Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2015) Volume 3, Issue 1
Pozol is an acid fermented pre-hispanic beverage, consumed as part of the diet of ethnic groups in Southern and Southeastern Mexico. Streptococcus sp. a major component of pozol microbiota, was analyzed to assess their in vitro adherence ability to HEp-2, HeLa, HT-29 and Caco-2 cell lines. Adhesion tests were performed in 35 strains, and four of them with adherence to the different cell lines were analyzed using a Scanning Electron Microscope (SEM). Thirty-one (89%) strains could adherent to at least one of the cell lines, adherence on Caco-2 cells was the most frequently observed (63%). Diffuse and aggregative adherence phenotypes similar patterns to those described for Escherichia coli, were observed in the trial. The SEM analysis showed in one of the strains, an amorphous structure in which a large number of bacteria were included. The SEM images of other three strains, showed the presence of bacterial projections that connect them with each other and with the cells. The results showed that Streptococcus sp. strains isolated from pozol, adhere to different epithelial cell lines likely through structures which may correspond to exopolysaccharides and/or surface adhesins in bacteria. The adherence ability of these bacteria to different cultured cells could be associated with different epithelial cell colonization and the possible use of these lactic acid bacteria as probiotics if the safe use is confirmed.
Keywords: Streptococcus sp; Cell lines; SEM
“Pozol” is a pre-Hispanic, fermented drink prepared by cooking corn grain with 1% lime solution (nixtamalization). The corn grains are washed and ground to obtain a dough, which is made into balls, wrapped in banana leaves and left to ferment at room temperature during two to seven days. The fermented dough is dissolved in water and consumed as food or a refreshing drink, as much by the indigenous population as by the mixed-race population in the Southeast of Mexico (Yucatán, Quintana Roo, Campeche, Tabasco, Chiapas and Oaxaca) and Guatemala [1]. Different microorganisms including fungi, yeasts and Lactic Acid Bacteria (LAB), have been isolated from different pozol samples [2-4]. LAB are comprised of Gram-positive bacteria whose main characteristic is lactic acid production during sugar fermentation [5]. Studies relating to the characterization of microorganism components of the “pozol” biota, have shown that species of the genus Streptococcus constitute between 25 and 75% of the above-mentioned biota [2,6,7].
Analysis of the 16S subunit from the ribosomal DNA (rDNA) of Streptococcus strains isolated from “pozol” identified the existence of four different species of which S. infantarius was the predominant group [8]. LAB adapt very well to conditions in the gastrointestinal tract, in particular those microorganisms defined as probiotics that are ingested in adequate quantities and promote a positive effect on health [9,10]. One important characteristic that allows colonization of host epithelial cells is the adherence ability of microorganisms. However, the characteristics, origin of the structures and events that occur during interaction between bacteria and cells when LAB adherence is taking place are not fully defined [11-13].
Related to the factors that contribute to Gram-positive bacterial adherence is limited. Studies of some oral bacteria, such as Actinomyces naeslundii [14], S. parasanguinis [15] and S. salivarius [16-18] suggest that fimbriae may participate in the adherence process of such microorganisms.
In this study the in vitro adherence to different cultured cells lines of Streptococcus sp . strains isolated from “pozol”, was evaluated, and the cell adherence patterns of the bacteria, were also analyzed by Scanning Electron Microscope (SEM).
Thirty-five strains of Streptococcus sp . (not completely characterized) isolated from different “pozol” samples [8], were they evaluated in vitro to determine their cell adherence ability. In the assay Escherichia coli O42 (aggregative adherence), E. coli E2348/69 (localized adherence), E. coli 55784 (diffuse adherence), and E. coli HB101 (non-adherent) were used as control strains [19,20].
Adherence tests
The adherence assay described by Cravioto et al. [21] with some modifications was used. Briefly, HEp-2 (pharyngeal carcinoma), HeLa (cervical carcinoma), HT-29 (human colon adenocarcinoma) and Caco-2 (human colon carcinoma) cultured cells, were utilized in the study. Briefly, cells were grown on 72 cm2 plastic bottles in MEM (Minimum Essential Medium) (GIBCO, New York, USA) media for HEp-2 and HeLa cells; likewise, DMEM (Dulbecco’s Modified Eagle Medium) (Sigma, St. Louis, USA) to HT-29 and Caco-2 cells. In 24- well plates (Corning, New York, USA) containing 10 mm plastic lentils (Thermo Scientific, New York, USA) was added 1 ml of a cellular suspension (2.5 X 105 cells ml-1) by well and then incubated at 37°C for 24 hours in 5% CO2 and 85% humidity atmosphere. At the same time Streptococcus sp. strains were grown in MRS broth (Difco, Detroit, Mch., USA) with 1% D-Mannose (final concentration) and incubated at 35°C ± 2°C for 18 hours. The wells containing cells where washed and then added 900μL of MEM or DMEM media without serum or antibiotics together with 1% D-Mannose. The bacterial cultures were centrifuged (2000 g) for 15 min and the pellet suspended in 1 ml of MEM or DMEM media without serum or antibiotics and 1% D-Mannose. The bacterial suspension (1.0 X 108 cfu ml-1), in a 100 μL volume was added to each plate well and incubated at 37°C for 3 hours in a 5% CO2 atmosphere. Then the liquid was eliminated from each well and washed three times (PBS 1X), fixed with methanol (1 ml) for 1 min and stained with 1 ml of Giemsa (Química Meyer, México, D.F) during 10 min. Afterwards, Giemsa stain was removed and the wells washed three times with deionized water. A treatment with acetone, acetone-xilol 50/50, xilol was used to dehydrate the cells, and then each plastic lentil with the cells was resin fixed (Fisher Chemicals New Jersey, USA) on a glass slide. The slides were they observed under a light microscope (100X). An adherence test was it considered positive if at least 25% of 400 cells showed 10 or more adhered bacteria. The adherence tests were they carried out in two technical duplicates and by triplicate.
Scanning Electron Microscopy (SEM)
An ultrastructural analysis of the adherence assays was carried out using SEM according to previously reported methodology [22]. The preparations were obtained from an adherence assay in Caco-2 cells (as previously stated), of Streptococcus sp. strains 25245, A12203, 15124, and 25137 (adherent to the four cell lines used); while, the strain 25109 was used as a non-adherent control. The preparations were treated with 5% glutaraldehyde (Sigma, St. Louis, USA) for 48 hours and washed with a phosphate buffer solution (0.1M, pH7.3) continued until the glutaraldehyde remaining residues were removed. Later, with diluted osmium tetraoxide (2%) in phosphate buffer (0.1M, pH 7.3) the samples were they fixed. In addition, the preparations were dehydrated using from 30% to absolute alcohol with 10% increments. The samples were dehydrated to critical point, placing them on amyl acetate (10°C) and adding liquid CO2 for two times. Finally, the samples were placed on supports fixing them with colloidal silver before putting them in a gold bath. The preparations were observed under SEM (JEOL JSM-5900LV, North Billerica, MA) with an acceleration voltage of 13KV and Secondary Electron Imaging (SEI).
Adherence to cultured cells
Of the 35 Streptococcus sp . strains analyzed, 89% (31/39) showed adherence and of these 13% (4/31) adhered to all four-cell lines, 19% (6/31) to three, 13% (4/31) to two, and 55% (17/31) only to one cell line (Table 1). Analysis of each of the cell lines showed that 13 (37%) strains adhered to HEp-2 cells, 10 (29%) to HeLa cells, 14 (40%) to HT-29 cells, and 22 (63%) to Caco-2 cells (Table 2). Qualitative analysis by light Microscopy at the different preparations showed phenotypes like to diffuse (LDA), and aggregative (LAA) adherences (Table 1); which, were similar to those described to Escherichia coli [19]. In HEp-2 cells, in nine of the isolates the identified patterns were similar to diffuse adherence (LDA) and aggregative adherence (LAA) in four strains. For HeLa cells, seven strains showed LDA and three LAA patterns. In Caco-2 cells, 12 isolates showed LDA patterns (Figure 1a) and 10 LAA patterns (Figure 1b). In the HT-29 cells (Figure 1c), the 15414 strain displayed an adherence pattern that is similar to localized adherence (LLA) reported previously in E. coli diarrheogenic strains [19].
| Cell Cultured Lines | ||||||
|---|---|---|---|---|---|---|
| Hep-2 | HeLa-3 | HT-29 | Caco-2 | |||
| Strains | Adherence Pattern | |||||
| 15133 | - | LDA | LDA | LAA | ||
| 15430 | - | - | - | LAA | ||
| 15220 | - | LDA | - | - | ||
| 25245 | LDA | LDA | LDA | LAA | ||
| 15124 | LAA | LAA | LAA | LAA | ||
| 25113 | - | - | - | LAA | ||
| 25148 | LDA | LDA | - | LAA | ||
| 25139 | - | - | LDA | LDA | ||
| 25233 | LDA | - | LDA | LAA | ||
| 25421 | LDA | - | LDA | LDA | ||
| 25137 | LAA | LAA | LDA | LAA | ||
| 15125 | - | - | - | LAA | ||
| 15414 | - | LDA | LLA | LDA | ||
| 25124 | - | LDA | - | LDA | ||
| A56203 | LDA | - | - | - | ||
| A57103 | LDA | - | LAA | LDA | ||
| A57206 | - | - | LAA | LDA | ||
| A45208 | LAA | - | - | - | ||
| A37103 | - | - | - | LDA | ||
| A37202 | - | - | LDA | - | ||
| A36111 | LDA | - | - | - | ||
| A12203 | LAA | LAA | LAA | LDA | ||
| A56101 | - | - | LAA | - | ||
| A56201 | - | - | - | LAA | ||
| 15319 | - | LDA | - | - | ||
| A46112 | - | - | LDA | - | ||
| A46113 | - | - | - | LAA | ||
| A47212 | - | - | - | LDA | ||
| A56208 | LDA | - | - | - | ||
| A45201 | LDA | - | - | LAA | ||
| A45226 | - | - | - | LDA | ||
Table 1: Adherence on different cultured cells of Streptococcus sp. strains isolated from pozol samples, LDA=Like Diffuse Adherence, LAA=Like Aggregative Adherence, LLA=Like Localized Adherence, - =Non Adherent.
| Adherence Phenotypes (%) | ||||
|---|---|---|---|---|
| Cell Culture | LDA | LAA | LLA | Total (%) |
| Hep-2 | 9 | 4 | 0 | 13 (37) |
| HeLa | 7 | 3 | 0 | 10 (29) |
| HT-29 | 8 | 5 | 1 | 14 (40) |
| Caco-2 | 10 | 12 | 0 | 22 (63) |
| Non-adherent | - | - | - | 4(11) |
Table 2: Adherence phenotypes of Streptococcus sp. on different cultured cell lines. LDA=Like Diffuse Adherence, LAA=Like Aggregative Adherence, LLA=Like Localized Adherence.
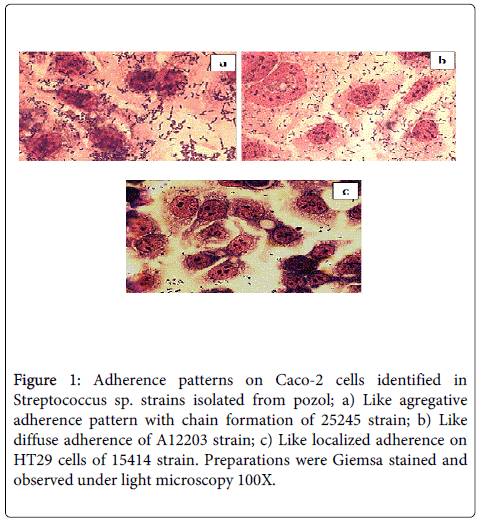
Figure 1: Adherence patterns on Caco-2 cells identified in Streptococcus sp. strains isolated from pozol; a) Like agregative adherence pattern with chain formation of 25245 strain; b) Like diffuse adherence of A12203 strain; c) Like localized adherence on HT29 cells of 15414 strain. Preparations were Giemsa stained and observed under light microscopy 100X.
Scanning Electron Microscopy (SEM)
strains 25137, 25245, A12203, and 15124 adherent to the four cell lines (Table 1), showed elongated fiber pilus-like interconnecting to bacteria; likewise, some cells of strain 25137 (LAA) forming microcolonies appeared deeply embedded in the brush border to mucosal cells (Figure 2). In addition, elongated fiber that connect bacteria to the cell (Figure 2), were observed in strains A12203 (LDA) and 25245 (LAA). With regard to strain 15124 (LAA), extended structures were observed that connect bacteria to each other and other smaller ones that link these bacteria to the cell. In the non-adherent strain 25109, no structures or prolongations it were identified (Figure 2).
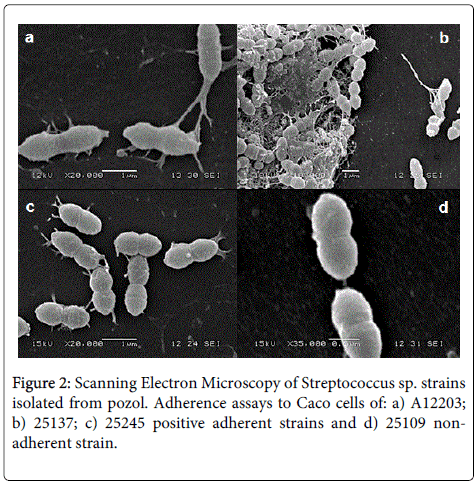
Figure 2: Scanning Electron Microscopy of Streptococcus sp. strains isolated from pozol. Adherence assays to Caco cells of: a) A12203; b) 25137; c) 25245 positive adherent strains and d) 25109 nonadherent strain.
Adherence to intestinal mucosa is a prerequisite of probiotic microorganisms to colonize the epithelium and compete with enteropathogenic bacteria found in such spaces [23,24]. Adherence of lactic bacteria in animal models is complicated and on occasion, difficult to evaluate. Bearing this in mind, some different in vitro systems have been adopted [25-27] and of these cultured intestinal epithelial cells are the more common [24,28-30]. In the current study, 89% of the Streptococcus sp. strains studied were adherent to different cell lines but with a higher frequency on intestinal HT-29 (40%) and Caco-2 (63%) cell lines. These results are similar to those reported by Sung-Mee and Dung-Soon [31] and Lee et al. [32], when they examined the adherence properties of LAB to Caco-2 and HT-29 cells and Lactobacillus strains to HT-29 cells. They observed that 28 (49%) of their samples adhered with a superior adhesion ability to that observed in L. rhamnosus GG [33]. These results and the observed in this current study, show that in general, LAB showed structures that contribute to the adherence of bacteria to intestinal cells.
Results of adherence test to HEp-2 (37%) and HeLa (29%) cells, demonstrate that Streptococcus sp. strains isolated from pozol, are able to colonize both the intestinal epithelium and others as the pharyngeal and cervical epithelia. The fact that the analyzed bacteria showed adherence in the presence of D-Manosse indicates that the elements related to adherence are resistant to manosse, probably in a similar way to that described for Gram-negative bacteria [19,21,34]. The adherence tests also revealed the presence of diffuse, aggregative and localized phenotypes, similar to those described for E. coli [19,34,35]. These same adherence phenotypes, have been identified in lactic acid bacteria strains isolated from cooked meat products [36], and in strains of Lactobacillus paracasei isolated from a fermented drink (honey water) obtained from an agave plant (Hernández- Ramírez – personal communication).
The strains described as 15124, 25139, 25421 and 25142 displayed adherence of the LDA (Like Difuse) and LAA (Like Aggregative) phenotypes in the four epithelial cell lines; however, some other (15133, 25245, 25148, 25233, 25137, 15414, A57103, A57206, A12203, and A45201) strains, showed different adherence patterns. Regarding adhesion of bacteria to intestinal, epithelial cells, passive electrostatic, and hydrophobic type forces, as well as covalent types of interactions induced by specific adhesins and their respective receptors may are playing a role [37]. In relation to adherence of Lactobacillus as probiotic, has not yet been well defined the adherence factors and therefore has been suggested that proteins, glycoproteins, teichoic or lipoteichoic acids mediate the interaction bacteria-cells [38-40]. With this in mind, it has been proposed that some components of the cell wall induce different types of bacteria-cell interactions, explaining the presence of different adherence phenotypes [41-43]. Lévesque, et al. [17] reported in a strain of S. salivarius , the presence of genes that code for a protein related to bacterial adherence. On the other hand, Sara and Uwe [44], Logan, et al. [45], Ossowski, et al. [33] and Sanchez, et al. [46] in different BAL strains, also have described the participation of bacteria surface proteins in promoting adhesion to intestinal tissues.
SEM conducted as part of this current study, revealed that strain 25137 forms a matrix that consists probably of exopolysaccharides with bacteria aggregates (Figure 2b). Maldonado, et al. [47] reported that LAB strains secrete polysaccharides, involved with the adhesion to intestinal mucosa, the formation of bacterial agglomerates, and prolongations that contribute to the cells adhesion. However, is possible the participation of different structures in the bacteria cell adherence, this because the same procedure with strains 25245 (LAA), A12203 (LDA) and 15124 (LAA), showed the presence of filament- type structures that connect bacteria to each other and to epithelial cells (Figures 2a and 2c). The results of this study indicate that even between bacteria of the same genus and species, adherence could be related to different structures and events making the characterization of these adhesins important to determine.
The biochemical characterization of Streptococcus sp . of the strains of our study show that them are members of Streptococcus bovis/Streptococcus equinus complex (SBSEC) species, however, molecular studies are necessaries to define the species. Overall SBSEC group members are considered as putative pathogens, however, PCR and Southern blotting analyses of S. macedonicus ACA-DC 198, indicated the absence of several Streptococcus pyogenes pathogenicity genes [48]. PCR test conducted in our strains to identify emm (M protein), SpeA/SpeC (erythrogenic toxin), sic (complement inhibitory protein) and of (opacity factor), showed negative results in all strains.
Although it requires more studies, one could propose that our strains can be considered as potential probiotic bacteria.
The authors would like to thank the General Office of Academic Personnel Affairs – UNAM (DGAPA) and the project “Uso Potencial como Probióticos de Bacterias Lácticas Aisladas del Pozol Bebida Fermentada de Uso Tradicional en México” (PAPIIT-IN219611-3) for their support.
In addition, N.L. Ramírez-Chavarin and C.A. Eslava-Campos are grateful for the support of the Program for Post-Doctoral Grants at the Faculty of Medicine, UNAM-DGAPA and UNAM-CONACYT.