Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 1
Previous studies showed that chronic administration of the monoamine depleting agent reserpine in low doses promotes progressive cognitive and motor impairments in rats, and this protocol has been used as a pharmacological progressive model of Parkinson's disease. These behavioral alterations are accompanied by increased brain oxidative stress. We aimed to verify the effects of the concomitant treatment with the antioxidant agent alpha-tocopherol on the motor and cognitive deficits induced by chronic reserpine in rats. Rats were repeatedly treated with 0.1 mg/kg reserpine with or without a concomitant treatment with 40 mg/kg alpha-tocopherol. Across the treatment, motor and cognitive performances were evaluated by the catalepsy and novel object recognition tests, respectively. As expected, reserpinetreated rats showed progressively increased duration of catalepsy together with short-term memory deficits in the object recognition test. Importantly, these detrimental outcomes due to reserpine treatment were prevented by concomitant daily administration of the antioxidant agent alpha-tocopherol. The results show a preventive role of alpha-tocopherol on behavioral alterations induced by repeated reserpine treatment. This is relevant to the investigation of possible neuroprotective interventions in Parkinson’s disease.
<Keywords: Reserpine; Parkinson’s disease; α-tocopherol; Motor impairment; Short-term memory impairment
NOR: Novel Object Recognition; PD: Parkinson’s Disease; RES: Reserpine; ROS: Reactive Oxygen Species; TOC: Alpha- Tocopherol; VR: Vehicle For Reserpine; VT: Vehicle For Alpha- Tocopherol; PKC: Protein Kinase C
Reserpine precludes the storage of monoamines through the blockage of the synaptic vesicles transporters [1]. Consequently, synaptic vesicles are still available but there is a reduction in the amount of dopamine in the synaptic cleft. Because an important loss of dopaminergic neurons is the core feature of Parkinson´s disease (PD) [2], reserpine administration to rodents is a valid approach to study this disease in animal models [3-5]. The acute administration of a high dose of reserpine (above 1.0 mg/kg) leads to severe motor impairment [4]. In addition, acute injection of reserpine in lower doses causes memory deficits in the absence of motor damage [6,7]. However, although both cognitive and motor impairments are symptoms of PD, their emergence shortly after an acute injection is not compatible with the gradual progression of symptoms found in the clinical situation. More recently, studies have shown that the chronic administration of reserpine in low doses can promote progressive cognitive and motor impairments, along with decreased tyrosine hydroxylase levels in the nigrostriatal pathway [8]. This protocol is suggested as a progressive pharmacological model of PD [8,9].
Besides its classical mechanism of action (i.e. blockage of the vesicular transport of monoamines), there is clear evidence that reserpine also causes an increase in cellular oxidative stress, possibly potentiated by the rise in the levels of dopamine in the cytoplasm, which undergoes oxidative metabolism [10]. In this respect, the central nervous system is quite vulnerable to reactive oxygen species (ROS), which play a very important function in the pathogenesis of neurodegenerative disorders, including PD [11]. For example, there is evidence that the inclusion of antioxidant agents in the pharmacological treatment of PD has advantages over the treatment based only in dopamine replacement [11-13]. In addition, the repeated treatment with reserpine that induces progressive features compatible with PD also leads to increased brain oxidative stress [9]. However, it is unclear if a possible oxidative damage is responsible for the behavioral deficits presented by animals repeatedly treated with reserpine.
Antioxidant agents mainly act as a reinforcement of endogenous antioxidant defenses. An important antioxidant agent is vitamin E (alpha-tocopherol; TOC), which plays an essential role in protecting the body against the damaging effects of ROS. Specifically, TOC blocks the propagation step of lipid peroxidation of polyunsaturated fatty acids in membranes and lipoproteins [14], mainly by neutralizing the effects of peroxides and oxygen free radicals [15].
The aim of this study was to evaluate the effects of the antioxidant agent TOC on motor, cognitive and neuronal parameters in animals submitted to a progressive pharmacological animal model of PD, i.e., the repeated treatment with a low dose of reserpine.
Animals
We used 75 five-month-old male Wistar rats (300-500 g). The animals were obtained from the Physiology Department at the Federal University of Rio Grande do Norte, and were housed in groups of four, in plastic cages, under controlled conditions of ventilation, temperature (23 ± 1ºC), and light/dark cycle (12h/12h, lights on 6:30 a.m.), with free access to water and food. The rats were handled according to the Brazilian law for the use of animals in scientific research (Law Number 11.794) and all the procedures described were approved by the local ethical committee (CEUA/UFRN nº 051/2011).
Drugs
Reserpine (RES; Sigma Chemical Co., St. Louis, MO) was dissolved in acetic acid and further diluted in distilled water at the concentration of 0.1 mg/mL, pH ≈ 6.5. We used this vehicle (glacial acetic acid diluted in water) as a control for reserpine treatment (VR). RES and VR were given s.c. on alternate days. The antioxidant alpha-tocopherol (TOC; Sigma Chemical Co., St. Louis, MO) was diluted in distilled water with Tween-80 at the concentration of 40 mg/mL. We used the vehicle used to dilute TOC (VT) as a control for TOC treatment. These solutions were injected i.p. daily. The volume of injection was 1 mL/kg of body weight in all cases. We prepared all solutions every 48 hours and kept them at 4ºC between administrations.
Experimental design
The rats were randomly assigned to the following groups: VR + VT (n=18), RES + VT (n=19), RES + TOC (n=19) and VR + TOC (n=19). Drug treatment lasted 30 days. Animals received 15 s.c. injections of RES (0.1 mg/kg) or VR every 48 hours, concomitantly to daily i.p. administration of TOC (40 mg/kg) or VT.
Before the beginning of the experiments, all animals were submitted to a daily 5-minute handling session for five consecutive days. Throughout the treatment, all the animals were subjected to catalepsy tests (performed daily) and part of the animals (n=35, 7-11 per group) went through the novel object recognition (NOR) tasks (days 2, 12 and 18 of treatment). The experimental design is shown in Figure 1. Both behavioral tests were performed as described in our previous study [8] and were conducted before the injections of that day. Thus, all behavioral evaluations were performed 48h after the last injection of reserpine in order to avoid acute effects of the drug. NOR sessions were recorded with a digital camera fixed above the arena and the behavior was analyzed through video-tracking software (Anymaze, Stoelting Co, Wood Dale, Illinois, and USA). Before each experimental procedure, the apparatuses were cleaned with a 5% alcohol solution, and the experimental groups were alternated across testing.
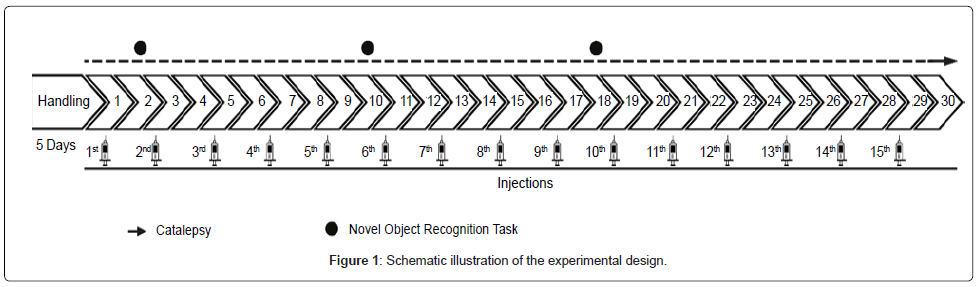
Figure 1: Schematic illustration of the experimental design.
Statistical analysis
We analyzed the performances in catalepsy test (total time spent in immobility until the animal removed both forepaws of the bar) by the two-way ANOVA with repeated measures followed by Tukey’s multiple comparison post hoc test. In the NOR task we conducted one-way ANOVA followed by Bonferroni’s multiple comparison post hoc test in order to compare old versus familiar object exploration. Analyses for the exploration ratio throughout test sessions and among experimental groups were conducted through two-way ANOVA followed by Tukey’s Post Hoc test.
Catalepsy
Figure 2 shows that from day 15 onwards there was an increase in catalepsy behavior of the group RES + VT compared to all other groups (RM two-way ANOVA; days of treatment [F(29,2130) = 16.72, P < 0.0001], treatment [F(3,2130) = 211.0, P < 0.0001] and days of treatment × treatment interaction effects [F(87,2130) = 4.876, P < 0.0001]). This increase was not detected for the group RES+TOC.
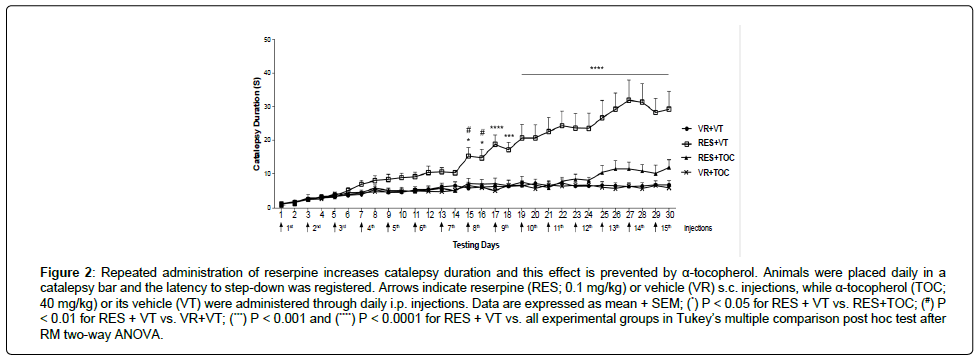
Figure 2: Repeated administration of reserpine increases catalepsy duration and this effect is prevented by α-tocopherol. Animals were placed daily in a catalepsy bar and the latency to step-down was registered. Arrows indicate reserpine (RES; 0.1 mg/kg) or vehicle (VR) s.c. injections, while α-tocopherol (TOC; 40 mg/kg) or its vehicle (VT) were administered through daily i.p. injections. Data are expressed as mean + SEM; (*) P < 0.05 for RES + VT vs. RES+TOC; (#) P < 0.01 for RES + VT vs. VR+VT; (***) P < 0.001 and (****) P < 0.0001 for RES + VT vs. all experimental groups in Tukey’s multiple comparison post hoc test after RM two-way ANOVA.
Novel object recognition
We found that all animals spent more time exploring the new object in the second day of protocol (first test; Figure 3A; one-way ANOVA [F(7,62) = 11.23; P < 0.0001]). Reserpine treatment impaired short-term memory after the 12th day of protocol (second and third tests). Conversely, treatment with α-tocopherol was able to prevent the short-term memory impairment (Figure 3B; one-way ANOVA [F(7,74) = 6.864; P < 0.0001] and Figure 3C; one-way ANOVA [F(7,68) = 10.00; P < 0.0001]). We also performed statistical analyses in order to evaluate the effect of drug administration in objects exploration ratio throughout test sessions and among experimental groups. We found that in the third test session animals’ receiving RES differs on exploration rate of new (Table 1; two-way ANOVA [F(6,89) = 2.843; P < 0.05]) and old objects (Table 1; two-way ANOVA [F(6,89) = 2.843; P < 0.05]) when comparing to both VR + VT and RES + TOC. Yet, we found that only RES + VT group presented alterations in object discrimination across tests. More accurately, exploration of old and new objects increased and decreased, respectively, comparing first and second tests (Table 1; two-way ANOVA [F(3,89) = 2.760; P < 0.05]) and first and third tests (Table 1; two-way ANOVA [F(3,89) = 2.649; P < 0.05]).
| Groups | |||||
|---|---|---|---|---|---|
| Tests | Objects | VR + VT | RES + VT | RES + TOC | TOC |
| First Test Post 1st injection |
Old | 36.94 ± 6.61 | 26.06 ± 5.01 | 34.11 ± 5.84 | 40.6 ± 4.91 |
| New | 63.06 ± 6.61 | 73.94 ± 5.01 | 65.89 ± 5.84 | 59.4 ± 4.91 | |
| Second Test Post 6th injection |
Old | 34.69 ± 5.92 | 43.97 ± 5.31¥ | 26.85 ± 5.87 | 39.17 ± 6.32 |
| New | 65.31 ± 5.92 | 56.03 ± 5.31¥ | 73.15 ± 5.87 | 60.83 ± 6.32 | |
| Third Test Post 9th injection |
Old | 27.71 ± 4.97 | 59.99 ± 6.95€ # | 31.99 ± 6.43* | 37.07 ± 3.25 |
| New | 72.28 ± 4.97 | 40.01 ± 6.95€ # | 68.01 ± 6.43* | 62.93 ± 3.25 | |
Table 1: Exploration rate in the NOR task throughout the test sessions. Data are expressed as mean ± SEM. (*) P < 0.05 and (€) P < 0.01 when comparing RES + VT vs. RES + TOC and VR + VT vs. RES + VT respectively. (¥) P < 0.05 and (#) P < 0.001 when comparing the first vs. second test and first vs. third test respectively. All statistical analyses were conducted through two-way ANOVA followed by Tukey’s Post Hoc test.
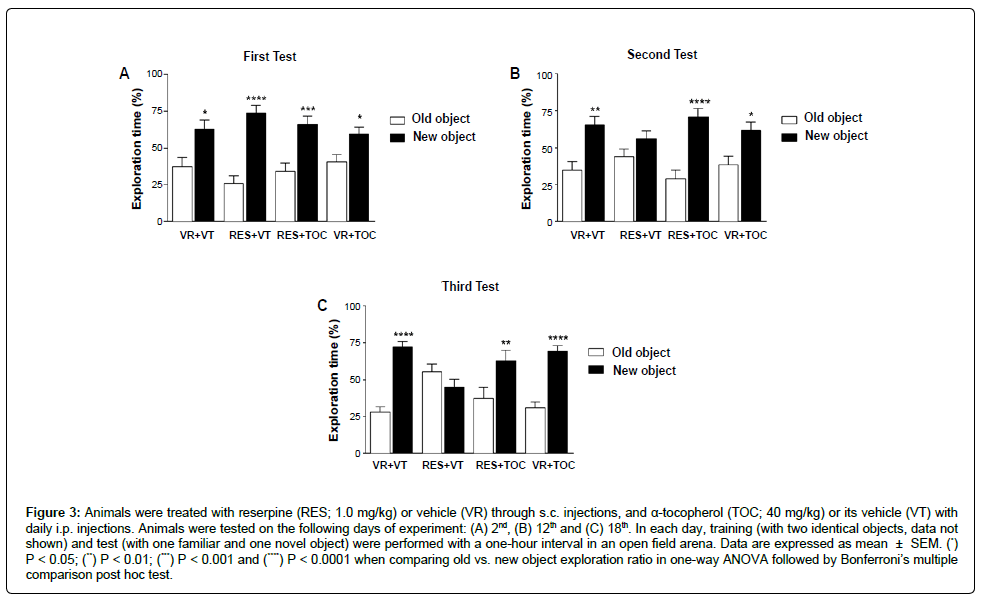
Figure 3: Repeated administration of reserpine increases catalepsy duration and this effect is prevented by α-tocopherol. Animals were placed daily in a catalepsy bar and the latency to step-down was registered. Arrows indicate reserpine (RES; 0.1 mg/kg) or vehicle (VR) s.c. injections, while α-tocopherol (TOC; 40 mg/kg) or its vehicle (VT) were administered through daily i.p. injections. Data are expressed as mean + SEM; (*) P < 0.05 for RES + VT vs. RES+TOC; (#) P < 0.01 for RES + VT vs. VR+VT; (***) P < 0.001 and (****) P < 0.0001 for RES + VT vs. all experimental groups in Tukey’s multiple comparison post hoc test after RM two-way ANOVA.
In this study, we investigated the effects of concomitant treatment with TOC on catalepsy behavior and NOR task in rats submitted to a chronic treatment with a low dosage of reserpine. We observed that the motor and cognitive impairments induced by chronic treatment with reserpine were prevented by treatment with TOC. These results can be seen in the evaluation of catalepsy behavior performed 48 h after each reserpine injection (Figure 2) as well as in the analysis of exploration time in the novel object recognition task (Figure 3 and Table 1).
As previously observed in studies by our group [8,9], repeated treatment with a low dose (0.1 mg/kg) of reserpine in rats induced the progressive appearance of motor impairment. This impairment is marked by a gradual increase in the duration of catalepsy behavior. Indeed, as one can see in Figure 2, reserpine-treated (RES + VT) animals start differing from control subjects after 7 reserpine s.c. injections. It is well documented that catalepsy in rodents indicates akinesia and rigidity that are important symptoms of PD [16-18]. Importantly, we did not observe this impairment in the group that was concomitantly treated with TOC. Indeed, the group RES + TOC (Figure 2) presented catalepsy duration similar to control across the treatment.
Besides motor assessment, the protocol used in the present study includes the cognitive evaluation. Cognitive deficits have been reported as symptoms of PD, and can even appear before the motor deficits. In a previous study, we have shown that the protocol of reserpine treatment used here induces short-term memory deficits before the appearance of increased catalepsy behavior and other motor signs [8]. The present study corroborates those findings. We used the NOR task, which involves recognition memory and executive functions, both functions that can be impaired in PD [19,20]. Our results corroborated the previous study showing that animals treated with reserpine failed to discriminate the objects in the test session (in the second and third tests, Figure 3). Further, similarly to that described for motor evaluations, the deficit was prevented by TOC administration. Indeed, animals treated with both reserpine and TOC presented increased novel object exploration in all tests, similarly to control subjects. In addition, comparisons among experimental groups showed that animals treated with RES had worse object discrimination compared to both control and RES + TOC groups in the third test. Finally, when performances across the three tests were analyzed, only the group treated with reserpine alone presented discrimination deficits in the second and third tests compared to the first test (Table 1). These additional analyses reinforce the prevention of the reserpine-induced object recognition impairment by co-treatment with TOC.
As mentioned, reserpine is a non-selective inhibitor of the vesicular monoamine transporter [1]. Thus, one could raise the possibility that the behavioral alterations induced by reserpine treatment are related exclusively to the dopamine depletion caused by this blockage. In other words, the alterations could be a consequence of an additive effect on dopaminergic function. However, there is evidence that favors the hypothesis that the progressive effect of the repeated treatment with reserpine is due to oxidative damage. First, a previous study has shown that the classical acute treatment (with a dose 10 times higher than the one we used) did not cause a reduction in tyrosine hydroxylase staining (an indicative of dopaminergic neuronal function), although causing an important motor impairment [21]. Conversely, the protocol used here (repeated treatment with a low dose) reduced tyrosine hydroxylase staining in the substantia nigra and striatum, and part of the alterations induced by the treatment were not recovered after 30 days of treatment withdrawal [8]. Second, it has been shown that reserpine treatment increases brain oxidative stress and this alteration is accompanied by behavioral deficits [10,22,23]. In addition, in a previous study [9] the repeated treatment with a low dose of reserpine induced an increase in striatal level of lipid peroxidation, which occurred concomitantly to the motor impairment. These results lead us to question if cotreatment with TOC would prevent the progressive motor and cognitive alterations induced by the repeated treatment with a low dose of reserpine. As discussed above, treatment with TOC was able to prevent these deficits. This preventive effect might be explained by a neuroprotection mechanism, probably by a reduction the in neurotoxic dopamine oxidation bioproducts [24].
Despite the well-known antioxidant properties of vitamin E, it is important to mention that tocopherol and other antioxidant agents can have pro-oxidant effects as well. Indeed, the ability of these compounds to accept and donate electrons enables them to cause oxidative damage under certain conditions [25]. However, this prooxidant action is mainly found in vitro, and under high concentrations [26,27]. Some in vivo studies have also shown pro-oxidant effects of classical antioxidants, but they are variable depending on substance, concentration, age of the subject and target molecules [25,28-30]. Further, it seems that their preferential action is antioxidant when an oxidant insult from another source is present [31]. In the case of the present results, there was no evidence of a pro-oxidant action regarding possible behavioral alterations.
Nevertheless, an antioxidant role of vitamin E in ameliorating neurodegeneration in PD has been consistently proposed by in vitro and animal studies [32-37]. On the other hand, despite strong evidence favoring an antioxidant effect, the exact mechanism of action of vitamin E in Parkinson´s disease is still under investigation [32]. There is evidence that vitamin E, particularly alpha-tocopherol, can act through other mechanisms not related to modulation of oxidative stress. For example, studies showed that alpha-tocopherol regulates the expression of several genes [38,39] and inhibits protein kinase C (PKC) activity [40,41]. The later could be related to the neuroprotective action of this compound, because PKC activation has been implicated in cell death signaling pathways related to PD [42]. This relationship was found in studies with animal models of PD induced by the toxins 1-methyl-4-phenylpyridinium [43] and paraquat [44]. If PKC activation is also relevant for reserpine-induced Parkinsonism it is still unknown.
Regardless of the specific mechanism related to the prevention of behavioral alterations found in the present study, there is evidence that increased oxidative stress underlies the physiopathology of neurodegenerative diseases such as PD [45-48]. Further, clinical data suggest that neuroprotective treatments based on increasing antioxidant defenses are able to delay the progression of the pathology [49-56]. Thus, a neuroprotective intervention could be a relevant line of investigation in animal models of this disease. However, the usual acute pharmacological models include severe motor impairment upon a single injection of reserpine or specific neurotoxins [4,57-60]. This approach is not suitable for the investigation for testing neuroprotective interventions because they usually present a preventive and/or a neurodegeneration delaying profile. Further, most of the previous studies investigating the effects of vitamin E treatments on PD models did not investigate progressive behavioral deficits related to the clinical symptoms of the disease [33,35-37]. In this sense, the need for animal models of PD more compatible with clinical outcomes when investigating neuroprotective therapies has been pointed out. Thus, the present findings reinforce the idea that the protocol of progressive Parkinsonism induction with reserpine is suitable for investigating possible neuroprotective interventions in animal models of PD.
In conclusion, concomitant treatment with alpha-tocopherol prevents behavioral alterations induced by repeated reserpine. Although the antioxidant action of vitamin E is probably related, the exact mechanism underlying this preventive effect remains to be investigated. Finally, the progressive behavioral motor and cognitive alterations induced by repeated reserpine treatment seems an adequate protocol to investigate possible neuroprotective interventions for PD.
The authors would like to thank Antonio Carlos Queiroz de Aquino for capable technical assistance. This study was supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil), Fundação de Amparo a Pesquisa do Estado do Rio Grande do Norte (FAPERN, Brazil), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil).