Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 2
The genetic manipulation in filamentous fungi has always remained a challenge, which is necessary to study gene functions in pathogenic fungi so as to gain insight into the molecular processes necessary for pathogenesis. The fulfillment of this objective depends on the availability of suitable transformation method. Fusarium oxysporum is a plant pathogenic filamentous fungus, which causes vascular wilt disease in a wide variety of plants. The current transformation methods used for this fungus are particle bombardment, Agrobacterium tumefaciens- and protoplastmediated transformation. In this study, we have developed an easy, rapid and reproducible protocol for transformation of F. oxysporum by using glass beads in combination with polyethylene glycol. We utilized the GFP gene construct for developing this transformation protocol. The transformation frequency of about 15 transformants per μg of DNA was obtained. This simple transformation method may be adopted for other filamentous fungi.
<Keywords: Fusarium oxysporum, Transformation, Glass beads, Agrobacterium-mediated transformation, GFP
Fusarium oxysporum f.sp lycopersici is a soil borne phytopathogenic fungus of economic significance which infects tomato leading to symptoms typical of vascular wilting [1]. F. oxysporum mediated disease development process has been extensively studied. This fungus colonizes the root cortex and proliferates within the xylem vessels leading to characteristic disease symptoms like vein clearing, leaf epinasty, wilt, defoliation and ultimately host plant death [1]. It has also been reported as an emerging human pathogen, causing opportunistic mycoses [2,3]. The genome of this fungus has been sequenced recently by the Broad Institute and has provided with large amount of genetic information that has to be decoded so as to apply the valuable information to understand the process of pathogenesis. This in turn would strengthen our approach towards devising the novel strategies for disease control. The important consideration for applying any genetic engineering approach in an organism is the requirement of an efficient transformation method.
The commonly available transformation methods to achieve the delivery of exogenous DNA in fungi are polyethylene glycol-mediated transformation using protoplasts; Agrobacterium tumefaciens mediated transformation and electroporation method [4]. The successful protoplast-mediated transformation of Saccharomyces cerevisiae was first reported in the year 1979 [5]. Later, the use of protoplasts has been extended to several filamentous fungi [6,7]. However, there are many fungi which do not yield sufficient protoplasts or whose protoplasts do not regenerate efficiently. Therefore, alternative methods of fungal transformation were developed, which are exempted from the use of protoplasts. These methods include, biolistic transformation [7] and Agrobacterium- mediated Transformation (AMT) [8] are the commonly used methods, which work well with most of the fungi.
Importantly, implementing a particular transformation strategy depends on the fungal strain and needs to be optimized to achieve the significant transformation efficiency [4]. Although AMT method is the commonly available method for transformation of F. oxysporum, here we report an alternative novel transformation system for this fungus. This method uses glass beads to deliver DNA molecules in combination with polyethylene glycol. It is a simple, rapid and cost-effective method that worked efficiently in F. oxysporum. In this study, we utilized the reporter gene, GFP under the control of cauliflower mosaic virus (CaMV) 35S constitutive promoter for optimizing the transformation procedure. In addition, we also transformed the fungus with GFP gene by AMT method, which enabled us to compare the two methods of transformation. Although, the transformation efficiency achieved with AMT and glass bead transformation methods were high, the glass bead method yielded relatively higher transformation frequency with higher fluorescence intensity in most of the fungal transformed isolates as compared to AMT method.
Fungal spore collection and germination
F. oxysporum was maintained on solid PDA medium and after 7 days the spores were isolated by pouring sterile distilled water (5-6 ml) to the plate and scrapping it gently with sterile loop to release the spores. The resulting spore suspension was filtered through two layers of sterile cheese cloth to remove any mycelial fragments and the conidial suspension was centrifuged at 4,500 rpm, 4°C for 5 min. The spores were washed thrice with sterile water to remove the spore mucilage and finally resuspended in sterile water at a concentration of 1×106 spores/ml. The spores were germinated for 6 h in PDA broth at 28°C at 100 rpm.
Transformation of F. oxysporum with GFP gene
Transformation of F. oxysporum was done with pCAMBIA 1302 vector harboring sGFP gene under the control of CaMV35S promoter and hygromycin B phosphotransferase (HPT) as the selection marker gene (generously provided by Dr. Richard A. Jefferson, CAMBIA, Canberra, Australia). Two different protocols were followed- a) Agrobacterium-mediated transformation (AMT) method [9] and, b) glass-bead method.
Agrobacterium-mediated transformation
F. oxysporum spores were germinated in carboxy methyl cellulose (CMC) broth (15 g carboxymethyl cellulose, 1 g yeast extract, 0.5 g MgSO4, 1 g NH4NO3 and 1 g KH2PO4, for 1 litre broth) at 100 rpm, at 28°C for 1 week. The culture was filtered through two layers of cheese cloth and spores were harvested at 4,500 rpm, 4°C for 5 min. Using haemocytometer, spores were counted and diluted to 106 spores/ml of spore suspension.
Liquid culture of Agrobacterium strain LBA4404: pCAMBIA 1302 was prepared by inoculating 7 ml LB supplemented with 50 μg/ ml kanamycin and 50 μg/ml streptomycin with the glycerol stock of the strain and grown at 250 rpm, 28°C over night. The remaining transformation protocol was followed exactly as per the protocol standardized. The putative transformants were visible after 5-6 days as rapidly growing circular colonies. The potential transformants were transferred to fresh selection plates.
Glass bead transformation
F. oxysporum spores were isolated and spores were germinated by shaking at 100 rpm for 6 h at 28°C. Glass beads (0.45-0.52 in diameter) were washed with conc. H2SO4, then rinsed with sterile distilled water and sterilized by baking at 250°C for 2-3 h. 300 mg of sterile glass beads were added to 400 μl of spore suspension (106 spores/ml) along with 1 μg of linearized pCAMBIA1302 plasmid DNA. Finally, freshly prepared PEG (MW 3500; Sigma, USA) at final concentration of 5% w/v was added immediately before transformation along with 0.1 M lithium acetate. The transformation mix was agitated briefly at full speed on vortex in 15 ml polypropylene tubes. Beads were allowed to settle down and liquid suspension was placed on solid growth medium supplemented with 200 μg/ml hygromycin in Petri plates overlaid with nitrocellulose membrane and circular colonies were obtained in 6-7 days.
Genomic DNA isolation and PCR analysis of fungal transformants
Genomic DNA was isolated from fungal transformants by CTAB method [10]. The presence of the transgene was determined by PCR analysis for which about 50-100 ng of DNA from untransformed control and putative transformants was added separately to a PCR reaction mix tube consisting of 100 nM of forward and reverse primers specific to transgene of interest, 1X PCR buffer, 2 mM MgCl2, 100 μM dNTP mix and 0.5 U of Taq DNA polymerase (Biotools, Spain), the volume was made up to 25 μl with sterile double distilled water. The PCR conditions were set as follows: initial denaturation at 94°C for 5 min, followed by 30 cycles of 30 sec denaturation at 94°C, 1 min and primer annealing at appropriate temperature and 30 sec of extension at 72°C, with a final extension of 72°C for 10 min. The PCR products were analyzed on a 1 % agarose gel. The sequence of primers and the respective annealing temperatures used were as follows-F.P: 5’ ATT CTA GAG GCG CGC CAC TGG 3’, R.P: 5’ GCG GAT CCA TTT AAA TGT CTT 3’ at 59°C for checking GFP integration and F.P: 5’ CGC ATG AAA AAG CCT GAA CTC ACC GCG 3’, R.P: 5’ GCA GGC TCC CGT TTC CTT ATC GAT 3’ at 52°C to study HPT gene integration.
RT- PCR analysis to study transgene expression
Semi-quantitative RT-PCR was performed to check transcripts of the GFP transgene in fungal transformants. Prior to RT-PCR, RNA was isolated using Trizol reagent (Invitrogen, USA) and treated with DNase (Taurus scientific) as per the manufacturer’s protocol. DNaseI treated RNA was quantified using Nanodrop (Thermoscientifc, USA) and total RNA at the concentration of 500 ng was used as the template for RTPCR. One step RT-PCR kit (Bioscript) was used for this purpose. RNA template was mixed with reaction mix consisting of 1x RT-PCR buffer, one step enzyme mix (1 μl), forward primer and reverse primer (200 nM), RNase inhibitor (5 units) and remaining volume was made up with DEPC treated water to 25 μl. The thermal cycler was programmed as follows: initial reverse transcription at 42°C for 30 min, RT enzyme denaturation at 95°C for 10 min followed by 30-35 cycles of cDNA amplification.
The relative fluorescence intensity was measured in the germinating spores (104 spores/ml) of the transformed isolates using spectrofluorometer at 460-500 nm excitation wavelength and emission wavelength of 510-560 nm. For detection of GFP fluorescence by using confocal microscopy, the small amount of untransformed control and GFP transformed fungal tissue were taken separately on a glass slide in a drop of water. After placing the cover slip, the slides were sealed properly and the fluorescence was visualized under the microscope using GFP filters (excitation- 489 nm and emission-511 nm).
Transformation of F. oxysporum using glass beads and AMT method
The pCAMBIA 1302 vector carrying sGFP gene under the control of CaMV35S promoter and with hygromycin resistance gene, HPT as the selection marker gene (Figure 1) was utilized to transform F. oxysporum. The transformation of the fungus with GFP gene using glass beads yielded 15 transformants per μg of plasmid DNA. Although the original GFP gene from Aquorea victoria has been expressed in many organisms, it has been difficult to obtain detectable expression of this gene in filamentous fungi [11]. However, a modified version of the GFP gene, sGFP, where serine 65 was replaced with a threonine [12], has been successfully expressed at high levels in a number of filamentous fungi [11,13-15]. Interestingly, sGFP does not interfere with the fungal development process, therefore, can be utilized for studying disease infection process in host plants [13]. Due to these reasons original mGFP gene from the expression vector pCAMBIA 1302 was replaced with sGFP gene.
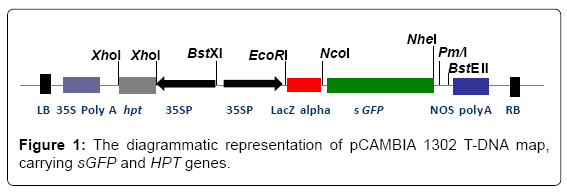
Figure 1: The diagrammatic representation of pCAMBIA 1302 T-DNA map, carrying sGFP and HPT genes.
The transformants obtained by AMT method appeared as single circular colonies on the selection medium plate supplemented with 200 μg/ml hygromycin. Each of the colonies were picked and cultured individually on PDA medium fortified with hygromycin. The phenotype of the transformants was normal when compared with wild-type fungus. In case of the glass bead transformation, the transgenes were directly introduced into the genome of the fungus without the need of Agrobacterium. Brief vortexing of glass beads might have created the temporary pores in the fungal cell wall which facilitate the uptake of the DNA molecules and the pores created must have repaired after some time. The addition of PEG further improved the DNA uptake efficiency. These results are in line with the previous report in yeast [16].
Transformation in F. oxysporum was also done by AMT method as per the standard protocol [9], which resulted in 10 transformants/ μg of DNA. The transformed isolates obtained by the two methods of transformation exhibited stable GFP expression after several successive transfers onto nutrient media without antibiotic selection.
Optimization of conditions for glass bead transformation
The important factor considered while applying glass bead method for fungal transformation was the size of the glass beads. The glass beads with 0.45-0.52 mm diameter were found to be suitable for transformation. The ratio of number of spores to the amount of glass beads is also crucial, and the ratio of 1:1 yielded better transformation efficiency. Prior to transformation, the plasmid DNA was linearized with EcoRI restriction enzyme. Polyethylene glycol (PEG 3500) was added to the transformation mixture at a final concentration of 5% wt/ vol and 0.1 M lithium chloride was also added to facilitate the DNA uptake. The use of PEG results into clumping of treated cells which facilitate the trapping of DNA [7], whereas the use of high concentration of lithium ions renders the cell walls permeable to DNA without the need for protoplasts, a method initially applied for transforming yeast, S. cerevisiae [17 ].
Molecular characterization of the fungal transformants
Genomic DNA was isolated from the transfromants and PCR analysis was done to study the presence of transgenes using GFP and HPT gene specific primers, and the expected amplicon size of 500 bp was obtained (Figure 2). The GFP expression in the transformants was checked by semi-quantitative RT-PCR using GFP primers. Some of the transformants obtained by AMT method showed high GFP expression like Fo.AT3, 4 and 5 as compared to other transformants in which the expression ranged from low to very negligible (Figure 3A). The transformants obtained by glass bead method were also analyzed for GFP expression and transformants Fo.G5, 6, 7 showed high GFP expressions, while Fo.G3 and 4 were low expressing transformants and some like Fo.G1, 2 and 8 did not exhibit any expression (Figure 3B). The variation in GFP expression in different transformed isolates may be due to the position effect of the transgene integration and dosage effect of the transgene copy number [18,19].
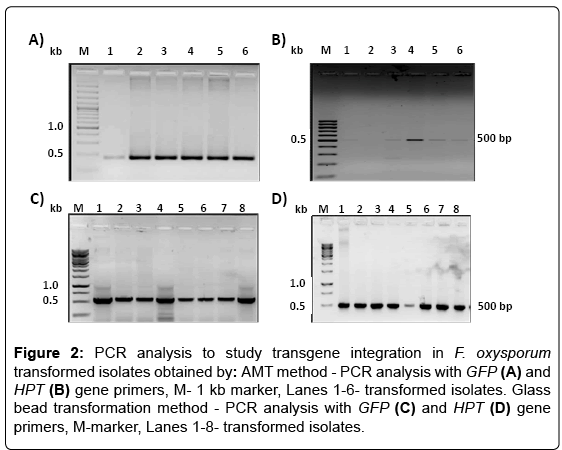
Figure 2: PCR analysis to study transgene integration in F. oxysporum transformed isolates obtained by: AMT method - PCR analysis with GFP (A) and HPT (B) gene primers, M- 1 kb marker, Lanes 1-6- transformed isolates. Glass bead transformation method - PCR analysis with GFP (C) and HPT (D) gene primers, M-marker, Lanes 1-8- transformed isolates.
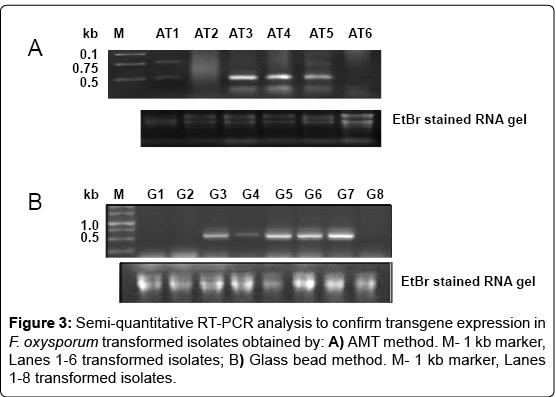
Figure 3: Semi-quantitative RT-PCR analysis to confirm transgene expression in F. oxysporum transformed isolates obtained by: A) AMT method. M- 1 kb marker, Lanes 1-6 transformed isolates; B) Glass bead method. M- 1 kb marker, Lanes 1-8 transformed isolates.
Intensity of GFP fluorescence in fungal transformants
The advantage of using GFP as a marker gene is that its fluorescence intensity is directly proportional to the amount of GFP accumulated and does not interfere with any of the endogenous proteins. To assess the level of fluorescence intensity in each of the transformed isolates, the germinating spores (104 spores/ml) were taken and tested for fluorescence using spectro-fluorometer at 460-500 nm excitation wavelength and 510-560 nm emission wavelength. Some of the transformants obtained by AMT method showed high fluorescence intensity like Fo.AT3, Fo.AT4 and Fo.AT5, while some had relatively low intensity, i.e. Fo.AT1, Fo.AT2 and Fo.AT6 (Figure 4A). Of the transformants obtained by glass bead method, Fo.G5, 6 and 7 had high fluorescence intensity as compared to other transformed isolates (Figure 4B). These results were in accordance with the results of transgene expression as analyzed by RT- PCR.
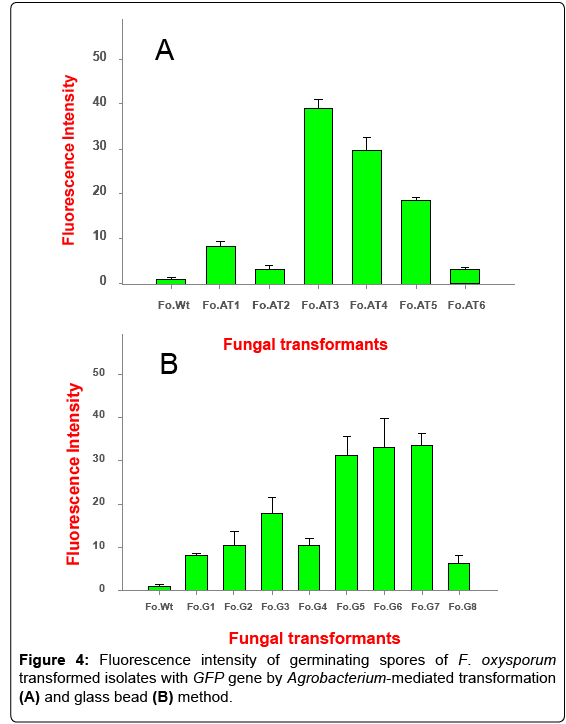
Figure 4: Fluorescence intensity of germinating spores of F. oxysporum transformed isolates with GFP gene by Agrobacterium-mediated transformation (A) and glass bead (B) method.
Visualization of GFP expression in fungal transformed isolates using confocal microscopy
The GFP expression in fungal transformed isolates was further analyzed by detection of GFP fluorescence using confocal microscopy. The fungal hyphae as well as spores were analyzed for fluorescence, and bright GFP fluorescence was observed in mycelial tissue as well as spores of the transformed isolates and the fluorescence was found to be more concentrated in the vacuoles and septae of spores (Figure 5). The GFP fluorescence was not detected in the untransformed control. The accumulation of GFP occurs in fungal cytoplasm and this is in accordance to the observations reported with other fungi [11,20,21].
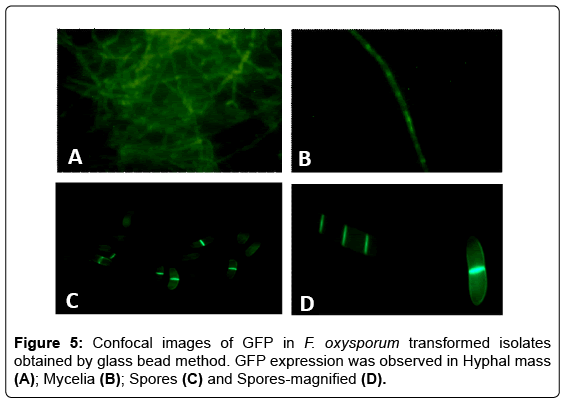
Figure 5: Confocal images of GFP in F. oxysporum transformed isolates obtained by glass bead method. GFP expression was observed in Hyphal mass (A); Mycelia (B); Spores (C) and Spores-magnified (D).
In this study, we have developed an easy, rapid and reproducible protocol for genetic transformation of F. oxysporum using GFP construct. This protocol is par with the most commonly used AMT method, and can be adopted for transformation of other filamentous fungi. In fact, we have been using this protocol for functional genomics studies in F. oxysporum and Alternaria solani using hair-pin RNAi constructs.
We are grateful to the Department of Biotechnology (Grant No. BT/R10713/ AGR/36/601/2008) and Department of Science and Technology (Grant No. SR/SO/ BB-29/2008), New Delhi for financial assistance. Neeru Singh is indebted to Council of Scientific and Industrial Research for awarding senior research fellowship. We thank K. Natarajaswamy for critically reading the manuscript.