Advanced Techniques in Biology & Medicine
Open Access
ISSN: 2379-1764
ISSN: 2379-1764
Review Article - (2013) Volume 1, Issue 2
Gene silencing by RNA interference has become a very promising technique in the fields of biomedical and life sciences during the past few years. In this approach, small interfering RNA molecules that specifically bind to and cleave transcripts of target genes are introduced into cells, thereby causing full or partial loss of gene function, i.e. comprising null or hypomorphic phenotypes. That is why accession to and characterizations of genes that are associated with diseases and/or disease progression has become easier and faster in recent times. In this review the discovery of gene silencing by RNA interference, its molecular mechanism, optimal design principles of small interfering RNAs, advantages and barriers of RNA interference as a gene silencing tool, and finally its therapeutic applications with outcomes in some clinical trials with future aspects including oncolytic viruses will be discussed.
Keywords: RNA interference; Gene silencing; Recombination
One of the major ways for determining a proteins biological function is to examine what phenotype it produces in an organism that carries a mutation in its gene. ‘Forward genetics’ refers to the process where collections of randomly mutagenized organisms like worms and flies are first screened for specific phenotypes and the responsible gene mutations are identified afterwards. Whereas, ‘reverse genetics’ describes the process where specific gene mutations are first introduced into the genomes of organisms like yeasts or mammalian cells by methods like homologous recombination and the resulting phenotypes are analyzed afterwards. Alternatively, protein function can also be investigated by completely removing its gene from the genome and then analyzing the resulting phenotype, a process also known as ‘gene knockout’; but, this technique can sometimes be very time consuming and expensive. Another way for determining a proteins function is to inhibit its synthesis at the ribosomes by targeting its mRNA transcripts and observing the resulting phenotype. This process is called ‘gene knockdown’ and can be either achieved by hindering the maturation process of mRNAs in nucleus with antisense oligonucleotides or by cleaving mature mRNAs in the cytoplasm by specific enzymes called ribozymes. Although useful, these techniques also have their limitations for wide usage. However, today a much easier, faster and more powerful gene knockdown tool has revolutionized the field of gene silencing: RNA interference (RNAi). RNAi is an evolutionary conserved post-transcriptional gene silencing mechanism which allows cells to specifically destroy exogenous mRNAs in their cytoplasm, like that from infectious viruses, and is easily applicable to many different cells and organisms.
In the year 1990, initial evidence for the existence of a novel and until then unexploited gene silencing mechanism came from genetic modification studies in plants. In these studies, researchers aimed to deepen the purple color of petunia flowers by overexpressing chalcone synthase (CHS), the enzyme responsible for pigment synthesis. For this purpose, cells were transfected with multiple copies of the CHS gene; but, instead of getting deep purple petunia flowers they ended up with variegated or white ones [1,2]. Somehow, the introduced transgenes were silenced as well as the plant’s endogenous purple-flower gene, a phenomenon called ‘co-suppression’ [3]. Later, similar observations were made in the fungus Neurospora crassa and the nematode worm Caenorhabditis elegans. When in 1998 the developmental biologists Andrew Fire and Craig C. Mello, who also worked with the model organism Caenorhabditis elegans, incorporated this approach into their studies the explanation for the process came through and the term “RNA interferance” emerged the first time [4]. They showed that when injecting double stranded RNAs (dsRNAs) into Caenorhabditis elegans cells, downregulation of protein expression could of be achieved by binding of the complementary RNA molecule to the corresponding mRNA leading to its degradation. Their results have also pointed out that; single-strand RNAs were less likely to inhibit the expression of that target gene rather than dsRNAs and the introduction of doublestranded RNA caused homology-dependent post-transcriptional gene silencing. Andrew Fire and Craig Mello got the Nobel Prize in 2006 in Physiology or Medicine for their discovery of RNA interference–gene silencing by double-stranded RNA. The molecular mechanism of RNAi was further unveiled in an in vitro model of the fruit fly Drosophila melanogaster. By using this system it could be shown that in the cells the long dsRNAs were cut into small fragments, which were 21 to 23 nucleotides long, and that actually these resulting small dsRNAs lead to the degradation of target mRNAs [5]. Until that time, all RNAi studies were accomplished in invertebrates and initial attempts to apply it to mammalian cells were unsuccessful. Then, in mammalian cells dsRNAs evoked an innate immune response that was characteristic for viral infections leading to the production of interferons and the inhibition of whole gene expression, followed by rapid cell death. Therefore, it first seemed that RNAi have totally been lost and replaced by a more recently evolved interferon system that did not exist in invertebrates [6]. However, there were some clues that RNAi might still exist in mammals. The breakthrough came when dsRNA molecules less than 30 nucleotides long were introduced into mammalian cells, without inducing an interferon response by the innate immune system. Also, inhibition of whole gene expression did no longer occur with these “small interfering RNAs” (siRNAs) and they still kept their ability of directing sequence specific degradation of complementary mRNAs very similar to that in plants, worms, and flies [7]. In brief, the term siRNA today describes all short dsRNAs that are typically 21 to 25 nucleotides long, specifically bind their target mRNA and induce their degradation through the RNAi system. The most effective siRNAs have a two nucleotide overhang at their 3’ ends [8].
siRNAs that induce the degradation of specific endogenous mRNAs are known as a beneficial biologic mechanism that is widely recruited by eukaryotic cells to inhibit protein production at post transcriptional level. RNAi can result in gene silencing or even in the expulsion of sequences from the genome. The RNAi process is initiated by short dsRNAs (21-25 nucleotides in length) that lead to the sequence specific inhibition of their homologous mRNAs. These siRNAs are normally produced in cells from cleavage of longer dsRNA precursors by Dicer: the ribonuclease III (RNase III) family member [9]. The cleaved parts are incorporated into a multi-component nuclease complex known as the RNA-Induced Silencing Complexes (RISC), which has the splicing protein Argonaute-2 (Ago-2) [10]. Then, the single stranded RNA derived from the short dsRNA acts as a guide sequence (the antisense strand) directing the complex to the specific target mRNA by intermolecular base pairing principle; in where a RISC-associated endoribonuclease cleavages the target mRNA and thus; allows greater efficiency of gene extinction [11-14] (Figure 1).
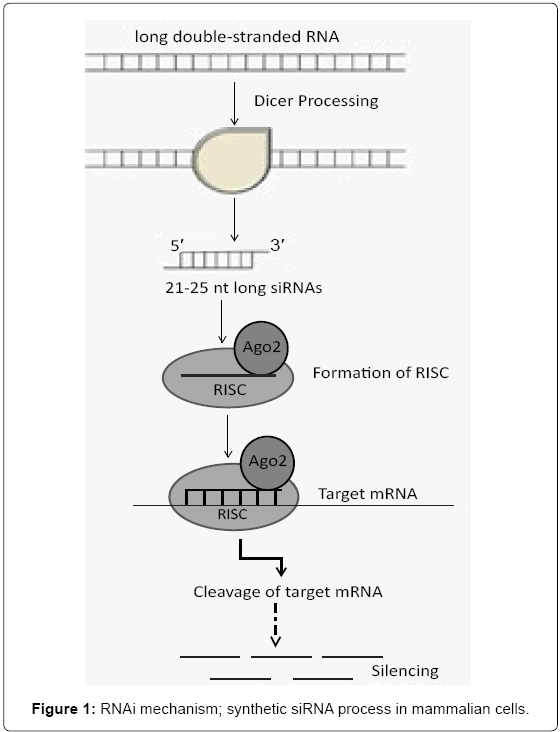
Figure 1: RNAi mechanism; synthetic siRNA process in mammalian cells.
siRNAs with 2 nucleotide overhang on their 3’ ends were found to be more efficient in reducing the expression of a target mRNA than siRNAs with blunt ends [3]. There are some recommendations in siRNA design approach: 1) The siRNA sequence should begin with the start codon AUG among the target gene transcript, AA dinucleotide sequences should be found downstream way and therefore; each AA and the 3 following 19 nucleotides are potential siRNA targets. 2) The sequences of the potential target sequences should be compared to sequences in the species from an appropriate genome database (www.ncbi.nlm.nih.gov/BLAST/) 3). Any target sequences that are homologous to other coding sequences should be eliminated. 4) Three or 4 different sequences along the length of the target gene for production of siRNAs should be selected to constitute an effective siRNA pool [15]. Further analyses pointed out that, application of an algorithm incorporating all eight criteria significantly improved potent siRNA selection [16]. There are also siRNA construct builder programs as “vector-based siRNA and siRNA cassettes” providing an all-inclusive utility tool in order to indicate the specific bioinformatics needed for siRNA design [17].
RNAi phenomenon was described as “Breakthrough of the Year” by Science journal in 2002, for having the potential of being a powerful therapeutic drug. RNAi-based therapeutics allow analysis in a matter of days, and enables to effect loss of gene function at cellular level that would have taken several months or even years by previous methods such as homologous recombination. It has also significant advantages over traditional approaches in curing diseases by having wide application areas, therapeutic susceptibility and selectivity avoiding side effects. This widespread applicability, relative ease of synthesis and low cost of production make siRNAs an attractive new class of smallmolecule drugs [6].
Among the RNAi triggering agents, synthetic siRNAs have been widely investigated and evaluated as a potential drug to overcome human genetic disorders since a siRNA can be designed to specifically target any disease- causing gene in various cell types [18]. The outcomes taken from in vitro and in vivo human studies carried out with synthetic siRNA have shown that long-term silencing of target genes was achieved without damaging the endogenous miRNA pathways [19] and have consistently deduced that, RNAi therapeutics could be an outstanding candidate for future clinical use. RNAi-based drugs are designed to destroy the target mRNA and therefore stop the associated protein production required for disease progression.
Ever since the first report by Elbashir et al. [7] have pointed out an efficient gene silencing achieved via transfection of synthetic siRNA with Lipofectamine 2000 in cultured human cells; significant improvement has been seen in siRNA delivery techniques with a short period of time. In another study reported by Kawasaki and Taira [20], expression of mutant p53 or RAS genes in SW480 human colon cancer cells were specifically silenced by synthetic siRNAs without affecting the wild type genes. In another study, target of siRNAs to human telomerase RNA inhibited telomerase activity in variety of human cancer cell lines by Kosciolek et al. [21]. In some other pioneering studies, researchers have demonstrated great possibilities for treating serious diseases caused by HIV and HCV viruses by using synthetic siRNAs [22,23].
Despite these numerous successful studies of RNAi on inhibition of specific mRNAs in the mammalian tissue culture system, the efficacy of each cell line is not identical and even very difficult in primary cell lines. Therefore; there are still some limitations such as inefficient delivery system to desired cell type-tissue-organ, poor intracellular uptake, stability, cytotoxicity possibility, off-target effects and immune responses that have to be overcame in RNAi therapeutics for both in vitro and in vivo [24-26].
As for using synthetic siRNAs as research tools, their therapeutic activity and pharmacokinetic properties should be increased. Delivery matters such as plasma stability, cellular uptake and disease targeting generates pharmacokinetic properties. In order to achieve maximum therapeutic effect, the most active form of drugs should be used [27]. In this purpose, rational design of siRNA and finding the right delivery tool system seems to be the key points [28-30].
Delivery limitations
Although siRNA has small size, its transportation across the cellular membrane is got hardened due to hydrophilicity and negative charge. In addition, siRNA is quickly cleared during in vivo circulation, before reaching the target disease site [31]. Following the application of siRNA therapeutics into mammals, they face various extra- and intracellular barriers. Thus, the therapeutic effect of siRNA therapeutics can be influenced by the administration route either with local or systemic delivery. Between them, systemic administration of siRNA therapeutics seems to be easier since there is no limit on target tissue or organ accessibilities in contrast to the local delivery system [32,33] but still these two different delivery systems have their own advantages and disadvantages [28-30,34]. As for non-viral vectors used for siRNA delivery, their useful forms were achieved by lipids (cationic liposomes, neutral liposomes) and polymers (polyethyleneimine-PEI, nanocapsules, dendrimers) [35]; moreover, hydrodynamic approaches by injecting siRNA [36], electroporation [37] and the gene gun as local delivery system [38] were also tested other techniques.
Stability and off-target effects
The other limitations of siRNA applications are stability, off-target effect and immune stimulation. siRNAs are sensitive to enzymatic degradation in vivo; and, since siRNA itself is relatively unstable inside the blood stream, they have a short half-life of less than ~15 min under physiological conditions [39]. The introduction of synthetic siRNA may also cause undesired off-target gene silencing effects by having “partial homology” instead of “perfect match” with target mRNA sequence. In addition, siRNA duplex can activate immune stimulation via the engagement with the major components of the innate immune system [40]. Indeed, dsRNA strings can trigger non-specific cellular innate immune response such as the interferon response. Hornung et al. [41] have demonstrated that dsRNAs longer than 30 nucleotides induced interferon response by binding to double stranded-RNAactivated protein kinase (PKR), 2’, 5’-oligoadenylate synthetase-RNase L system or several Toll-Like Receptors (TLRs). The experimental analyses whether interferon response was created or not could be determined by checking the expression level of an interferon-response gene such as oligoadenylate synthase-1 (OAS1) [42,43]. Another machinery triggering non-specific effect was, the saturation of RNAi process due to using high concentrations of siRNA and eventually causing cytotoxicity upon cells. Therefore, it is important to transfect the minimum amount of the siRNA duplex to eliminate the off-target effects [44].
In the last decade, various chemical modifications of siRNA have been attempted to overcome these inherent problems of siRNA therapeutics. Indeed, cautious chemical modifications made on the siRNA backbone could improve its physico-chemical properties that are required for RNAi without loss of gene-silencing efficiency [45-47]. For example, insertion of a 5' to 3’ phosphorothioate (P=S) linkage and the modification of 2’-O-methyl and 2’-fluoro for exonuclease and endonuclease resistance increased serum stability [45,47]. Also, it was demonstrated that the 2’-O-methyl modification in the seed region of siRNA reduced its off-target activity without compromising the ability of silencing the target genes [45]. Other valuable studies also give more precise and informative description of various chemical modifications in siRNA therapeutics [40,48,49]. Besides these, we have also used chemical modifications in order to increase silencing efficiency in primary cell line K-562 with cholesterol conjugation resided at the 5’- end of siRNA sense strand and substitution of 2’-FluoroUridine (2’- FU) with 2’-Fluorocytidine (2’-FC) which is applied to RNA’s 2’-ribose region (sugar modification); resulted in silenced target STAT genes efficiently in leukemia cells for a duration of 12 days both at mRNA and protein levels [50].
As for uses and benefits of RNAi is an accelerating potent in medicine. There have been many probable gene targets for therapeutic intervention using RNAi; and therefore, this approach has an advantageous process to cure diseases that thought to be occurring due to the presence of single defective gene [51]. The first application of RNAi therapy was carried out for age related macular degeneration (AMD) by using siRNAs to suppress the Vascular Endothelial Growth Factor (VEGF) pathway that causes abnormal growth of blood vessels behind the retina. This application was designed to be administered directly to the patient’s eye [52].
We will focus on different types of diseases that are widespread and for which RNAi approaches are currently being tested in preclinical studies. While RNAi mediated in vitro or transgenic mice studies used for inhibiting specific genes linking to cancer or human disorders are given in Table 1, examples of current trials for RNAi therapy can be found in Table 2. The advantage and disadvantage of each virus taking place in siRNA applications is summarized in Table 3 from the standpoint for clinical use.
| Targeted Gene | RNAi processing Type | in vitro/in vivo model | Final Outcome | Efficiency Duration | Year; Reference |
|---|---|---|---|---|---|
| Anti-hepatisis B virus pre-miR 31 | Pol II cassettes encoding primary (pri)-miR-31 | HuH-7 cells & a Mouse model | HBV replication was inhibited | 5 days | 2008; [76] |
| NS5-1, NS5-2, E NS1 genes causes of yellow fever virus | Short hairpin RNA (shRNA) plasmid | Vero E6 cells & Mouse model | YFP expression was inhibited | 10 days | 2009; [77] |
| Influenza M2 gene | shRNA cassettes | Madin-Darby cannie kidney and human embryonic 293T kidney cells | Influenza A virus replication was inhibited | 3 days | 2009; [78] |
| Insulin-like growth factor-I receptor (IGF-IR) in colon cancer | shRNA plasmid vector | Human colon cancer cell line SW480 | IGF-IR reduction and tumor growth inhibition | 2 days | 2009; [79] |
| Bcl-2 gene | Human telomerase RT promoter expressing mi-Bcl2 | Lung cancer cell lines: A549, Hela-S3, HepG2 | Suppression of Bcl-2 & induction of apoptosis | 7 days | 2009; [80] |
| c-myc oncogene in gastric tumor | Vector based siRNA | Human gastric cancer cell line SGC7901 & gastric cell line HFE145 | Knockdown of c-myc prevented growth and proliferation of gastric cancer cells | 7 days | 2010; [81] |
| APE1/Ref-1 gene related with human pancreatic cancer | Chemically modified synthetic siRNA | Human pancreatic cancer cell line SW1990 | Due to expessional downregulation of APE1/Ref-1, chemotherapy response was increased, apoptosis was induced | 3 days | 2010; [82] |
| EZH2 gene in tumorigenesis and liver metastatic pancreatic cancer | shRNA plasmid vector | Human pancreatic cancer cells SW1990 and PANC-1 & Athymic nude mice | EZH2 expressional silencing inhibited tumor growth in vivo | 45 days | 2010; [83] |
Table 1: RNAi mediated in-vitro or in-vivo studies used for inhibiting specific genes linking to cancer or human disorders [75]
| Drug | Target Disease | Target Gene | Phase/Patient number | Company |
|---|---|---|---|---|
| Atu027 | Advanced solid tumor | PKN3 | Phase I/33 | Silence Therapeutics AG |
| CALAA-01 | Solid tumor | M2: ribonucleotide reductase subunit | Phase I/36 | Calando Pharmaceuticals |
| siRNA-EphA2-DOPC | Advanced cancer | EphA2 | Phase I/40 | M.D. Anderson Cancer Center |
| PRO-040201 | Hypercholesterolemia | APOB | Phase I/23 | Tekmira Pharmaceuticals |
| TKM-080301 | Primary/secondary liver cancer | PLK-1 | Phase I/42 | National Cancer Institute |
| SYL1001 | Ocular pain dry eye | TrpV1 | Phase I/30 | Sylentis, SA |
| SYL040012 | Glaucoma | β2 adrenergic receptor | Phase I/30 | Sylentis, SA |
| Ocular hypertension | ||||
| QPI-1007 | Optic atrophy | Caspase 2 | Phase I/66 | Quark Pharmaceuticals |
| Non-arteric anterior | ||||
| Ischemic optic neuropathy | ||||
| I5NP | Injury of kidney | P53 | Phase I/16 | Quark Pharmaceuticals |
| Acute renal failure | ||||
| SPC2996 | Chronic lymphocytic leukemia | Bcl-2 | Phase I-II/46 | Santaris Pharma AS |
| Bevasiranib | Diabetic macular edema | VEGF | Phase II/48 | Opko Health, Inc |
| Macular degeneration | ||||
| ISIS104838 | Rheumatoid arthritis | TNF-α | Phase II/160 | Isis Pharmaceuticals |
| P04523655 | Choroidal neovascularization | RTP801 | Phase II/184 | Quark Pharmaceuticals |
| Diabetic macular edema | ||||
| Diabetic retinopathy | ||||
| Alicaforsen | Chron’s disease | ICAM-1 | Phase III/150 | Isis Pharmaceuticals |
Table 2: Current Approaches of clinical trials upon RNAi based Therapeutics [17,82,83].
| Virus Type | Content of the study | Advantages | Disadvantages | Literature |
|---|---|---|---|---|
| HBV | Phase Ib trial: plasmid encoded shRNA against 4 different HBV subtypes was delivered to 3 patients | The delivery system was safe and tolerable | Application showed only a modest effect against HBV Evidence of an anti siRNA immune response was detected in patients receiving siRNA | [101] |
| HIV | A lentiviral construct was developed that used shRNA to inhibit CCR5 expression shRNA for tat/rev genes of the virus and a decoy TAR sequence to inhibit HIV polymerase | For delivery of the lentiviral construct, haematopeitic stem cell transplantation was used that was being undertaken to treat patients for AIDS related lymphoma Patients received bone marrow transplantations of their own cryopreseved CD34+ stem cells They were able demonstrate long term engraftment up to 2 years, demonstrating the feasibility of the procedure to safely block viral replication Using patients own cells with lentiviral delivered shRNA therapy is one of the most promising for AIDS treatment | A small proportion (0.2%) of cells were transduced with the lentiviral construct due to safety concerns about the impact of the lentivirus on normal cell function Impact of the therapy on viral replication was unable to be assessed | [102, 103] |
| HCV | Phase 2a clinical trial with miravirsen (SPC3649) to assess safety and tolerability in treatment-naïve 36 patients with chronic HCV | These clinical studies pave the way to designing effective RNAi strategies in patients Drug inhibits miR-122, a microRNA important for Hepatitis C viral replication, thereby significantly reducing the levels of Hepatitis C virus Miravirsen given as a four-week monotherapy treatment provided robust dose-dependent anti-viral activity with a mean reduction of 2 to 3 logs from baseline in HCV RNA (log10 IU/mL) that was maintained for more than four weeks beyond the end of therapy | headache and a hand abscess among the placebo-treated patients loss of consciousness in one patient 9 weeks after the last dose of 7 mg of miravirsen/kg, which occurred after a fall and also resulted in pelvic bone injury | [104] |
Table 3: The advantage and disadvantages of siRNA applications targeting HBV, HIV, HCV viruses in clinical applications.
Cancer
A life threatening disease cancer led the scientists direct to new and alternative curative strategies. Although point mutations are common in epithelial tumors, activated chimeric fusion onco-proteins generated by chromosomal translations are seen in hematological malignancies [53]. Since proto-oncogenes and oncogenes are activated in various types of cancer, there is an uncontrollable proliferation of cancerous cells. Another typical feature of cancer cells is being resistant to programmed cell death as a result of abnormalities in proteins that direct apoptosis [54]. The significance of RNAi application can be seen in cancer therapy in order to knock down the expression of either a cell cycle regulatory gene and/or an anti-apoptotic gene found in a cancer cell that gives rise to inhibition of tumor growth and inducing apoptosis. It has great importance to develop a therapy directly affecting cancerous cells; but not the healthy ones [26]. To selectively affect cancer cells without damaging normal cells, RNAi technology will be beneficial for targeting a gene of interest or deliver specifically designed siRNAs into cancer cells.
The studies revealed by RNAi therapy against cancer cells is used in for directly targeting the oncogenes; and therefore, stop progression and invasion of the tumor cells, and also increase the sensitization of tumor against drug [55].
Another important progress of RNAi was reported in leukemia and lymphoma. In this study, inhibition of multiple oncogenic gene fusion and suppression of disease development was achieved via siRNA transfection [56]. In a preclinical model, it is reported that; siRNA was keen on resisting the development of tumor by targeting cellular p53 gene that is involved in cancer development [57].
RNAi technique can be also used against the spread of tumor growth giving rise to an increase in tumor cell’s sensitization towards commonly used drug for treatment.
Experiments revealed on breast cancer cells indicated that; anti- CXCR4 siRNA (CXCR4; a chemokine receptor) treatment was capable of blocking the further expansion of breast cancer, by suppressing the function of the target gene [58]. In another study, it was shown that, tumor cells became sensitized to chemotherapy agents since siRNA applications suppressed the function of the anti-apoptotic bcl-2 gene [59].
RNAi treatment for HIV
HIV was the first infectious agent targeted by RNAi process. Today, HAART is the only technique to combat against HIV which can only extend the life span of the affected people; but prolonged usage might cause cytotoxicity [60]. So, RNAi is a promising technique against HIV to find an alternative way. Targeting early and late HIV-encoded RNAs via synthetic siRNAs in cell lines and in primary hematopoietic cells resulted with inhibition of them [61,62].
It is thought that HIV can interact with siRNA and make them resistance against the silencing mechanism. So, in order to avoid this, targeting the cellular receptors and co-receptor (CD4, CCR5 and CXCR4) that are responsible for the entry of HIV into the host cell was carried out and the expression or replication of the virus was prevented [63]. When siRNA application was specified for either p24 gene of HIV-1 or CCR5 co-receptor before infecting in a non-dividing macrophage; it resulted with suppression of infection up to 20 days and two siRNA targets for CCR5 co-receptor has been found to be efficient [64]. Finally in a humanized mice study, when they challenged the mouse with HIV, following anti-CCR5 siRNA treatment; resistance to infection was enhanced, which was assessed by the reduction in plasma viral load and disease associated CD4 T-cell loss. This finding pointed out the importance of RNAi-mediated CCR5 silencing by LFA-1-targeted nanoparticles by preventing HIV Infection in mice [65]. Hopefully, RNAi mediated knock down of gene expression offers a novel alternative therapy for overcoming HIV infection.
Hepatitis
Hepatitis is an inflammation of the liver and caused by Hepatitis B Virus (HBV) infection that threat to health and increase the risk of chronic liver disease and hepatocellular carcinoma in human. RNAi approach was used to inhibit the production of HBV in mice which are transfected with a HBV plasmid. In Immunohistochemical analyses, HBV core antigen expression was 99% was reduced in hepatocyres upon RNAi treatment [66].
Hepatitis C Virus (HCV) is a major cause of chronic liver disease. In HCV treatment, a set of siRNAs were used to reduce the destroying effects of HCV replication on the liver [67]. It was also reported that RNAi approach was specifically inhibited HCV RNA replication and protein expression in Huh-7 cells by using a selectable sub-genomic HCV replicon cell culture system [68]. In another study targeting the gene encoding Fas (death receptor) was achieved in order to protect mice from liver failure and fibrosis in two models of autoimmune hepatitis. In this experiment, intravenous injection of anti-Fas siRNA resulted with specifically reduced Fas protein levels in mice liver for duration of 10 days. So, Fas siRNA treatment demonstrated a hepatoprotective effect [68].
Other diseases
Alzheimer’s disease, Parkinson’s disease, Huntington’s disease and Amyotrophic Lateral Sclerosis (ALS) are of relatively common age related neurodegenerative disorders characterized by dysfunction and death of specific neuron populations. There are studies that cultured neurons are efficiently transfected with siRNAs; and as a result, target genes are silenced. For example, of the neurotrophin receptor p75 which caused neuronal apoptosis was functionally inhibited by siRNA application in cultured neurons [70]. In two separate studies, when pro-apoptotic members of bcl-2 gene family and caspases were silenced by siRNA treatments, neuronal cell death was prevented in Drosophila in order to define the cause of neurobiological diseases [71,72]. As for viral infections, Chikugunya is one of the most important arboviral infections with no antiviral or licensed vaccine in therapy use. Specifically designed anti-nsP3-siRNA and anti-E1-siRNA treatments had caused 99% inhibition; making RNAi as a promising therapeutic approach [73]. For cardiovascular diseases, a high level of cholesterol in blood is a major risk factor for atherosclerosis and heart disease. It has been reported that RNAi mediated silencing of PCSK9 gene has lowered LDL cholesterol in cynomolgus monkeys and rodents without affecting HDL or triglyceride levels; and, targeting of PCSK9 could be an alternative way to treat high cholesterol in human body [74].
RNA interference is a very promising tool in the field of medicine by having the ability of selective gene silencing and being frequently used for establishing gene function. The recent improvements made on siRNA applications and RNAi screening resulted in accelerated courage to use it in diverse area of medicine; especially for cancer therapy. The newly published studies triggered RNAi applications come of age are as follows: novel mechanisms clarified in signal transduction pathways [86], new insights into fundamental cell biological processes such as autophagy [87] and pluripotency [88]; identification of therapeutic targets for pathological processes like tumourigenesis [89], and resistance to cancer treatment [90]; defining hundreds of biological contact points between host organisms and pathogenic viruses [91]. Genome-scale RNAi screening established the ability to identify tumour vulnerabilities that are not oncogenic per se, resulted in illuminating a growing trend in cancer research: discovering and targeting Non-Oncogene Addiction (NOA) in tumours [92]. These NOAs are not genetically mutated and, thus, would not be detected in an audit of a cancer’s genome. Since cancer genome is unstable and the signalling pathways exhibit degenerated activation, drugs targeting a single oncogene (such as BCR-ABL in chronic myeloid leukemia) leads to adaptive tumour evolution and drug resistance [93]. To improve outcomes for patients and overcome cancer, development of “smart therapeutics” that stands for treatments which attack cancer in various ways and offer multiple drug targets has come into prominence: Oncolytic Viruses (OVs) belong to this class [91].
OVs are replicating therapeutics that are engineered to grow in tumor cells but are unable to productively infect normal tissues. OVs are more efficient than small molecules because, instead of trying to inhibit a single oncogenically activated pathway; OVs have the access upon whole malignant signaling networks which build up the cancer phenotype [94]. They are self-replicating biological weapons, that copy their genetic material, express virally encoded proteins and selfassemble, which in turn generates more therapeutic viral particles. Once the virus initiates infection within a tumor cell, it set up a series of capable viral proteins that invades the cell’s metabolic machinery. Administration of transcriptional and translational machinery by the virus means that, the infected cell will be unable to support its own basic functions and defeat; releasing OV particles that can move on to the next tumor cell. Another approach of OVs are not only directly killing or infecting tumor cells; but also use a second strayegy as killing the tumor bed, resulted in good therapeutic outcomes [95]. As for HIV from RNA viruses, a coctail of drugs inhibiting multiple virus proteins for efficient management of illness is needed for preventing virus from developing resistance. A major step came forward in 2007, when the FDA approved the chemokine (C–C motif) receptor 5 inhibitor; maraviroc, as the first-in-class drug targeting a host factor required for a pathogenic virus HIV–1, R5 strain [96]. Clinically used OVs such as vaccinia virus JX–594 (Jennerex Biotherapeutics, CA, USA) [97] and OncoVEX (Amgen, CA, USA) [98] are considered as oncovaccine and used in melanome treatment.
Recent developments in genome wide RNAi screening has developed an approach related with targetability of non-oncogene support pathways for cancer treatment, for outcoming the interactome between cancer cells and an oncolytic rhabdovirus Maraba. In this study, Mahoney et al. [99] identified a non-oncogene addiction to the ER stress response as a synergist target for oncolytic virus therapy by virus-triggered apoptosis of resistant cancers.
As for advances in genome scale RNAi screening technology, a number of technical innovations accelerated the phenomenon in three important ways. Initially, development of next-generation inducible shRNA vectors and advances in mouse shRNA transgenic technology will facilitate in vivo validation more applicable reflections in clinic. Secondarily, these these newly designed vector systems, coupled with recent advances in barcode deconvolution and bioinformatics, will enhance the utility of pooled shRNA screening in vivo. And finally, the development of a massively parallel sensor assay for unbiased, highthroughput, functional evaluation of shRNA sequences is a prelude to the production of next-generation genome-wide shRNA resources that will be more potent and less noisy than the existing libraries [100].
The RNAi biological process has emerged as a powerful tool for identification of gene function studies as well as well as a new promising therapeutic approach by a specific gene knockdown. Despite challenges such as off-target effects, cytotoxicity, need for efficient delivery methods; RNAi therapeutics appears to mark a new era for the therapy of commonly seen diseases. Despite the widespread interest in developing siRNA drugs, their clinical implementation still requires further advances in both delivery vehicles and siRNA activity itself. Most important challenges are non-specific gene silencing, activation of innate immune system, the lack of efficient in vivo delivery systems still remain to be handled for achieving success in pre-clinics. Besides, development of efficient tissue-specific and differentiationdependent expression of siRNA is essential for transgenic and therapeutic approaches. Despite these obstacles in this new field of gene therapy, there are successful in vitro and in vivo experiments for raising hopes in treating human disease with RNA interference. In the follow up clinical studies, rational design of siRNA delivery vehicles should be constructed for successful clinical outcomes of siRNA based therapeutics. By applying oncolytic viruses in RNAi application field, the safety and efficacy of the process is improved in clinical use. Recent advances in genome scale RNAi screening tools will potentially accelerate the validation of oncolytic viral therapy for clinical approval.