Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2014) Volume 5, Issue 3
The genetic variation of wild and hatchery populations of swimming crab Portunus trituberculatus based on observation of amplified fragment length polymorphism (AFLP) was described. A group of 180 genotypes belonging to five wild samples, Dongying (DY), Weifang (WF), Weihai (WH), Qingdao (QD), Rizhao (RZ) and one hatchery population, Yantai (YT) were screened using eight different AFLP primer combinations. A total of 396 loci were screened in the six studied populations. 49.9%, 48.5%, 52.3%, 51.2%, 50.3% and 44.5% of these loci were polymorphic among the individuals tested in the DY, WF, WH, QD, RZ and YT populations, respectively. The number of polymorphic loci detected by single primer combinations ranged from 22 to 37. The average heterozygosity of the DY, WF, WH, QD, RZ and YT populations were 0.087, 0.085, 0.096, 0.092, 0.090 and 0.068, respectively. The WH population showed the highest genetic diversity in terms of total number of AFLP bands, total number of polymorphic bands, average heterozygosity and percentage of low frequency (0-0.2) polymorphic loci among all the populations, while the WF population was the lowest among the wild populations. Compared with the wild populations, the hatchery population showed a low genetic viability.
Keywords: Swimming crab, Portunus trituberculatus , AFLP marker, Genetic variability
The swimming crab Portunus trituberculatus, is distributed mainly on sandy and muddy bottoms in the coastal waters of Japan, Korea, and China. It is one of the most common edible crabs in China and Korea and supports a large crab fishery and aquaculture in China [1]. It is now being cultured in North China, especially in Shandong Peninsula, because of its high commercial interest. A few reports are available on molecular phylogeny and population structure in this species using different molecular marker techniques [1-4]. Long-term conservation of genetic diversity is important for any species [5]. Swimming crab resource management and enhancement are a recent practice to maintain long-term resource sustainability. A basic understanding of stock structure among geographical swimming crab samples is thus required.
In general, the effective sizes of founder populations are restrained by farming conditions, in which only a few individuals as broodstock are used. This practice may lead to the erosion of genetic diversity of stocks, thereby compromising industrial performance. Intentional and accidental release of cultured swimming crabs into wild environment could have major ecological consequences. If a large number of cultured swimming crabs escape or are released from aquaculture facilities, they could significantly alter the genetic composition of wild populations by either displacing them or interbreeding with them [6]. Most hatchery stocks typically show a reduced genetic variability, which may possibly result in the reduction of the population’s capability to adapt to new environments [7]. Therefore, it is important to establish baseline information on genetic background of the aquaculture population both for genetic enhancement programs as well as protection of the genetic integrity of natural populations. Unfortunately, to date little is known about the population structure of swimming crab in China. Molecular markers provide useful tool for the assessment of genetic variations. Among the several marker systems, amplified fragment length polymorphism (AFLP) is highly reliable for the assessment of genetic variation among and within populations [8-10]. AFLP does not require previously known genetic information, a feature especially useful with species for which there are no established polymorphic markers, or for which there is limited sequence information [11,12]. AFLP is a PCR-based, multi-locus fingerprinting technique that has tremendous power for revealing polymorphism [13].
The aim of this study is to assess the genetic diversity of among samples of swimming crab from Shandong peninsula in China, and that genetic differences were observed among five wild and one cultured samples of swimming crab.
Swimming crab sampling
A total of 180 individuals of swimming crab specimens, based on six sample sets, 30 individuals each, were collected in 2009 and genetically screened in the present study. The Weights of the samples are from 80 to 120 g. Geographic locations, sample sizes are given in Figure 1. Wild swimming crab were collected at five sites, the coast of Dongying (DY), the coast of Weifang (WF), the coast of Weihai (WH), the coast of Qingdao (QD) and the coast of Rizhao (RZ). Hatchery swimming crabs were from a hatchery station in Yantai (YT). The hatchery sample was founded using wild caught individuals from Bohai sea. Samples were stored frozen (-20ºC) until genetic analysis was performed.
Figure 1: Sampling localities of swimming crab Portunus trituberculatus samples. DY, Dongying (n=30); WF, Weifang (n=30); YT, Yantai (n=30); WH, Weihai (n=30); QD, Qingdao (n=30); RZ, Rizhao (n=30).
Genomic DNA extraction
Genomic DNA was extracted from swimming crab muscle. About 150 mg muscle tissue was digested overnight at 37ºC in 0.85 ml of lysis buffer (6 M urea, 10 mM Tris–HCl, 125 mM NaCl, 1% SDS, 10mM EDTA, pH 7.5) and 50µl of proteinase K (20 mg/ml). DNA was extracted twice with phenol and once with chloroform. DNA was precipitated by adding 200µl of 7.5 M ammonium acetate and 500µl of ethanol. DNA was collected by brief centrifugation and washed twice with 75% ethanol, air-dried, and dissolved in TE buffer.
AFLP reactions
Procedures of AFLP analysis were essentially based on Vos et al. [13]. Genomic DNA was processed using the ‘‘AFLP Analysis System I’’ (Invitrogen Corp.) according to the manufacturer’s instructions with some modifications. The EcoR I primers used were not radioactively labeled. Instead, a modified silver staining method detailed below was used. After partial digestion and ligation to adaptors, samples were pre-amplified and amplified through two sequential steps. PCR pre-amplification, with primers carrying one selective nucleotide, was performed with 30 cycles at 94º C for 30s, 56º C for 1 min and 72º C for 1 min. Samples were processed immediately for the following step or stored at -20º C. After dilution up to 20 times in water, PCR products from the pre-amplification reaction were used as templates for selective amplification. The amplification step consisted of 12 cycles at 94º C for 30 s, 65º C for 30 s (with a decreasing ramp of 0.7º C each cycle) and 72º C for 1 min, followed by 30 cycles at 94º C for 30 s, 56º C for 30 s and 72º C for 1 min. Altogether eight primer combinations that produced clear and reproducible fragments were selected for further analysis (Table 1).
| Primer combination | EcoRI primers | MseI primers |
|---|---|---|
| P1 | EcoR I primer + ACG | Mse I primer + CTG |
| P2 | EcoR I primer + ACT | Mse I primer + CTC |
| P3 | EcoR I primer + AGC | Mse I primer + CTT |
| P4 | EcoR I primer + AAC | Mse I primer + CTC |
| P5 | EcoR I primer + AGC | Mse I primer + CTA |
| P6 | EcoR I primer + AAC | Mse I primer + CAT |
| P7 | EcoR I primer + AAC | Mse I primer + CAG |
| P8 | EcoR I primer + AGG | Mse I primer + CTG |
Table 1: AFLP primer combinations used in the study. The sequences for the EcoRI primer were 5’-GACTGCGTACCAATTC-3’, and for MseI primer were 5’-GATGAGTCCTGAGTAA-3’.
Gel electrophoresis and silver staining
The PCR products were mixed with an equal volume of formamide dye (99% formamide, 10 mM EDTA, 0.05% bromophenol blue and 0.05% xylene cyanol). The samples were heated to 95º C to denature for 5 min and immediately placed on ice. The gel was pre-electrophoresed at 60 W for 30 min, then 5.0 µl of the amplified DNA was loaded and run through a 5% denaturing polyacrylamide gel (4.75% acrylamide, 0.25% bisacrylamide, 7.5 M urea and 1×TBE buffer) with 1×TBE buffer on a DNA sequencing system (Liuyi Corporation, China) at 80 W for 120 min.
Silver staining was conducted using the procedures of Merril and Liu with modifications [14,15]. After electrophoresis, the gel was fixed in 10% ethanoic acid for at least 30 min. The gel was rinsed in distilled water three times and stained with a mixture of 0.1% silver nitrate and 0.15% formaldehyde for 30 min. The stained gel was rinsed again with distilled water and immersed in a developing solution (3% sodium carbonate, 0.15% formaldehyde, 0.02% sodium thiosulphate). The development was subsequently stopped with 10% ethanoic acid when the bands became visualized and reached desirable intensity. Band sizes were estimated by a standard AFLP DNA ladder (Takara Corporation, China).
AFLP reproducibility test
The reproducibility of AFLP fingerprints was evaluated to increase the consistency of the results by comparing the fingerprints of the same swimming crab individual. Two selected individuals were independently processed from the beginning of the AFLP analysis for three replicates with two different primer combinations (No. 1 and 2 from Table 1). Reproducibility was calculated as the percentage of bands that showed consistent results over the three replicates analyzed per swimming crab sample.
Data analysis
AFLP bands were scored for presence (1) or absence (0) using the Crosscheck freeware 8 [16], and transformed into a 0/1 binary character matrix. Fragments that could not be scored unambiguously were not included in the analysis. The data matrix was analyzed for population genetic diversity using POPGENE software package 1.3.1[17]. Population genetic relationships were estimated by constructing a UPGMA tree based on Nei’s standard genetic distance [18]. Analysis of Molecular Variance (AMOVA) was performed to analyze genetic distance among samples using ARLEQUIN 3.1 [19]. Average heterozygosities and percent polymorphic loci were estimated using the TFPGA program 1.3 [20]. Average heterozygosity estimates were calculated for each locus and then averaged over loci according to Nei’s unbiased heterozygosity formula [18]. The percentages of polymorphic loci were estimated based on the percent of loci not fixed for one allele. Confidence intervals were generated by bootstrapping analysis at the 99% confidence level with 1000 replications. All the above significances were tested using t-test (P<0.05 and P<0.01). Estimation of pairwise Fst values for all sample combinations were also performed using ARLEQUIN program and were evaluated by a test analyogous to the Fisher’s exact test using the Markov-Chain method. Significance value was adjusted for multiple comparisons using the sequential Bonferroni correction [21].
AFLP polymorphism of six populations of swimming crab
Three-time repeated analysis of two randomly selected DNAs with primer combinations 1 and 2 showed highly reproducible results (reproducibility score = 99%), although some quantitative variances of the given amplicons were observed (data not shown). AFLP analysis of 180 swimming crab individuals using eight primer combinations produced a total of 396 scoreable bands, of which 49.9%, 48.5%, 52.3%, 51.2%, 50.3% and 44.5% were polymorphic over all the individuals tested in the Dongying, Weifang, Weihai, Qingdao, Rizhao and Yantai populations, respectively (Table 2). The total number of polymorphic bands over all populations varied from 22 (for primer combination 6) to 37 (for primer combination 3) per primer combination. The average heterozygosity of the DY, WF, WH, QD, RZ and YT populations were 0.087, 0.085, 0.096, 0.092, 0.090 and 0.068, respectively.
| Population | Number of polymorphic loci | N | H | P | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | Total | ||||
| DY | 18 | 22 | 34 | 23 | 23 | 19 | 25 | 23 | 187 | 375 | 0.09 | 49.9 |
| WF | 20 | 21 | 33 | 21 | 24 | 17 | 23 | 22 | 181 | 373 | 0.09 | 48.5 |
| WH | 23 | 24 | 37 | 23 | 26 | 22 | 27 | 25 | 207 | 396 | 0.1 | 52.3 |
| QD | 22 | 26 | 34 | 21 | 23 | 21 | 25 | 24 | 196 | 383 | 0.09 | 51.2 |
| RZ | 19 | 25 | 35 | 20 | 24 | 18 | 26 | 23 | 190 | 378 | 0.09 | 50.3 |
| YT | 17 | 18 | 30 | 19 | 22 | 16 | 21 | 20 | 163 | 366 | 0.07 | 44.5 |
Table 2: A summary of AFLP analysis of genetic variations in swimming crab using the eight primer combinations as listed in Table 1. P1 through P8 indicate primer combinations 1 to 8 as listed in Table 1. Total number of AFLP bands (N), average heterozygosity (H), percentage of polymorphic loci (P), Dongying (DY), Weifang (WF), Yantai (YT), Weihai (WH), Qingdao (QD) and Rizhao (RZ) are given.
Population structure and genetic differences between wild and hatchery populations
The WH population showed the largest number of total AFLP bands, total polymorphic bands and average heterozygosity among all the populations, while the YT population had the smallest number. The WF population displayed the smallest number of total AFLP bands, total polymorphic bands and average heterozygosity in wild populations. A greater number of total AFLP bands were observed from the wild populations than from the hatchery population. There were 375, 373, 396, 383 and 378 AFLP loci detected in the DY, WF, WH, QD and RZ populations, while it was 366 in the YT population, respectively (Table 2). The total number of polymorphic bands was also higher (P<0.05) in the wild populations than in the hatchery population. The total polymorphic loci were 187 in the DY population, 181 in the WF population, 207 in the WH population, 196 in the QD population, 190 in the RZ population and 163 in the YT population. However, no significant difference was found in the proportion of polymorphic bands among the five populations.
Differences in the percentage of polymorphic loci were found among populations as shown in Figure 2. The WH population showed the highest percentage of low frequency (0-0.2) polymorphic bands among all the studied populations, while the YT population had the lowest one. Among the wild populations, the WF population displayed the smallest percentage of low frequency polymorphic bands. In total, the percentage of low frequency polymorphic bands in the hatchery population was lower (P<0.05) than that in the wild populations, while the percentage of high frequency (0.6-1.0) polymorphic bands had a tendency to be higher in the hatchery population as compared with the wild populations.
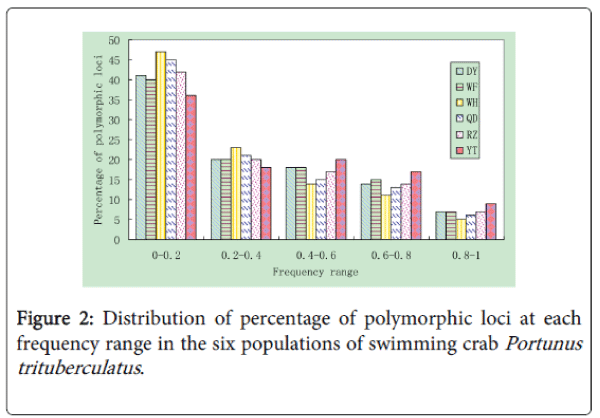
Figure 2: Distribution of percentage of polymorphic loci at each frequency range in the six populations of swimming crab Portunus trituberculatus .
The UPGMA dendrogram constructed on the basis of the inter sample genetic similarity is shown in Figure 3. Genetic distances between samples are summarized in Table 3. Significant genetic differentiation was detected using AMOVA among samples (P<0.05). The pairwise Fst values in Table 3 also indicated significant differentiation among the six samples except between DY and WF, WF and YT, WH and YT, and QD and RZ. These results suggested that genetic divergence between these samples may have arisen.
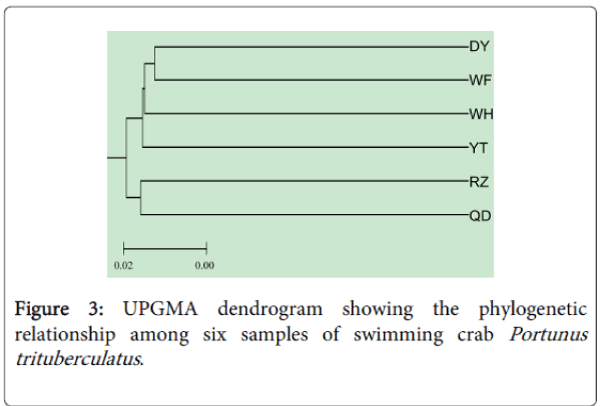
Figure 3: UPGMA dendrogram showing the phylogenetic relationship among six samples of swimming crab Portunus trituberculatus.
| Pop ID | DY | WF | WH | QD | RZ | YT |
|---|---|---|---|---|---|---|
| DY | *** | 0.017 | 0.022* | 0.038* | 0.037* | 0.029* |
| WF | 0.14 | *** | 0.021* | 0.036* | 0.033* | 0.019 |
| WH | 0.146 | 0.144 | *** | 0.031* | 0.029* | 0.018 |
| QD | 0.158 | 0.156 | 0.15 | *** | 0.015 | 0.028* |
| RZ | 0.151 | 0.16 | 0.152 | 0.147 | *** | 0.028* |
| YT | 0.149 | 0.142 | 0.147 | 0.15 | 0.153 | *** |
Table 3: Genetic distance and Fst values for pairwise comparison among different samples of Portunus trituberculatus . Genetic distance and Fst values are below and above the diagonal respectively. Dongying (DY), Weifang (WF), Yantai (YT), Weihai (WH), Qingdao (QD) and Rizhao (RZ) are given. Asterisk indicates significant genetic differentiation test by Fisher’s technique after a sequential Bonferroni correction (P<0.0125).
The WH population showed the highest diversity among all the populations while the YT population was the lowest. The WF population displayed the lowest diversity among wild populations. This phenomenon could be explained that the WF population lives in the Bohai sea, a semi-closed water. QD and RZ population are near geographically and have more chance for gene flow. In the dendrogram, the QD and RZ populations clustered together and were clearly separated from the DY, WF, WH and YT populations. The UPGMA dendrogram showed that the WH population clustered closer to the WF population than to the QD population. The reason is presumably due to the absence of physical barriers to migration between the WH and QD populations. Some differences in the percentage of polymorphic loci among populations were also found in the present study. The question is whether the polymorphic loci distribution maintained in each population temporally is stable or not. The present study did not address this question, and the only way to improve the robustness of the data presented here is repetitive sampling taking into consideration sample size, sampling time and the life stage of sample. Beside such molecular approaches, accumulation of biological and ecological data is crucial. This is because high levels of migration among populations would provide sufficient gene flow to prevent remarkable genetic population differentiation.
Genetic variability is an important attribute of the species under domestication, since those with higher levels of variation are most likely to present high additive genetic variance for productive traits. Wild populations represent the primary source of genetic variability for aquacultured stocks. Genetic variability of the hatchery strain seems likely to have been substantially reduced. This would be caused by losses of many low-frequency alleles due most likely to the small effective number of parents when the strain was founded, suggesting that the hatchery strain was bottlenecked. The YT population was founded using about 36 individuals, but the number of effective parents may be smaller than that. This suggests that the genetic diversity in the wild swimming crab is still not being fully exploited. In view of this, there will be a higher chance of enhance diversity viability by frequent outcrossing of the cultivated swimming crab population with the wild population rather than by selective breeding followed by inbreeding among cultivated varieties. In this way, the genetic variability among the cultivated varieties would also be enhanced, thus preventing the occurrence of further different inbreeding that will probably take place if the present situation of unplanned breeding persists. Therefore, for high sustainability of the culture of swimming crab, proper breeding programs must be implemented with careful management and monitoring such that there is frequent outcrossing with the wild forms as well as maintenance of any newly emerged traits by inbreeding.
Many cultured aquatic stocks represent genetically exogenous populations, thus, the intra-specific hybridization with wild stocks may result in reduction of fitness in wild populations [22-24]. Even cultured populations that originated from the same local population may threaten the fitness of the local population through the reduction of its effective population size [25], especially when the absolute size of the wild population is small [26]. Hence, caution should be exercised to avoid significant release of hatchery stocks of swimming crab into the wild, either intentionally, or accidentally.
This work was supported by a grant from Yantai University (SM13B22).