Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2011) Volume 1, Issue 1
Keywords: Shiga toxin producing E. coli, Jejunal Haemorrhage Syndrome, Haemorrhagic Enteritis, Colonization, Prebiotic, Probiotic
Plant disease epidemics have created an ecologically unbalanced system in modern agriculture. Deterrence of such epidemics for the most part achieved through use of chemical fungicides which have greater repercussion on environment and human health. Also progressive confrontation in a midst of pathogen resistance to accessible chemical plant protectants has engrossed the need of alternative methods of disease control.
Fungi of the genus Trichoderma are important biocontrol agents of several soil borne phytopathogens. The Trichoderma species serves as a potential alternative to chemical control measure and growing pathogen resistance crop cultivars. This may be attributable to their diverse metabolic capability and aggressively competitive nature [1,2]. Trichoderma use different mechanism for control of phytopathogens which includes mycoparasitism, out compete pathogenic fungi for nutrients, secretion for antibiotics and fungal cell wall degrading enzymes [3,4]. Trichoderma sp. includes a plethora of strains which differ in their innocuousness and effectiveness as biocontrol agents. Furthermore it is difficult to predict the degree of synergism and the behavior of a biocontrol agent in a natural pathosystem. The molecular technique like Random Amplified Polymorphic DNA (RAPD) developed by Williams et al. [5] has been used for genetic and taxonomic studies for several fungi including Trichoderma sp. [6,7]. Inter Simple Sequence Repeats (ISSR) have been used as another effective method to characterize genetic variability. Since the evolutionary rate within ISSR is considerably higher than other types of DNA, the likelihood of finding polymorphism is greater compared to RAPD [8]. The Internal Transcribed Spacer (ITS) region is perhaps the most widely sequenced DNA region in fungi. It has typically been most useful for molecular systematic study at species level, and even within species [9-12]. Kindermann et al. [13] attempted a first phylogenetic analysis of the whole genus of Trichoderma using sequence analysis of the ITS1 region of rDNA. Thus the present study was to characterize the cryptic species of Trichoderma associated with biological control and to establish the relation between bioefficacy and molecular characters existed if any.
Isolation and identification of Trichoderma
Soil samples were collected from various experimental fields of Indian Agricultural Research Institute (IARI), Pusa, New Delhi. A total no. of twelve isolates of Trichoderma species was isolated and identified in specific selective medium. The identification of Trichoderma isolates were confirmed and deposited in Indian Type Culture Collection (ITCC), IARI, Pusa, New Delhi.
Confrontation assays in vitro
In vitro confrontations were studied by performing dual culture technique described by Dennis and Webster [14] was used to test the antagonistic ability of Trichoderma isolates against the phytopathogenic fungi Pythium aphanidermatum and Sclerotinia sclerotiorum. The host fungus and Trichoderma were grown on potato dextrose agar (PDA) for a week at room temperature (28±2°C). Growth parameters in all dual cultures were read after 7 days. The plates containing only the target pathogenic organisms without Trichoderma were taken as control to evaluate the percent growth inhibition.
Molecular characterization
The total genomic DNA was extracted from each isolate of Trichoderma based on Cetrimide Tetradecyl Trimethyl Ammonium Bromide (CTAB) mini extraction method of Crowhurst et al. [15] with modification.
Random Amplified Polymorphic DNA (RAPD) analysis
RAPD-PCR conditions for Trichoderma isolates in the present investigations were standardized. Forty random primers of 10 bp long each (10mer) of OPA and OPX (Operon Technologies, USA) series were used to amplify genomic DNA of the isolates. Out of these, sixteen primers which presented strong band resolution were chosen for the study viz., OPA 1, OPA 2, OPA 4, OPA 5, OPA 9, OPA 11, OPA 12, OPA 16, OPX 2, OPX 5, OPX 8, OPX 12, OPX 13, OPX 17, OPX 18 and OPX 20. Following the experiments for optimization of component concentrations, PCR amplification was carried out with 20 ηg/μl of genomic DNA, 2 mM MgCl2, 1U Taq DNA polymerase, 1X PCR buffer with 15 mM MgCl2, 0.6 μM decamer primers and 0.2 mM of dNTP mix. The volume was made up to 25 μl. PCR reactions were carried out in a Perkin Elmer Thermocycler under the conditions involving denaturation at 94°C for 2 minutes; 40 cycles of denaturation at 94°C for 1 minute, primer annealing at 35°C for 1 minute and primer extension at 72°C for 2 minutes; final extension step at 72°C for 4 minutes.
Inter Simple Sequence Repeats (ISSR) analysis
The procedure described by Bornet et al. [16] with minor modifications was used for carrying out the PCR reactions for ISSR analysis. Ten primers were tested for amplification at different annealing temperatures of genomic DNA of the isolates. Out of these, eight primers viz., 5’(AG)8AT3’, 5’(AG)8AC3’, 5’(GA)8AC3’, 5’(GA)9T3’, 5’(GA)9AC3’, 5’BDB(GA)8T3’, 5’YT(CA)7T3’ and 5’YC(TG)7T3’ consisting of anchored ISSR gave satisfactory amplification and band resolution. The PCR amplification was carried out with 25 ng of genomic DNA, 2.5 mM MgCl2, 1U Taq DNA polymerase, 1X PCR buffer without MgCl2, 1 μM ISSR primer and 0.2 mM dNTP mix. The volume was made up to 25 μl. PCR reactions were carried out in a Perkin Elmer GeneAmp 9600 thermocycler under the conditions involving denaturation at 94°C for 7 minutes; 30 cycles of denaturation at 94°C for 30 seconds, primer annealing at temperature specific to each primer for 45 seconds and primer extension at 72°C for 2 minutes; final extension step at 72°C for 7 minutes.
The PCR products of both RAPD and ISSR reactions were resolved on 1.4% agarose gel in 1X TBE buffer pre-stained with ethidium bromide (1μg/1ml) and electrophoresis was carried out at 90 volts for 1.5 hrs followed by 70 volts for 2 hrs and visualized under UV light on a UV- Transilluminator. The gel was photographed using a Gel documentation system.
Data analysis
The amplification products were scored for the presence “1” and absence “0” and missing data as “9”. The genetic associations between isolates were evaluated by calculating the Jaccard’s similarity coefficient for pair wise comparisons based on the proportion of shared bands produced by the primers. The similarity matrix was subjected to cluster analysis by unweighted pair group method for arithmetic mean (UPGMA) and a dendrogram was generated. The computations were performed using the program NTSYS – PC. version. 2.02 h [17].
The Jaccard’s similarity matrix was subjected to principal component analysis. This coordination method makes use of multidimensional solution of the observed relationships. PCA resolves complex relationships into a function of fewer and simpler factors. In this technique, the data matrix is derived from the distances (or similarities) between the operational taxonomic units. To simplify the description of these ‘clouds’ of points, the (principal) axes through the hyper-ellipsoid are calculated. The successive principal axes, representing the first major axis, the second axis etc., account for the greatest, the second greatest, etc. amount of variation.
Internal transcribed spacer region
The internal transcribed spacer (ITS) regions of the rDNA repeat from the 3’end of the 18s and the 5’end of the 28s gene were amplified using the two primers, ITS A and D which were synthesized on the basis of conserved regions of the eukaryotic rRNA gene (White et al. 1990). The PCR-amplification reactions were performed in a 50 μl mixture containing 50 mM KCl, 20 mM Tris HCl (pH 8.4), 2.0 mM MgCl2, 200 μM of each of the four deoxynucleotide triphosphates (dNTPs), 0.2 μM of each primer, 40 ηg/μl of template and 2.5 U of Taq polymerase. The cycle parameters included an initial denaturation at 94°C for 5 min, followed by 40 cycles of denaturation at 94°C for 1 min, primer annealing at 55°C for 2 min and primer extension at 72°C for 3 min and a final extension for 10 min at 72°C. Amplified products were separated on 1.2% agarose gel in TAE buffer, pre-stained with ethidium bromide (1 μg/ml) and electrophoresis was carried out at 60 volts for 3 hours in TAE buffer. One Kb ladder (MBI, Fermentas) was used as a marker. The gel was observed in a trans-illuminator over ultra violet light. The desired bands were cut from the gel with minimum quantity of gel portion using QIAGEN gel extraction kit.
Nucleotide sequencing and In silico analysis
The sequencing of the PCR product was carried out in automated Sequencer at Genome – Bio, Pune, India. Related sequences were searched using BLAST programme from the Gen-Bank database (https://www.ncbi.nlm.nih.gov/blast/) [18]. The multiple sequence alignment and pairwise alignment were made using BioEdit version 5.09 [19]. The neighborhood-joining bootstrap tree was created using CLUSTAL W 1.6 matrix by the CLUSTAL X programme ver. 1.81 [20].
Confrontation assay in vitro
The isolates of Trichoderma spp. viz., Th3, Th10, Th30, Th31 and ThAg belonging to T.harzianum; Tv2, Tv4, Tv12, Tv15, Tv32, TvChen and TvNir of T.viride were evaluated for their antagonistic ability against the phytopathogenic fungi Pythium aphanidermatum and Sclerotinia sclerotiorum based on percent inhibition mycelial growth and mean mycelial growth (in cm2) (Table 1). The isolate Th3 of T.harzianum found to have the maximum inhibiting effect on the growth of P. aphanidermatum (86.45%) and S. sclerotiorum (90%) which directly corresponded to their mycelial growth rate of 23.04 cm2 and 25 cm2 against the target fungus. Among the T.viride isolates TvChen was more aggressive with 80% inhibiting effect on P.aphanidermatum and TvNir had 85% impact on S.sclerotiorum, closely followed by Tv4 in either of the cases.
| Isolate no: | Species | Pythium aphanidermatum | Sclerotinia sclerotiorum | ||
| Percent inhibition of mycelial growth (%) of pathogen | Mean mycelial growth (cm2) of Trichoderma | Percent inhibition of mycelial growth (%) of pathogen | Mean mycelial growth (cm2) of Trichoderma | ||
| Th3 | T.harzianum | 86.4 | 23.04 | 90.2 | 25.00 |
| Th10 | T.harzianum | 83.0 | 21.16 | 88.1 | 24.01 |
| Th30 | T.harzianum | 78.3 | 18.49 | 85.0 | 23.04 |
| Th31 | T.harzianum | 80.0 | 20.25 | 74.5 | 16.81 |
| ThAg | T.harzianum | 74.5 | 16.81 | 72.7 | 14.40 |
| Tv2 | T.viride | 68.0 | 14.44 | 83.3 | 23.24 |
| Tv4 | T.viride | 75.0 | 16.81 | 80.5 | 18.49 |
| Tv12 | T.viride | 70.0 | 15.21 | 74.7 | 16.81 |
| Tv15 | T.viride | 56.5 | 12.96 | 60.9 | 12.96 |
| Tv32 | T.viride | 65.8 | 14.44 | 50.5 | 12.25 |
| TvChen | T.viride | 77.0 | 17.64 | 82.0 | 20.25 |
Table 1: Bioefficacy of Trichoderma isolates on Pythium aphanidermatum and Sclerotinia sclerotiorum.
Molecular characterization
Random amplified polymorphic DNA: The isolates of Trichoderma were subjected to RAPD analysis where 16 random primers resulted in robust and reproducible RAPD fragment patterns. The selected primers generated 112 RAPD bands and the size of the amplification products ranged from 250 bp to 900 bp. The percent polymorphism ranged from 60 to 100 with 4 primers OPA4, OPA5, OPX5 and OPX13 displayed 100% polymorphism (Plate 1). The genetic similarity between the isolates of Trichoderma was determined on the basis of Jaccard’s similarity coefficient. The mean value of the Jaccard’s similarity coefficient of the RAPD marker was 0.80. The highest genetic similarity was observed between the isolates Th31 and ThAg (80%) and the least was between the isolates Tv2 and Tv32 (30%). The dendrogram was generated using Unweighted Pair Group Method Arithmetic (UPGMA) based on NTSYS pc version 2.02i software resulted in three major clusters with isolates Tv2 and Tv32 as outliers (Figure 1). It was evident from the cluster analysis that the isolates representing to T.harzianum clustered in one group which had a similarity coefficient of 0.81 while isolates of T.viride segregated into two groups. Tv4, TvChen and TvNir formed second group with 78% similarity and Tv12 and Tv15 formed the third group which shared 82% similarity. The 3D depiction of principle components evidenced the close relatedness of T.harzianum isolates which formed into one subgroup and Tv4, TvChen and TvNir formed the other subgroup (Figure 2).
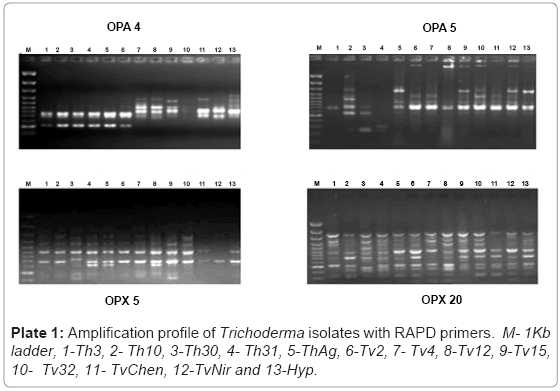
Plate 1: Amplification profile of Trichoderma isolates with RAPD primers. M- 1Kb ladder, 1-Th3, 2- Th10, 3-Th30, 4- Th31, 5-ThAg, 6-Tv2, 7- Tv4, 8-Tv12, 9-Tv15, 10- Tv32, 11- TvChen, 12-TvNir and 13-Hyp.
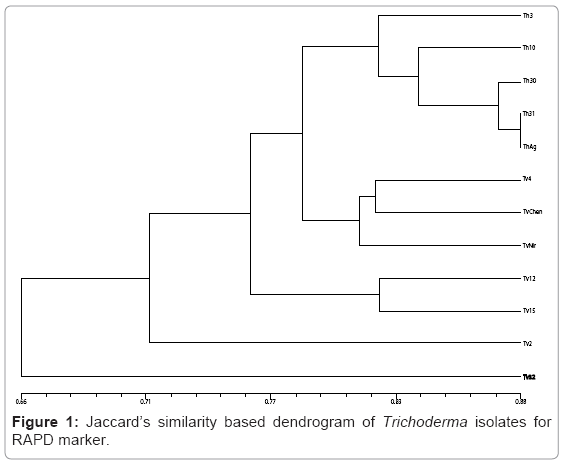
Figure 1: Jaccard’s similarity based dendrogram of Trichoderma isolates for RAPD marker.
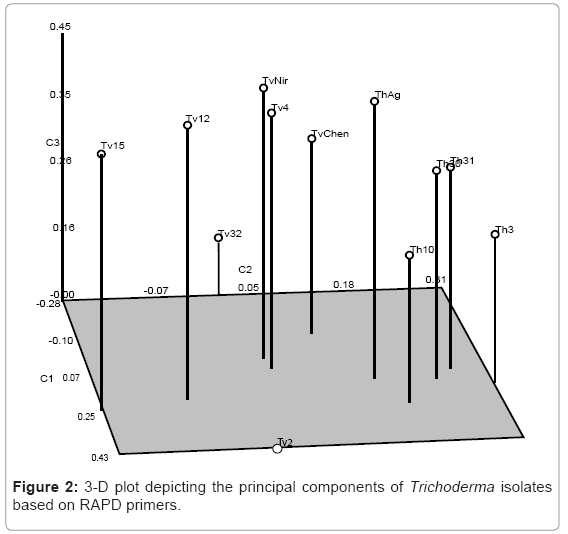
Figure 2: 3-D plot depicting the principal components of Trichoderma isolates based on RAPD primers.
Inter simple sequence repeats
Fifteen ISSR primers were screened and eight of them giving satisfactory amplification and band resolution were taken for the study. The selected primers generated 70 ISSR bands and the size of the amplification products ranged from 100 bp to 950 bp. The primer 5’ (GA) 8AC 3’ produced the maximum number of polymorphic loci (6) out of the total loci (7) followed by 5’ (GA) 9AC 3’ (6) (Plate 2). The percent polymorphism ranged from 40 to 86. The mean value of the Jaccard’s similarity coefficient of the ISSR marker was 0.76. The dendrogram based on the above data separated the isolates into two major clusters with similarities ranging from 76% to 94% (Figure 3). All the five T.harzianum isolates clustered into a single group sharing similarity around 80%. The second major cluster consisted isolates of T.viride, however these isolates formed two subgroups within the cluster. The isolates Tv2, T15 and Tv32 formed the first subgroup while Tv4, TvChen and TvNir formed the other. However Tv12 did not cluster with any of these two subgroups but stood separately with only 65% similarity with main branch. The 3D depiction of principle components reconfirmed the results obtained through dendrogram analysis (Figure 4).
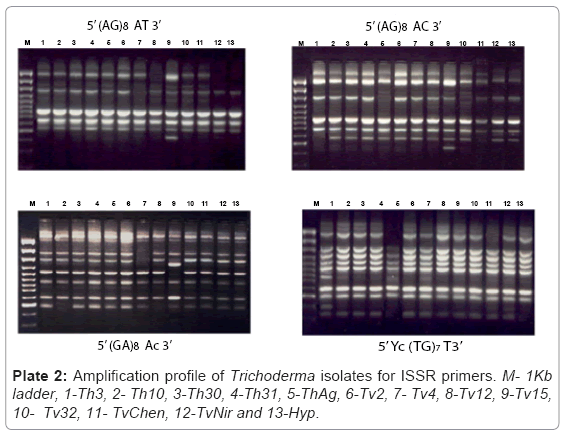
Plate 2: Amplification profile of Trichoderma isolates for ISSR primers. M- 1Kb ladder, 1-Th3, 2- Th10, 3-Th30, 4-Th31, 5-ThAg, 6-Tv2, 7- Tv4, 8-Tv12, 9-Tv15, 10- Tv32, 11- TvChen, 12-TvNir and 13-Hyp.
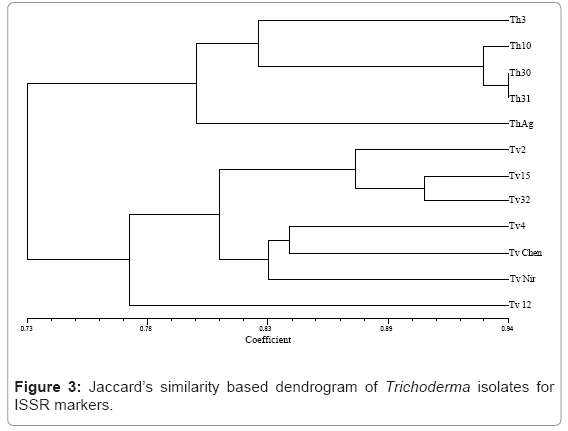
Figure 3: Jaccard’s similarity based dendrogram of Trichoderma isolates for ISSR markers.
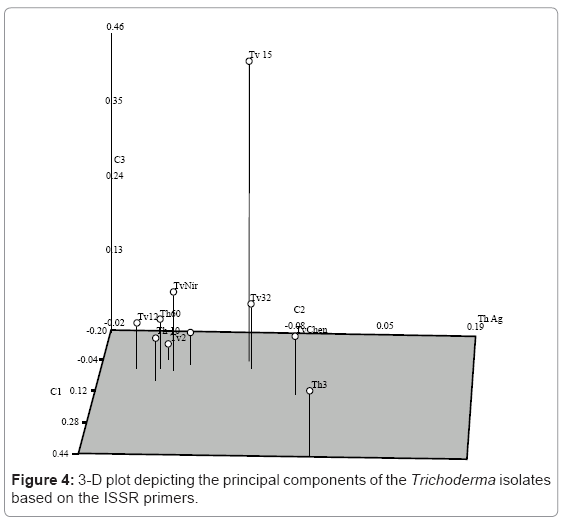
Figure 4: 3-D plot depicting the principal components of the Trichoderma isolates based on the ISSR primers.
Internal transcribed spacer
The universal primers ITS A and ITS D were used to amplify the internal transcribed spacer regions of rDNA yielding products of approximately 600bp as estimated by agarose gel electrophoresis (Plate 3). The amplified PCR products was eluded and sequenced by automated sequencer which encompass ITS 1, ITS 2 and 5.8s rDNA gene and also 18s rDNA gene having base pairs ranging 35-49 bp at the 3’ end and 20-25 bp of the 5’ end of 28s rDNA. The sequences so obtained were subjected to BLAST search for its identity and confirmation, and subsequently submitted to National Centre for Biotechnology Information (NCBI) GenBank. The accession number for sequences of ITS 1 and ITS 2 region for the twelve isolates of Trichoderma. The multiple nucleotide alignment of ITS regions was analyzed using BioEdit programme. There was substantial disparity in length of ITS sequences between T.harzianum (199 bp) and T.viride (181 bp) isolates. The isolates of T.harzianum showed 100% homology in nucleotide sequence. Conversely T.viride isolates showed nucleotide divergence of 2.2% in ITS 1 and 2.26% in ITS 2 region. The isolates Tv4, TvChen and TvNir displayed 100% homology in nucleotide sequence, however they differed from other isolates at 62nd (A instead of C) and 150th (A instead of T) nucleotide position in ITS 1 region and 27th (T instead of G) and 40th (C instead of T) in ITS 2 region. To elucidate the genetic closeness of the Trichoderma isolates a phylogenetic tree was constructed based on sequence analysis of ITS 1 and ITS 2 regions using the neighbour-joining method in treecon for windows version 1.3b on sequences aligned using ClustalW 1.7 version (Figure 5 & 6). A random sequence was used as an out-group to demonstrate the situation of the root. Bootstrap analysis of ITS 1 region with 1000 bootstrap replication demonstrated two main branches. All the isolates of T.harzianum formed one group which supported with a bootstrap value of 99.9%. The other cluster consisted of T.viride isolates which separated into two subgroups supported by bootstrap value higher than 75%. The isolates Tv4, TvChen and TvNir formed one subgroup, while Tv15, Tv2 and Tv32 formed the other subgroup. Tv12 did not group with any of the two subgroups but stood separately in this cluster. The bootstrap analysis of ITS 2 region also showed similar results, however Tv12 which stood separately in ITS 1 clustered into one of the subgroups consisting of Tv15, Tv2 and Tv32. This grouping was supported by more than 70 percent bootstrap values.
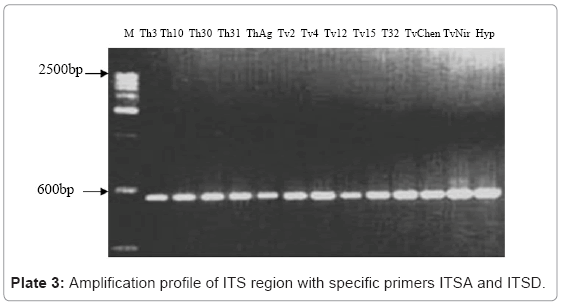
Plate 3: Amplification profile of ITS region with specific primers ITSA and ITSD.
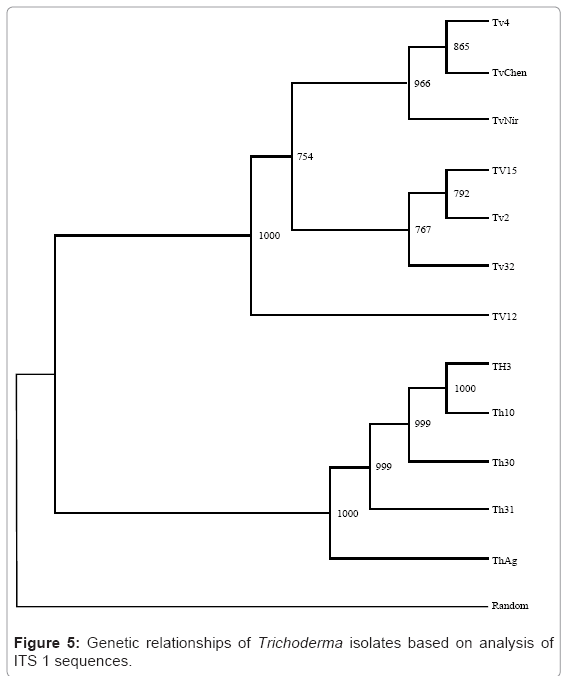
Figure 5: Genetic relationships of Trichoderma isolates based on analysis of ITS 1 sequences.
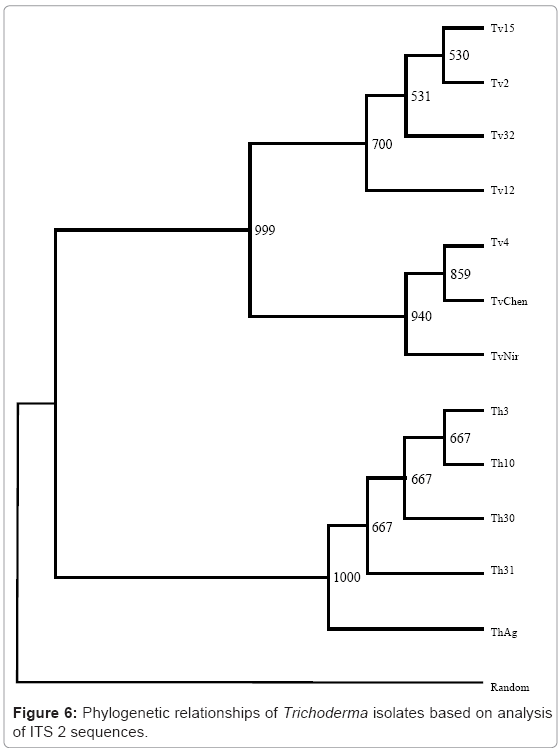
Figure 6: Phylogenetic relationships of Trichoderma isolates based on analysis of ITS 2 sequences.
In pursuit for a probability of corroborating the mycoparasitic antagonism ability of Trichoderma with respect to their molecular characters the current study was undertaken. Such corroboration will certainly be a needful aspect in elucidating the molecular mechanism involved in mycoparasitic antagonism. Dual petri-plate marriage of Trichoderma with pathogen Pythium aphanidermatum, and similarly with Sclerotinia sclerotiorum endorsed the mycoparasitic antagonism of T.harzianum and T.viride. T.harzianum isolates were more aggressive than T.viride in their antagonistic effect which ranged between 80% to 86% against P. aphanidermatum and 83% to 90% against S. sclerotiorum, on the other hand T.viride isolates ranged between 56% to 81% against P. aphanidermatum and 50% to 80% against S. sclerotiorum. However the percent inhibition of mycelial growth of Tv4, TvChen and TvNir was around 80% which was comparable with antagonistic ability of T.harzianum.
The molecular markers could not establish any variation within T.harzianum isolates instead grouped all the isolates into one cluster. The sequence identity matrix of ITS 1 and ITS 2 region showed 100% homology among these isolates of T.harzianum. None of the markers were able to identify distinct variation among T.harzianum isolates and also it is prominent that these isolates exhibited less variation in their antagonism. Therefore it can be interpreted that these isolates taken for study had no much dissimilarity among them genetically.
The RAPD markers characters divided the isolates of T.viride into two subgroups, but here Tv4, TvChen and TvNir formed a single group. Though RAPD marker formed 80% polymorphism it failed to differentiate Tv2 and Tv32. Singh et al. [21] reported that RAPD primers exhibited both inter and intra specific variation in the isolates and were able to distinguish the pathogenic isolates T.harzianum and T.virens causing green mould diseases in cultivated mushroom which was in conformity with our results. Though RAPD analysis provided good discrimination within and between species it is generally not considered a robust enough technique because of its poor reproducibility.
The ISSR technique which is more reliable reproduced similar dendrogram pattern as that of RAPD marker grouping TV4, TvChen and TvNir separately. The isolates Tv2 and Tv32 which stood as outliers in RAPD clubbed to form a subgroup, but Tv12 stood separately within the main branch. The combined ISSR and RAPD dendrogram also authenticated the above results.
The nucleotide sequence alignment of ITS 1 and ITS 2 region revealed the presence of hotspots with nucleotide substitution at four positions supported the distribution of Tv4, TvChen and TvNir into one subgroup. The dendrogram pattern obtained through ISSR was almost indistinguishable from the phylogenetic tree obtained from ITS 1 and ITS 2 sequences. This authenticates the soundness of our perception that the bioefficacy of these three isolates were manifested at the molecular level which made them to distinguish from other T.viride isolates. Conversely Goes et al. (2002) reported that there was no relationship between the polymorphism showed by the Trichoderma isolates and their hardness against R. solani based on RAPD marker. Similar kind of result was reported by Shalini et al. [22] and Shanmugam et al. [23]. However the results of molecular markers employed in this present study was complementary and confirmatory in nature which substantiates our corroboration between bioefficacy of Trichoderma isolates and molecular characters.
I thank Council of Scientific & Industrial Research, India for awarding senior research fellowship with financial support and Indian Agricultural Research Institute for its facilities provided to accomplish the research.