prostate cancer showing high zinc in the malignant loci virtually does not exist.
The concurrent down-regulation of RREB1 and ZIP3 suggests that RREB1 might regulate ZIP3 gene expression. In prostate cells, the human ZIP1 gene promoter region contains RREB1 binding sites that regulate its expression [22,23]. However RREB1 exhibits suppression of ZIP1 gene expression; so that RREB1 is upregulated in prostate cancer, which silences ZIP1 expression and results loss of zinc accumulation in the malignant cells. Pancreatic cells appear to present a different relationship in which RREB1 might be a positive modulator of ZIP3 gene expression. RREB1 has been shown to function either in the induction or repression of gene expression depending upon the cell type and the specific gene; and RREB1 is emerging as a human oncogene that might suppress or promote carcinogenesis and tumor progression [24]. Thus the silencing of RREB1 in relation to ZIP3 downregulation and loss of zinc could represent one of the earliest events in pancreatic oncogenesis.
Potential clinical relevance of the zinc relationship in pancreatic adenocarcinoma
The preceding discussion indicates that the zinc relationship can have important implications for the treatment and diagnosis of pancreatic cancer. The fatal outcome of pancreatic cancer is due to the absence of any effective chemotherapeutic agent or approach for treatment once the cancer has been identified in a subject; and there exists no effective biomarker for the identification of early malignancy or premalignant at-risk individuals. It is plausible to expect that a treatment regimen that will promote the uptake and accumulation of zinc in the malignant cells will result in the cytotoxic effects and will abort the malignancy. However, such an approach to be successful must take into consideration important relationships of zinc as we have described [2]. The idea that a regimen that will increase the concentration of zinc delivered to the malignant site (such as increased dietary or supplemental zinc to increase plasma zinc concentration) will be effective is an erroneous expectation. The down-regulation of the functional ZIP transporter in the malignant cells prevents the uptake and accumulation of zinc. An approach to circumvent this problem is the potential use of a zinc delivery process such as a zinc ionophore that results in the permeation of zinc across the plasma membrane of the malignant cells; and enters the cytosol as an exchangeable reactive form of zinc. If successful, such an approach might also be employed to prevent the development of malignancy in at-risk subjects with premalignant lesions. Since the loss of RREB1, ZIP3, and zinc occurs in PanIN and early stage malignancy, this could provide the potential of being effective biomarkers for early detection of malignancy or premalignant conditions. Thus, the zinc relationship has potential in the diagnosis, treatment and prevention of pancreatic adenocarcinoma.
Pancreatic cancer continues to be an untreatable deadly cancer with no currently available early diagnostic procedure or marker. The factors and conditions that lead to development of pancreatic cancer are still largely unknown. Alterations of the status of zinc has been identified and implicated in several cancers. Consideration and studies of a potential role of zinc in pancreatic cancer had received little attention until recently. This presentation describes and reveals the recent evidence that zinc is an important factor in the development and progression of pancreatic adenocarcinoma. Consequently, focused attention and required clinical and experimental research into the implications of zinc and zinc transporters is of paramount importance. Correspondingly, such research must be conducted with conditions and understanding of the physiological and biochemical relationships of zinc in mammalian systems; and the appropriate integration of molecular genetics, proteomics, and experimental cell and animal models. This is essential to provide valid and accurate interpretations and translational application of the involvement of zinc and zinc transporters in human normal pancreas and adenocarcinoma.
This review reveals some seemingly divergent views of the status of zinc and zinc transporters associated with pancreatic cancer; which, over time with pursuant research, will be reconciled. Despite this, the unanimity is that the currently available studies provide evidence that zinc is an important factor in pancreatic cancer,
Based on the evidence from the studies with human pancreatic tissue, we provide a new concept for the development of pancreatic cancer. The concept proposes that zinc is decreased in pancreatic cancer in order to eliminate its cytotoxic effects on the malignant and premalignant cells in situ; and this is achieved by the silencing of expression of ZIP3, the functional zinc uptake transporter. This is an early and required event in pancreatic carcinogenesis. We present this concept with the hope that it will promote, stimulate and motivate the interest and research support and involvement of other biomedical clinical investigators.
The studies of the authors (LCC and RBF) described in this review were supported by NIH grants CA79903, CA93443, and DK42839.