Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Review Article - (2013) Volume 2, Issue 2
Keywords: Industrial enzyme, Promoter, Copy number of genes,Signal peptide, Codon bias
Traditional monospore isolation, mutation breeding and optimization of fermentation process have been widely used to improve the production of industrial enzymes. However, these methods have obvious disadvantages, such as difficulty in obtaining the purebred microorganism, little favorable mutation and the long fermentation period, etc. With the development of genetic engineering, a lot of studies have been carried out to improve the production of industrial enzymes from the molecular level. Genes of some enzymes, such as glucoamylase, amylase, lipase, cellulase and phytase, etc., have been cloned and expressed efficiently in heterologous host systems, such as Saccharomyces cerevisiae; Aspergillus niger and Pichia pastoris [1-11]. In this paper, advanced strategies for improving the production of industrial enzymes were reviewed from the following aspects: the introduction of the strong promoter, increasing the copy number of genes, changing the signal peptide sequence and designing the codon bias (Figure 1).
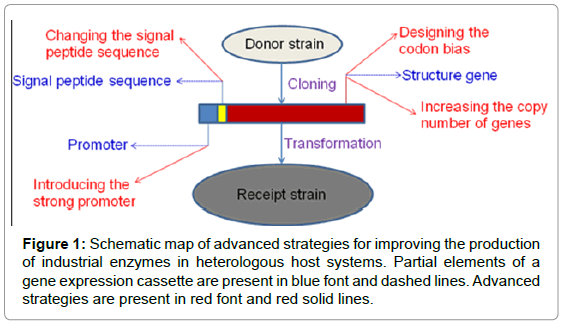
Figure 1: Schematic map of advanced strategies for improving the production of industrial enzymes in heterologous host systems. Partial elements of a gene expression cassette are present in blue font and dashed lines. Advanced strategies are present in red font and red solid lines.
Use of the strong promoter to improve the production of industrial enzymes
Introducing exogenous genes to host strains often results in strains that cannot start the expression of exogenous genes. As a result, the expression of exogenous genes is low. This can be resolved by introducing strong promoters as listed in Table 1 when expression vectors are constructed.
| Abbreviation | Full name | Sources |
|---|---|---|
| PGK | phosphoglycerate kinase 1 promoter | Mouse |
| GDP | glyceraldehyde-3-phosphate dehydrogenase promoter | S. cerevisiae |
| ADH1 | alcohol dehydrogenase promoter | S. cerevisiae |
| AOX1 | alcohol oxidase promoter | S. cerevisiae |
| GAP PHO5 GAL1 SUC2 |
glyceraldehyde-3-phosphate dehydrogenase promoter acid phosphatase promoter galactokinase promoter sucrose invertase promoter |
P. pastoris S. cerevisiae S. cerevisiae S. cerevisiae |
Table 1: Strong promoters used in constructing industrial enzymes-producing engineered strains.
Hyung et al. [12] introduced the yeast sucrose invertase SUC2 promoter and the glucoamylase gene sta into S. cerevisiae cells through the plasmid construction, improving the production of glucoamylase. Glucoamylase can be also effectively produced by cloning an Aspergillus oryzae glucoamylase gene into a high-copy yeast shuttle plasmid, using the glyceraldehyde-3- phosphate dehydrogenase (GPD) promoter and its own signal peptide sequence [13]. Genes encoding mouse salivary α-amylase and glucoamylase were inserted into the plasmid pMS12 that used a strong alcohol dehydrogenase (ADH1) promoter, and this leads to the effective production of them in S. cerevisiae. In the last five days of the transformant culture, the average conversion rate of starch was 93% [14]. Ma et al. [15] integrated a strong promoter and the lipase-coding cDNA from Bacillus subtilis IFFI10210 into the original plasmid pBD64 to build a new plasmid pBSR2, transformed it into the B. subtilis A.S.1.1655 strain, and obtained the lipase-producing strain BSL2 whose yield was 8.6 mg/g (cell dry weight), 100-fold higher compared to the original strain. The structural gene encoding the super thermo- and acid-stable α-amylase was amplified from an extremely thermophilic ancient bacterium Pyrococcusfuriosus, and was introduced into P. pastoris GS115 by constructing the expression vector pPIC9K under the control of the α-factor signal peptide and the AOX1 promoter [16]. The screened transformant can secrete 3000 U/ml of amylase after the methanol induction for 7 d. Liu et al. [17] constructed a recombinant vector that contained a gene encoding cellobiose hydrolase, and integrated it into the chromosome of P. pastoris GS115. Owning to the use of the strong promoter AOX1, the screened strain can produce up to 1.2 mg/ml of cellobiose hydrolase.
Increasing the copy number of genes to improve the production of industrial enzymes integrating a multi-copy gene into the chromosome of A. niger or P. pastoris can obviously improve the production of industrial enzymes. Verdoes et al. [18] investigated the relationship of the copy number of the glucoamylase gene and its expression. The result showed that the production of glucoamylase could be improved from 50 mg/ml to 900 mg/ml within 20 copies of its gene. However with further increase in the gene copy numbers, the glucoamylase yield decreased gradually and recombinant strains became genetically unstable. The fermentation of the recombinant strain A. niger B1 bearing 20 copies of the glucoamylase gene was performed to obtain a maximal glucoamylase production of 1510 mg/(g•h) at a maximal growth rate [19]. The multi-copy glucoamylase gene from bacterium was integrated into the chromosome of strains A. awamori and A. niger [20] glucoamylase activities of recombinant strains in the liquid fermentation were 2-fold higher compared to the original strain, and they were improved by 85% in the solid-state fermentation. The glucoamylase gene glaA from A. niger F0410 was cloned, and the expression vector bearing a multi-copy glaA was constructed and transformed into A. niger. The screened transformant can obviously improve the production of glucoamylase. After the pilot experiment in a 30 l fermentor, the glucoamylase activity reached 51000 IU/l in the recombinant strain GB0506 [21]. The result indicated that the glucoamylase activity could be improved in the industrial strain A. niger F0410 that the multi-copy glaA were integrated into its chromosome. Moreover, 2-3 copies of glaA are appropriate for the glucoamylase overproduction. However, some studies showed that the glucoamylase yield was present in a nonlinear relationship with the copy number of glaA integrated into the chromosome of A. niger F0410. This may be perhaps because integrated sites of glaA and the number of transcriptionally regulatory proteins limit the secreted production of glucoamylase. Ding et al. [22] performed the electro-transformation to the GS115EG11 strain into which a neutral endocellulase gene eg1 had been integrated, and obtained 60 positive transformants from YPDSZ plates containing 2000μg/L of zeocin. This greatly improves the transformation rate, and finally a strain GS115MEG1 that could effectively produce the neutral endocellulase was successfully screened. The production of the neutral endocellulase in this strain was increased by 3.8-fold compared to the original GS115EG11 strain at the flask level. Luo et al. [23] modified the P. pastoris expression vector pGAPZα-A to be methanol-inducible. This vector can be integrated into the GAP locus of the chromosome of P. pastoris. Using the strain 74# containing a phytase gene appA_m that had been integrated into the AOX1 locus of its chromosome, as the recipient strain, the recombinant strain that integrated another phytase gene into the same locus could be obtained by electroporation. The phytase yield in the screened recombinant strain in a 5 l fermentor reached 4 g/l. Wang et al. [24] ligated the phytase gene from A. niger N25 with the expression vector pPIC9K to construct pPIC9K-phyA, and transformed it into P. pastoris GS115. The transformant that included high-copy phytase genes was screened by increasing G418 concentration. This strain can produce 35646.7 U/ ml of phytase.
Improving the production of industrial enzymes by designing the codon bias
Genetic code has degeneracy and usually an amino acid is encoded by 2-6 codons. The usage of synonymous codons shows a big bias in every organism, which is very obvious for highly expressed genes. There are many reasons which result in the intraspecific codon bias, and the tRNA abundance is considered to be the most important. In highly expressed genes, the codon usage should match with the number of tRNA, possibly making use of codons which carry many tRNA molecules, and thus speeding up the rate of the protein synthesis [25]. When heterologous genes are expressed in host cells, a low or no production of exogenous proteins is often presented due to different codon biases. This can be resolved by replacing rare codons by hostpreferred ones and increasing the amount of rare tRNAs [26]. Amino acids of C. rugosa lipase contain 47 serine residues. More than 20 serine residues including Ser209 located in the active center are encoded by CUG, a codon that often encodes Leu in most organisms. The active lipase cannot be obtained when the C. rugosa lipase gene is directly expressed in S. cerevisiae cells. Based on the codon bias of S. cerevisiae and P. pastoris, serine-encoding codons and the 5’-UTR region of the C. rugosa lipase gene were optimized to make it achieve the effective expression in the two host cells. This greatly shortens the fermentative cycle [27,28]. Wei et al. [29] optimized a thermostable α-amylase gene (pfa) sequence based on the Pichia codon bias and artificially synthesized it. This sequence was ligated to the vector pPIC9 to construct the plasmid pPIC9-pfa. After being linearized, this plasmid was transformed into the chromosome of P. pastoris by electroporation to obtain the recombinant strain. The screened strain can secrete the highest amylase activity of 220 U/ml, 2-fold higher compared to the recombinant E. coli (109 U/ml). The phytase gene phyA2 from A. niger 963 was modified by removing its introns and signal peptide encoding sequence, and optimizing Arg codons that play a key role in its efficient expression by site-directed mutagenesis. The modified phytase gene was fused to the 3’-terminal of the α-factor signal peptide encoding sequence of the expression vector pPIC9, and was transformed into P. pastoris GS115 by electroporation. Copy numbers of the modified phytase gene integrated into the chromosome of P. pastoris were confirmed by southern-blotting analysis. This gene can be normally transcribed and expressed in host cells. The phytase activity in the screened transformant is up to 15000 U/ml, 37- and 3000-fold increases compared to the screened strain with normal Arg codons and the original strain, respectively [30]. Chen et al. [31] optimized two Arg codons of the conserved sequence R81HGXR85XP from the active site of phytase using the Pichia-preferred Arg codon AGA to replace low-frequent codons 81-CGT and 85-CGG. The phytase activity in the codon-optimized strain is 47600 U/ml, 1-fold higher than that of the original strain. These results indicate that Pichia prefers to use biased codons. This to some extent enhances the translation rate, and hence increases the enzymatic activity.
Impact of signal peptide sequences on the production of industrial enzymes
The N-terminus of the secreted protein often contains a leader sequence comprising of 15-30 amino acids. This sequence is called the signal peptide which includes 2-3 polar amino acids at or near its N-terminus. The central sequence of the signal peptide is a sole hydrophobic core, or consists of many hydrophobic amino acids. A significant character of the signal peptide is that it can form α-helical structure by the action of hydrophobic regions. It has been demonstrated that the modification of the signal peptide can improve the expression of heterologous genes [32]. Liu et al. [33] used the MF-α signal peptide from S. cerevisiae to replace the one of glucoamylase itself, and found that the secreted production of glucoamylase was improved by 3.6- and 56-fold when its copy numbers were 1 and 7, respectively. The high phytase-producing recombinant A. niger can be obtained by cloning its gene (phyA) into the downstream from the amyloglucosidase promoter from A. niger. Two recombinant strains bearing AG signal peptide sequences comprising of 18 or 24 amino acids can secrete 1.1 and 0.5×105 U/ml of phytase, and the third one bearing the signal peptide of phytase itself can secrete 2.8×105 U/ml of phytase. Phytase yields in three strains have a significant improvement compared to the original strain (100 U/ml) [34]. Effect of signal peptide sequences on the secreted production of heterologous proteins in P. pastoris was investigated in detail by Xiong et al. [35]. S. cerevisiae α-factor signal peptide sequence MF4I was first synthesized based on the Pichia codon bias, and then 1-10 N-terminal amino acids of the Pichia AOX1 protein were respectively introduced into the N-terminus of MF4I to construct 10 different signal peptide sequences that are used for the production of phytase. All of these signal peptides significantly improve the production of phytase, especially one whose N-terminus includes A, I and P. Compared to the original signal peptide from widetype S. cereviaiae, the average production of phytase was improved by 5-fold in screened strains at the shake flask level (90 mg/l). Moreover, increasing 10 amino acids (EEAEAEAEPK) between MF4I signal peptide and endoprotease can further improve the secreted efficiency of the neutral phytase, and this makes its production improved by 35% (120 mg/l).
Given that industrial enzymes are widely used in many fields, studies on improving their production have been a hot topic. In this paper, advanced strategies, including the introduction of the strong promoter, the increase in the gene copy number, the use of preferred codons and the alternative to the original signal peptide, were reviewed to improve the production of industrial enzymes. There will be an enormous potential for improving the production of industrial enzymes in heterologous host systems by above-mentioned strategies in future.
This study is supported by the National Natural Science Foundation of China (No.31171658).