Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2016) Volume 5, Issue 2
Epinephrine is an important neuroendocrine regulator to control growth hormone (GH) secretion in vertebrates. Somatolactin (SL), the latest member of the GH family, is a novel pituitary hormone with diverse functions in fish. In a previous report it was shown that epinephrine had a potent inhibitory effect on SL release in fish. However, very little is known about the mechanisms responsible for epinephrine inhibition of SL gene expression. In primary cultures of tilapia neurointermediate lobe (NIL) cells, epinephrine not only reduced SL mRNA levels, but could also abolish pituitary adenylate cyclase-activating polypeptide (PACAP)-stimulated SL gene expression. The inhibitory effects of epinephrine on SL gene expression were mimicked by additions of α2-adrenergic agonists clonidine and UK14304, whereas similar treatments with the α1-agonist cirazoline or the β-agonist isoproterenol had no effects in this regard. In parallel experiments, the SL response to epinephrine was significantly abolished by co-incubations with the α2- antagonist yohimbine, but the α1- or β-antagonist was not effective in this regard. In tilapia NIL cells, the α2-adrenergic agonist clonidine suppressed cAMP production and blocked forskolin and PACAP induction of total cAMP production. By using a pharmacological approach, the adenylate cyclase (AC) activator- and cAMP analog-stimulated SL responses were blocked treatment with clonidine. Furthermore, adrenergic inhibition of SL gene expression was also mimicked by inhibiting AC and blocking protein kinase A (PKA). These results, as a whole, suggest that α2-adrenergic stimulation can downregulate SL gene transcription by inhibiting the AC/cAMP-dependent mechanism at the tilapia pituitary level.
<Keywords: Epinephrine; Somatolactin; Tilapia; Pituitary cells; Signaling pathways
EP: Epinephrine; SL: Somatolactin; GH: Growth Hormone; PACAP: Pituitary Adenylate Cyclase-activating Polypeptide; NIL: Neurointermediate Lobe; cAMP: Cyclic Adenosine Monophosphate; AC: Adenylate Cyclase; PKA: Protein Kinase A
In vertebrates, the adrenergic system is known to be involved in the regulation of growth hormone (GH) secretion and gene expression at the hypothalamic and pituitary levels. The adrenergic receptors (or adrenoceptors) mediating central and peripheral actions of epinephrine are typically subdivided into three main families (α1, α2, and β) based on their pharmacology, structure, and signaling mechanisms [1,2]. The α1-adrenoceptors increase levels of intracellular calcium, whereas α2- and β-adrenoceptors inhibit and stimulate adenylyl cyclase (AC), respectively [1]. In mammals, the influences of epinephrine on GH secretion are selective for one type of adrenoceptors at the hypothalamic or pituitary cells levels. Results of studies in both primates and rodents have shown that α2-adrenoceptors play an important role in stimulating GH secretion in the central nervous system [3]. In contrast, α2-adrenoceptors are not involved in GH secretion [4] and GH gene expression [5] at the pituitary cell level. Although still controversial, β-adrenergic activation is also involved in regulation of GH secretion. In the baboon, central action of norepinephrine was mediated by β-adrenergic receptors to inhibit GH release [6], whereas β-adrenergic receptor agonists can stimulate rat GH secretion in rat pituitary cells [7] and increase GH gene expression in ovine pituitary cells [5]. Similar to mammals, the role of adrenergic as a GH-releasing factor has also been confirmed in lower vertebrates including fish. In goldfish, intraperitoneal injection of norepinephrine resulted in decreased serum GH levels, whereas intraventricular injection of norepinephrine has no effects on GH secretion in vivo [8]. Since the pituitary lies outside of the blood-brain barrier, these results suggest that norepinephrine may directly act on the pituitary to inhibit GH secretion. This idea is further corroborated by in vitro studies showing that norepinephrine is effective in suppressing goldfish GH secretion [9] and grass carp GH gene [10] in pituitary cells via α2-adrenoceptors.
Somatolactin (SL), the latest member of GH family, is a pituitary hormone unique to fish species. SL is derived from GH during early stages of gnathostome evolution [11], but lost secondarily in the lineage leading to land vertebrates after the lungfish branched off [12]. SL appears to have significant roles in chromatophore regulation and lipid metabolism [13,14], maturation [15], stress response [16], ion transport [17] and acid-base balance [18]. Among the known SL inhibitors in fish, somatostatin can inhibit SL gene expression through coupling of AC/cAMP and PLC/IP3/PKC cascades [19]. In addition to somatostatin, epinephrine has been implicated as a SL-releasing inhibitor. A previous in vitro study had demonstrated that epinephrine dose-dependently inhibited SL release in the organ-cultured pituitary of rainbow trout [20]. However, no information is available regarding the mechanisms responsible for epinephrine inhibition of SL gene expression at the pituitary level.
In the present study, using primary cultures of tilapia neurointermediate lobe (NIL) cells as a model, the effects of epinephrine on the SL mRNA expression were investigated at the pituitary level. The receptor specificity was further characterized using adrenergic analogs for α1-, α2-, and β-adrenoceptors. The post-receptor signaling mechanisms for SL gene expression were also investigated. In this study, we have demonstrated for the first time that epinephrine can act at the pituitary level via α2-adrenoceptors to regulate SL gene expression through the AC/cAMP-dependent mechanism.
Animals
Sexually mature male tilapia (Oreochromis mossambicus) (standard length: 11 ± 0.5 cm, body weight: 50 ± 5.0 g) were maintained in freshwater aquaria at 28ºC under 10 hr dark/14 hr light photoperiod. The fish were fed commercial diet (40% protein, 12% fat, 2% fiber, 8.5% moisture, 8% ash, Tongwei, China) to satiety twice a day at 10:00 and 16:00. During the process of tissue sampling, the fish were sacrificed by spinosectomy after anesthesia with 0.05% MS222 (Sigma, St Louis, MO) according to the procedures approved by the Animal Ethics Committee of Sichuan University.
Pharmacological agents
Epinephrine and Ovine PACAP38 were obtained from Sigma (St. Louis, Mo). Adrenergic analogs including clonidine, UK 14304, cirazoline, isoproterenol, propranolol, 2-{[β-(4-Hydroxyphenyl) ethyl] aminomethyl}-1-tetralone hydrochloride (HEAT), and yohimbine were obtained from Tocris (Bristol, UK). These pharmacological compounds have been previously used in goldfish [9], grass carp [10], tilapia [21] and zebrafish [22], confirming that they are highly selective for respective targets in fish models. 3-isobutyl-1-methylxanthine (IBMX), 8-(4-chloro-phenylthio)-cAMP (CPT-cAMP), forskolin, H89, MDL12330A and actinomycin D were obtained from Calbiochem (San Diego, CA). Stock solution of PACAP was dissolved in double-distilled deionized water and stored frozen in small aliquots at -80ºC. Epinephrine and clonidine were dissolved freshly in culture medium right before drug treatment to avoid oxidation caused by prolonged storage. Other test agents were first dissolved in dimethyl sulfoxide (DMSO). Stock solutions of test substances were diluted with prewarmed (28ºC) culture medium to appropriate concentrations 15 min prior to drug treatment. The final dilutions of DMSO were less than 0.1% and had no effects on SL gene expression in tilapia NIL cells.
Primary culture of tilapia NIL cells
Tilapia pituitary cells were prepared by trypsin/DNase II digestion method as described previously [23]. Briefly, The NIL of individual pituitaries was isolated by manual dissection under a stereomicroscope, diced into 0.6 mm fragments using a McILwain tissue chopper (Brinkmann, Mississauga, Ont.), and digested in type II trypsin (4 mg/ ml, GIBCO) for 30 min at 28ºC with constant shaking. After that, the reaction was terminated by adding trypsin inhibitor (2.5 mg/ml, Sigma) and pituitary fragments were dispersed in Ca2+- free MEM [S-MEM with 26 mM NaHCO3, 25 mM HEPES, 1% antibiotic-antimycotic, and 0.1% BSA; pH 7.7] with DNase II (0.01 mg/ml, Sigma). After that, total cell yield and percentage viability were estimated by cell counting in the presence of trypan blue using a hemocytometer. NIL cells were cultured in 48-well culture plates (Costar, Corning Inc., N.Y.) at a seeding density of ~1 × 106 cells/well in M199 (Invitrogen). NIL cells were incubated overnight at 28ºC under 5% CO2 and saturated humidity to allow for the recovery of membrane receptors after trypsin digestion. On the following day, culture medium was replaced with serum-free M199 and drug treatment was initiated for the duration as indicated in individual experiments.
Real-time PCR measurement of SL
Tilapia NIL cells were seeded at a density of ~1 × 106 cells/ml/well in 48-well culture plates and treated with hormones or drugs for the duration as indicated in individual experiments. After that, total RNA was isolated using RNAzol (MRC, Cincinnati, OH, USA), digested with RNase-free DNase I to remove genomic DNA contamination, and reversely transcribed using M-MLV (TaKaRa, Dalian, China). After that, real-time PCR assays were performed on the CFX96 Real- Time PCR Detection System (Bio-Rad Laboratories, CA, USA). PCR reaction were conducted with a SYBR Select Master Mix kit (Applied Biosystems) using the primers specific for tilapia SL (GenBank No: AB442015) [SL forward primer: 5' CCCACTCCCTTTGCGACTT 3' and SL reverse primer: 5' TAGCGGTCCAGTGTCGTCT 3']. Real-time PCR for the SL were performed with initial denaturation at 94°C for 3 min followed by 35 cycles of amplification with denaturation at 94°C for 30 sec, annealing at 56°C for 30 sec, and extension at 72°C for 30 sec and then fluorescent signal collection at 80ºC for 1 sec. For data calibration, serial dilutions of plasmid DNA containing the ORF of SL was used as the standards for these real-time PCR assay. As an internal control, real-time PCR for 18S rRNA was conducted using the primers specific for 18S rRNA [forward primer: 5' GGACACGGAAAGGATTGACAG 3' and reverse primer: 5' GTTCGTTATCGGAATTAACCAGAC 3']. In these experiments, no significant changes were observed for 18S rRNA expression. The quantitative results were normalized as a ratio of the target gene/18S rRNA expression level.
Measurement of cAMP production
The NIL cells were seeded at a density of ~1 × 106 cells/2 ml/dish in poly-D-lysine precoated 35-mm dishes and cultured overnight at 28ºC as previously described [19]. On the following day, culture medium was replaced with 0.9 ml HEPES-buffered Hanks’ balanced salt solution with 0.1% BSA and 0.1 mM IBMX. IBMX, the inhibitor for phospho-diesterases, was included to prevent cAMP degradation in NIL cells. Drug treatment was initiated with various combinations of drugs at appropriate concentrations and the cells were allowed to incubate at 28ºC for 30 min. After that, culture medium was harvested for the measurement of cAMP release whereas cellular cAMP content was extracted from NIL cells with 1 ml PBS. These cAMP samples were quantified by using a cAMP ELISA kit (Wuhan EIAab Science Co., Ltd).
Data transformation and statistics
All of the experiments were performed at least twice, and all of the treatments in each experiment were tested in quadruplicate from 40 individual fish. For real-time PCR of SL transcripts, standard curves with a dynamic range of 105 and correlation coefficient of ≥ 0.95 were used for data calibration with Bio-rad CFX 3.0 software. The quantitative results were normalized as a ratio of the target gene/18S rRNA expression level. Data of SL gene expression were transformed as a percentage of the mean value in the control group without drug treatment (referred to as “% Ctrl”). Data presented (as means ± SEM, N = 4) were analyzed using ANOVA followed by Duncan’s test using Prism 6.0. Differences between groups were considered as significant at P < 0.05.
Adrenergic regulation of SL gene expression in tilapia NIL Cells
To examine adrenergic regulation of SL gene expression at the pituitary level, we used a tilapia NIL cells as a model to test the direct effects of epinephrine on SL mRNA expression. As shown in Figure 1A, treatment with epinephrine (1 μM) could time-dependently inhibit SL mRNA expression, and the maximal inhibition on SL mRNA expression was observed at 48 h. The duration of drug treatment was fixed at 48 h unless stated otherwise in subsequent experiments. In parallel experiments, increasing concentrations (0.01-10 μM) of epinephrine were effective in triggering a dose-dependent decrease in SL mRNA levels (Figure 1B). The minimal effective dose for epinephrine to inhibit SL gene expression was 10 nM, and SL transcripts were reduced to 45% of control levels at maximal effective doses of 1 μM.
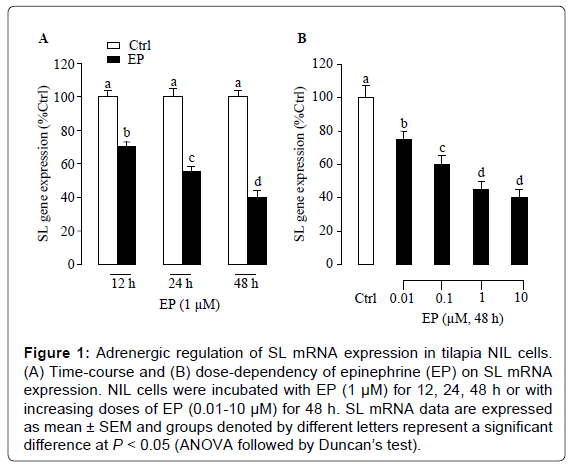
Figure 1: Adrenergic regulation of SL mRNA expression in tilapia NIL cells. (A) Time-course and (B) dose-dependency of epinephrine (EP) on SL mRNA expression. NIL cells were incubated with EP (1 μM) for 12, 24, 48 h or with increasing doses of EP (0.01-10 μM) for 48 h. SL mRNA data are expressed as mean ± SEM and groups denoted by different letters represent a significant difference at P < 0.05 (ANOVA followed by Duncan’s test).
Effects of adrenergic agonists on SL gene expression
To clarify the receptor specificity for epinephrine action, the effects of adrenergic agonists on SL gene expression were examined in tilapia NIL cells. As shown in Figure 2A, the inhibitory effects of epinephrine on SL gene expression were mimicked by the increasing levels of α2- agonist clonidine (0.01-10 μM) and UK14304 (0.01-10 μM), whereas α1-agonist cirazoline (0.01-10 μM) or β-agonist isoproterenol (0.01-10 μM) was not effective in altering SL mRNA expression (Figure 2B).
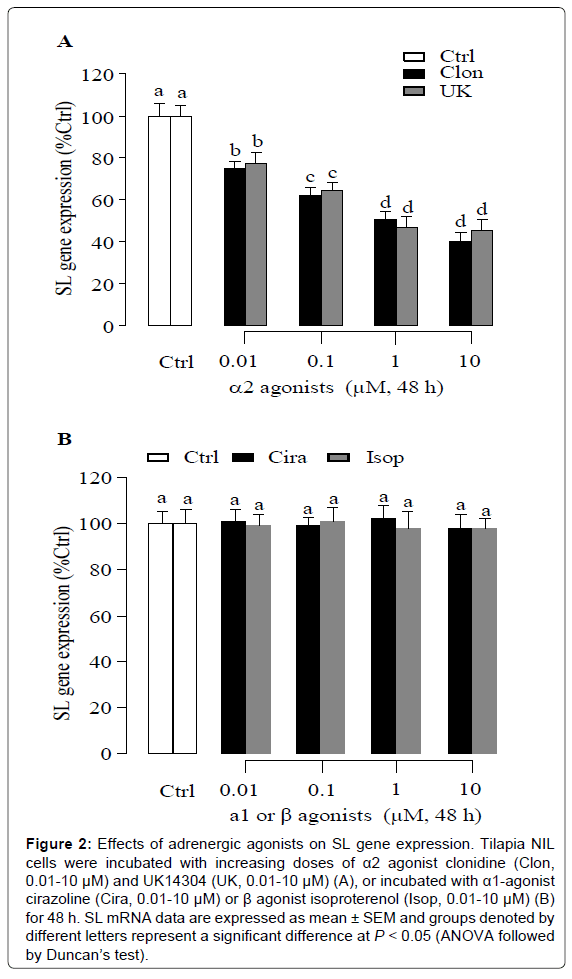
Figure 2: Effects of adrenergic agonists on SL gene expression. Tilapia NIL cells were incubated with increasing doses of α2 agonist clonidine (Clon, 0.01-10 μM) and UK14304 (UK, 0.01-10 μM) (A), or incubated with α1-agonist cirazoline (Cira, 0.01-10 μM) or β agonist isoproterenol (Isop, 0.01-10 μM) (B) for 48 h. SL mRNA data are expressed as mean ± SEM and groups denoted by different letters represent a significant difference at P < 0.05 (ANOVA followed by Duncan’s test).
Effects of adrenergic antagonists on epinephrine inhibition of SL gene expression
To further characterize the receptor specificity of epinephrine inhibition on SL gene, tilapia NIL cells were exposed to adrenergic antagonists specific for different subtypes of adrenoceptors. In these experiments, the inhibitory effect of epinephrine was tested in the presence or absence the α2-antagonist yohimbine, or α1-antagonist HEAT, or β-antagonist propranolol. In this case, epinephrine consistently suppressed SL gene expression and this inhibitory action was blocked by simultaneous treatment with α2-antagonist yohimbine (5 μM, Figure 3A), but not affected by treatment with the α1-antagonist HEAT (Figure 3B) and β-antagonist propranolol (Figure 3C). To further examine the specificity of α2 receptor activation on SL gene expression, the effects of α2-antagonist yohimbine on the SL response to the α2-agonists clonidine and UK14304 were studied in tilapia NIL cells. In the present study, α2-agonist clonidine- and UK14304- inhibited SL gene expression were prevented in the presence of the α2- antagonist yohimbine (Figures 3D and 3E).
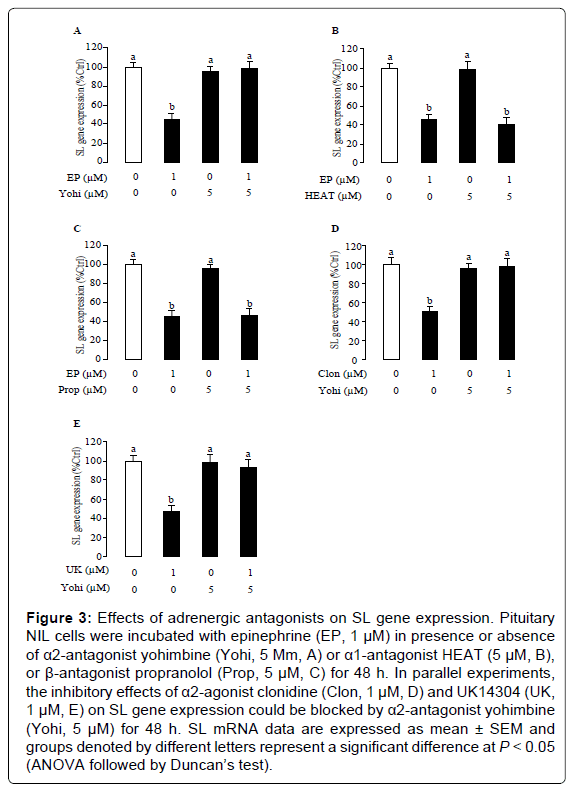
Figure 3: Effects of adrenergic antagonists on SL gene expression. Pituitary NIL cells were incubated with epinephrine (EP, 1 μM) in presence or absence of α2-antagonist yohimbine (Yohi, 5 Μm, A) or α1-antagonist HEAT (5 μM, B), or β-antagonist propranolol (Prop, 5 μM, C) for 48 h. In parallel experiments, the inhibitory effects of α2-agonist clonidine (Clon, 1 μM, D) and UK14304 (UK, 1 μM, E) on SL gene expression could be blocked by α2-antagonist yohimbine (Yohi, 5 μM) for 48 h. SL mRNA data are expressed as mean ± SEM and groups denoted by different letters represent a significant difference at P < 0.05 (ANOVA followed by Duncan’s test).
α2-adrenergic regulation of SL transcript stability
To shed light on the mechanisms for α2-adrenergic inhibition of SL mRNA expression, the effects of α2-agonist clonidine treatment on SL mRNA stability were also tested. Clearance analysis of SL transcript was performed in NIL cells pretreated with the transcription inhibitor actinomycin D (8 μM). In this case, the clearance rate of SL transcripts expressed as the time required for half of the original amount of SL mRNA to degrade (i.e., T1/2) was used as an index to monitor SL transcript stability. As shown in Figure 4, SL mRNA levels were reduced gradually in a time-dependent manner with a T1/2 value of ~ 31 h. However, the clearance profile or T1/2 value for SL transcripts was not affected by the co-treatment with clonidine (1 μM).
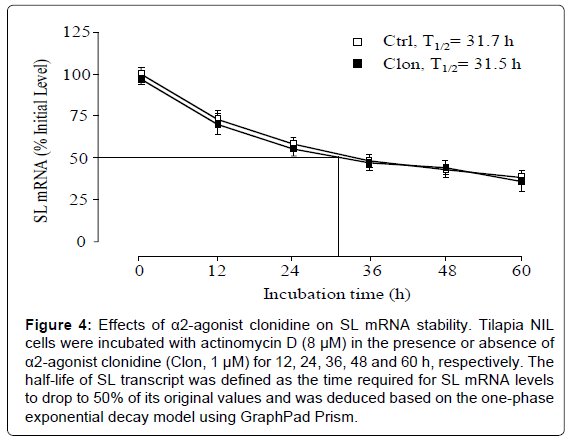
Figure 4: Effects of α2-agonist clonidine on SL mRNA stability. Tilapia NIL cells were incubated with actinomycin D (8 μM) in the presence or absence of α2-agonist clonidine (Clon, 1 μM) for 12, 24, 36, 48 and 60 h, respectively. The half-life of SL transcript was defined as the time required for SL mRNA levels to drop to 50% of its original values and was deduced based on the one-phase exponential decay model using GraphPad Prism.
Adrenergic regulation of basal and PACAP-stimulated SL mRNA expression
The role of PACAP as a novel SL regulator has received increasing attention in teleosts [24-28]. To test the functional interactions between PACAP and epinephrine in regulating SL gene expression, tilapia NIL cells were challenged with PACAP (10 nM) for 48 h in the presence or absence of epinephrine. In this study, basal levels of SL mRNA were elevated by PACAP treatment, and this stimulatory action could be blocked by simultaneous incubation with epinephrine (1 μM, Figure 5). Similarly, this stimulatory effect was also blocked by simultaneous treatment with the α2-agonist clonidine (1 μM).
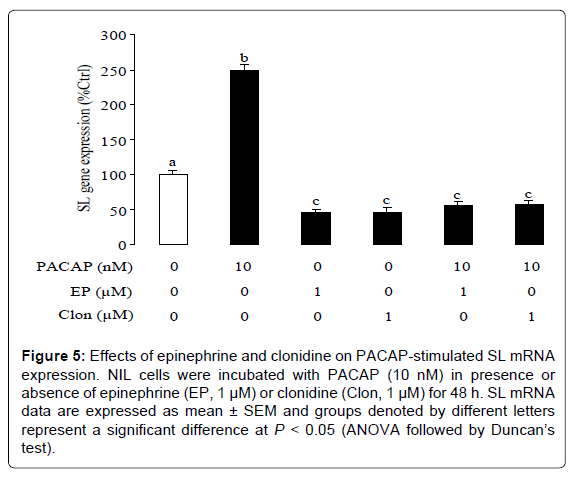
Figure 5: Effects of epinephrine and clonidine on PACAP-stimulated SL mRNA expression. NIL cells were incubated with PACAP (10 nM) in presence or absence of epinephrine (EP, 1 μM) or clonidine (Clon, 1 μM) for 48 h. SL mRNA data are expressed as mean ± SEM and groups denoted by different letters represent a significant difference at P < 0.05 (ANOVA followed by Duncan’s test).
α2-adrenergic inhibition of cAMP production
Given that is known to inhibit cAMP synthesis via activation of α2- adrenergic receptor at the pituitary level in fish [10], the effects of α2- adrenergic receptor agonist clonidine on cAMP production were tested in tilapia NIL cells. As shown in Figure 6A, forskolin was effective in elevating total cAMP production. In contrast, basal levels and forskolininduced increases in cAMP production were significantly attenuated by application of clonidine (Figure 6A). In tilapia NIL cells, cAMP contents were also significantly elevated by PACAP treatment (10 nM), and this stimulatory action could be alleviated by simultaneous incubation with clonidine (Figure 6A). In parallel studies, increasing levels (0.01-10 μM) of the AC inhibitor MDL12330A (Figure 6B) and the PKA inhibitor H89 (Figure 6C) could mimic the dose dependence of clonidine inhibition on SL gene expression. To further evaluate the functional role of the cAMP-dependent pathway in clonidine-induced inhibition of SL gene expression, tilapia NIL cells were exposed to clonidine (1 μM) in the presence of the AC activator forskolin (1 μM) and cAMP analog CPT-cAMP (100 μM), respectively. In this case, addition of clonidine (1 μM) inhibited not only basal SL gene expression but also forskolin- and CPT-cAMP-stimulated SL gene expression (Figure 6D).
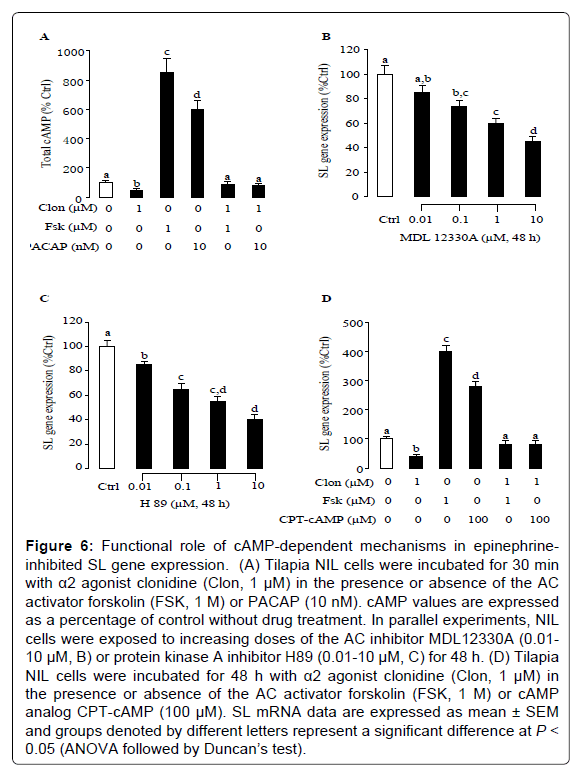
Figure 6: Functional role of cAMP-dependent mechanisms in epinephrine-inhibited SL gene expression. (A) Tilapia NIL cells were incubated for 30 min with α2 agonist clonidine (Clon, 1 μM) in the presence or absence of the AC activator forskolin (FSK, 1 M) or PACAP (10 nM). cAMP values are expressed as a percentage of control without drug treatment. In parallel experiments, NIL cells were exposed to increasing doses of the AC inhibitor MDL12330A (0.01- 10 μM, B) or protein kinase A inhibitor H89 (0.01-10 μM, C) for 48 h. (D) Tilapia NIL cells were incubated for 48 h with α2 agonist clonidine (Clon, 1 μM) in the presence or absence of the AC activator forskolin (FSK, 1 M) or cAMP analog CPT-cAMP (100 μM). SL mRNA data are expressed as mean ± SEM and groups denoted by different letters represent a significant difference at P < 0.05 (ANOVA followed by Duncan’s test).
Based on the studies in rainbow trout, epinephrine dosedependently inhibited SL release in the organ-cultured pituitary [20]. Since epinephrine does not readily cross the blood brain barrier [29], this catecholamine must directly act at the level of pituitary gland to inhibit SL secretion. This idea was further corroborated by our present studies that showed epinephrine could inhibit SL gene expression in a time- and dose-dependent manner, suggesting that SL response to epinephrine may represent a common phenomenon in fish models. To shed light on the receptor specificity for SL response to epinephrine at the pituitary level, we clarified the receptor specificity using a pharmacological approach. In the present study, epinephrine-induced inhibitory actions were mimicked by the α2- agonist clonidine and UK14304 but not by the α1-agonist cirazoline or β-agonist isoproterenol. Furthermore, epinephrine inhibition on SL gene expression could be prevented by the α2-antagonist yohimbine, whereas the α1-antagonist HEAT and the β antagonist propranolol were not effective in this regard. Apparently, α2 inhibition of SL gene expression could not be due to SL mRNA degradation as clonidine treatment did not alter the half-life of SL transcripts. These results, as a whole, provide evidence for the first time that the inhibitory action of epinephrine on SL gene expression is mediated by α2-adrenoceptors in tilapia NIL cells.
To further evaluate the functional role of epinephrine as a SL inhibitor in fish, we also examined functional interactions between PACAP and epinephrine in regulating SL gene expression. PACAP is a member of the secretin/glucagon/vasoactive intestinal polypeptide (VIP) family [30]. This peptide was first isolated in ovine hypothalamus based on its ability to stimulate AC activity in rat pituitary cells [31]. The mature peptide of PACAP reported in other vertebrate species, including fish, amphibians, and birds, exhibits more than 90% sequence homology when compared to the mammalian counterpart [32]. The role of PACAP as a hypophysiotropic factor is supported by the findings that: 1) PACAP nerve fibers are present in the median eminence and 2) PACAP can elevate basal secretion of GH, gonadotopin, prolactin, SL and adrenocorticotropic hormone [33]. In the present study, PACAP consistently stimulated SL mRNA expression in tilapia NIL cells, and these stimulatory actions could be blocked by epinephrine or the α2-agonist clonidine. Therefore, it would be logical to postulate that epinephrine may act as a negative regulator for PACAP induction of SL gene expression by acting through α2-adrenoceptors in the tilapia pituitary.
In general, α2-adrenoceptors are functionally coupled to Gi/Go G-proteins [34], which are negatively coupled with AC. However, the post-receptor signaling mechanisms for epinephrine-induced inhibition of SL gene expression are largely unknown. In tilapia NIL cells, α2-agonist clonidine treatment could inhibit total cAMP production and the ability of forskolin to increase cAMP production was also inhibited by clonidine. In our previous studies, PACAP-induced stimulation of SL mRNA expression can be attributed to the coupling of the AC/cAMP/PKA system [26]. These findings have prompted us to speculate that epinephrine may interfere with these signaling pathways to inhibit the stimulatory actions of PACAP on SL gene expression. This hypothesis was supported by the results of direct measurement of cAMP production in tilapia NIL cells. In the present study, PACAP consistently elevated cAMP levels in tilapia NIL cells, and these stimulatory actions could be blocked by simultaneous treatment with the α2-agonist clonidine. In parallel experiments, clonidine inhibition on SL mRNA expression could be mimicked by blocking cAMP-dependent pathway with the AC inhibitor MDL12330A and the PKA inhibitor H89. Furthermore, the α2-agonist clonidine was effective in abolishing the SL responses to CPT-cAMP and forskolin, suggesting that epinephrine may interfere with the AC/cAMP pathways to inhibit SL gene expression. These results, as a whole, suggest that α2-adrenergic stimulation can downregulate SL gene transcription by inhibiting the AC/cAMP/PKA pathway in tilapia NIL cells.
In summary, we have demonstrated for the first time that epinephrine can suppress SL gene expression by acting directly at the pituitary cell level. These inhibitory actions are mediate through α2- adrenoceptors negatively coupled to the cAMP-dependent pathway. This adrenergic inhibition not only affects basal SL gene transcripts but also attenuates the SL response to PACAP. Since PACAP is an important physiological SL-releasing factor in fish, the present findings may suggest that the α2 inhibitory influence at the pituitary level is an integral component of the neuroendocrine control of SL gene expression.
The project was supported by grants from the National Natural Science Foundation of China (31302165). We are grateful to Dr. Yajun Wang (Sichuan University) for providing technical assistance.
No conflicts of interest, financial or otherwise, are declared by the authors.