Andrology-Open Access
Open Access
ISSN: 2167-0250
ISSN: 2167-0250
Research Article - (2015) Volume 4, Issue 1
This review summarizes some information about the different ways in relation to sperm capacitation. On one hand, the classical pathway that define the functional changes that occur in sperm during in vitro capacitation with special emphasis on the factors that lead to the tyrosine Phosphorylation (PY), and on the other hand, molecules and process that are involved in new mechanisms involved in this event like reactive species, especially Nitric Oxide (NO) and protein nitrosylation.
<Keywords: Sperm capacitation; nitric oxide; protein nitrosylation; phosphorylation.
After mating or artificial insemination, millions of sperm are deposited in the female genital tract, of which only a small proportion is able to reach the caudal portion of the isthmus (Figure 1A). This sperm population encounters a sticky secretion of glycoprotein that modifies the sperm surface [1]. Motility decreases in this viscous medium and facilitates the sperm adhesion to the epithelium (Figure 1B). Sperm remains in the caudal portion of the oviductal isthmus, during the pre and peri-ovulatory time, forming the Sperm Reservoir (SR) [2,3]. This binding is a reversible process and the oviductal microenvironment signals stimulate sequential release of a limited number of sperm from the SR to the ampullary-isthmic junction. This ensures fertilization of oocytes in a time interval (Figure 1C), even if ovulation occurs over a long period of time [4]. However, the mechanisms which sperm are released from SR are unknown.
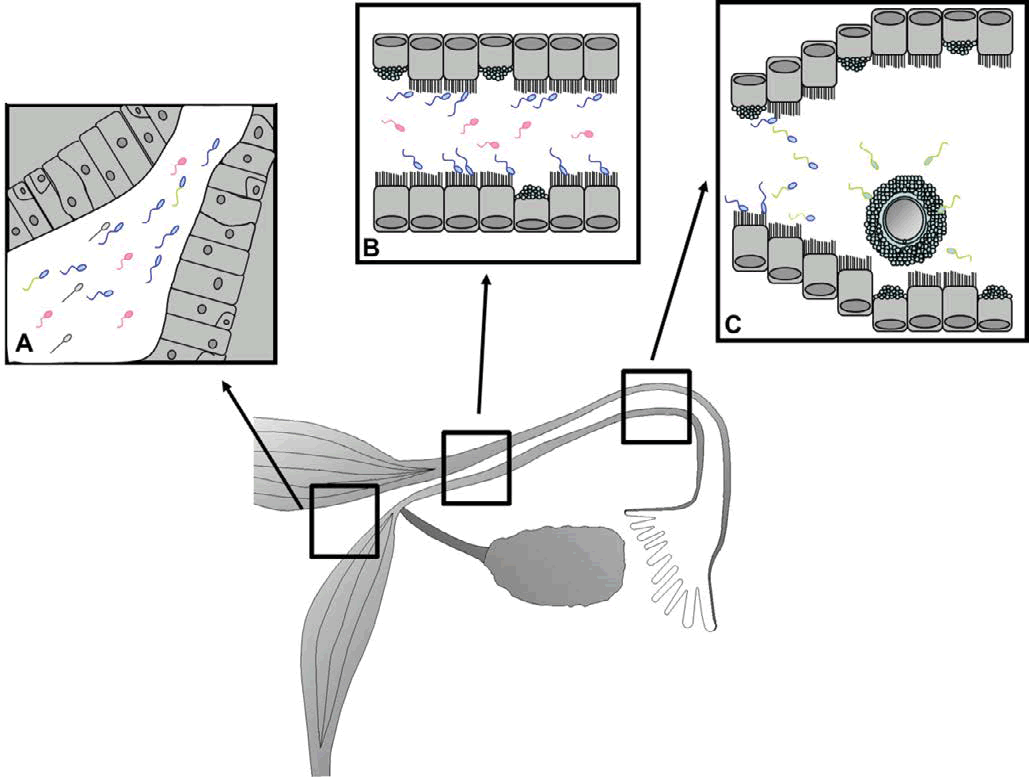
Figure 1: Capacitation process. A) After ejaculation, a heterogeneous population of sperm reaches the female reproductive tract. B) Only a few sperm achieves the oviduct and forms the Sperm Reservoir (SR) in the caudal portion of the isthmus. C) During peri-ovulatory time, sperm release from the SR and those who complete a correct sperm capacitation are able to contact with the oocyte and fertilize it. Different colours indicate distinct types of sperm: dead (grey), damaged (red), normal (blue), hyperactivated (green-blue) and successfully capacitated (green).
During the passage of sperm through the female genital tract, the spermatozoa undergo functional and molecular changes which confer ability to fertilize the oocyte (Figure 1). This process is known as sperm capacitation [5].
Capacitation is a complex process, which appears to be controlled by crosstalk between different pathways [6,7]. The most notable event is an increase in protein tyrosine phosphorylation [8,9] although an oxidative process also has been shown, including the nitric oxidedependent pathway [10,11]. In this sense, some papers have shown that NOS (Nitric Oxide Synthase) is present in the oviduct [12-14], oocyte, and cumulus and corona cells [15,16] of different species [12,17,18]. NOS isoforms are hormonally regulated in the oviduct and expresses differently throughout the oestrous cycle. In the oviduct, Nitric Oxide (NO) has been shown to regulate contractility [19], ciliary beating of the ciliated epithelial cells, the sperm motility or even inducing chemotaxis [20]. For .this reason NO also module sperm capacitation although the pathway is not known totally.
During the capacitation process, spermatozoa undergo a series of functional changes, which enables them to bind to the extracellular matrix of the oocyte and consequently require the acrosome reaction. Although the latter is under discussion as recently shown by Jin et al. [21] that the acrosome reaction in mouse sperm occurs before binding to the zona pellucida. Besides, the pattern of movement of sperm flagellum changes allowing penetration of the zona pellucida [22].
Capacitation process implied several changes sequentially. Some of these changes are rapid and occur at the moment of ejaculation. Others require a longer period of time in the female genital tract (in vivo) or in a medium capable of supporting this process (in vitro). All these processes (both rapid and slow), appear to be regulated by protein kinase A (PKA) and HCO-3, Soluble Adenylate Cyclase (SACY or sAC), and Cyclic Adenosine 3’5 ‘Monophosphate (cAMP) participate in this process (revised by [23]).
Traditionally, Reactive Oxygen Species (ROS) are considered to be injurious by products of cellular metabolism but also fundamentally participants in cell signalling and regulation mechanisms [24]. This apparent paradox also is true for spermatozoa, which are particularly susceptible to ROS-induced damage because their plasma membranes contain relatively large amounts of polyunsaturated fatty acids and their cytoplasm contains relatively low concentrations of scavenging enzymes [25], but require low concentration of ROS to acquire the fertilizing ability [6,26]. The essential role of ROS as modulators of capacitation is recognized in human [27], bovine [28], and mouse [27], and boar spermatozoa [29,30].
Facts during fast sperm capacitation
An early event during capacitation is the activation of sperm motility. Although sperm stored in the cauda epididymis being practically immobile consume oxygen in large proportions. The flagellum movement starts immediately after sperm are released from the epididymis and contact has been made with seminal plasma. This is due to exposure of sperm to the HCO-3 [31].
Facts during slow sperm capacitation
In contrast to the rapid activation of motility, other processes associated with capacitation require a longer period of time. During slow capacitation, sperm acquire the ability to fertilize, which is preceded by the preparation of the sperm to undergo the acrosome reaction and change the pattern of motility called hyperactivation. Components in oviductal fluid such as high weight molecular proteins and high density lipoproteins promote cholesterol efflux resulting in an increased capacitation and tyrosine Phosphorylation (PY) using the cAMP signalling pathway/PKA [32]. Additionally, these slow processes also are achievable in vitro by incubation of spermatozoa in defined media, which contain a protein source (usually bovine serum albumin (BSA)), and different ions, including HCO-3and Ca2+.
As we mentioned above, bicarbonate, calcium and cholesterol acceptor are essential during capacitation process entirely (Figure 2). These substances induce modifications lipid membrane, loss of cholesterol, activation of cAMP/PKA pathway, increase Ca2+ uptake and pH (pHi), hyperpolarisation of membrane potential, and PY [33]. However, there are other pathways in relation to capacitation as NO/ sGC/cGMP or protein nitrosylation are being studied.
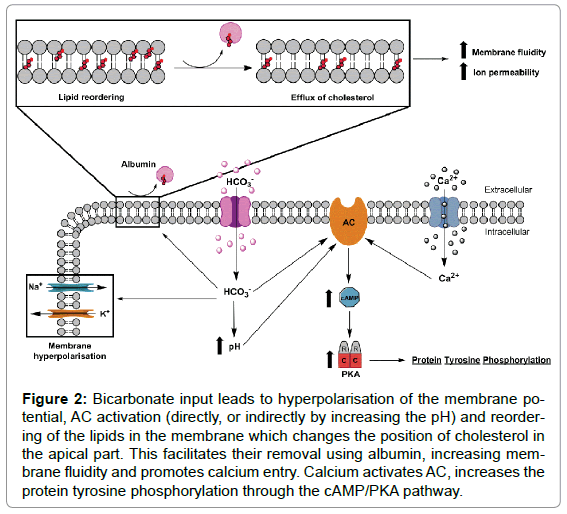
Figure 2: Bicarbonate input leads to hyperpolarisation of the membrane potential, AC activation (directly, or indirectly by increasing the pH) and reordering of the lipids in the membrane which changes the position of cholesterol in the apical part. This facilitates their removal using albumin, increasing membrane fluidity and promotes calcium entry. Calcium activates AC, increases the protein tyrosine phosphorylation through the cAMP/PKA pathway.
Bicarbonate and sperm capacitation
Several studies have shown that bicarbonate plays a key role in sperm capacitation and therefore achieve fertilization under both in vivo and in vitro [34-38]. Epididymal spermatozoa are exposed to low bicarbonate concentrations (3-4mM). However, when they arrive before the capacitation, takes place (oviduct). They are found in much higher level (> 20mM) [39]. Movement of HCO3 through the membrane has been associated with increased intracellular pH during capacitation [40]. Moreover, another likely target for the action of bicarbonate on sperm metabolism is the regulation of cAMP [41] by stimulation of sAC [23]. This in turn stimulates PKA to phosphorylate substrates, thereby allowing PY [42,43]. Furthermore, activation of the PKA results in activation of phospholipase D (PLD), which in turn stimulates the polymerization of F-actin [44], which is an event associated with the process of acrosome reaction.
Bicarbonate and lipid membrane structure: Bicarbonate also modifies the lipid structure of the sperm plasma membrane during capacitation and it is a reversible phenomenon (Figure 2) [36]. Gadella and Harrison [45] showed that influx bicarbonate during capacitation produces change in the lipid membrane structure using path sAC/ cAMP/PKA and so is augmented by inhibitors of phosphatases [46]. These changes lead to a reordering of membrane phospholipids phosphatidylethanolamine, phosphatidylserine, sphingomyelin and phosphatidylcholine. Lipid reordering allows relocating the cholesterol in the apical part of the sperm head. Apparently, this relocation has the function of removal of cholesterol [47]. Albumin, High-Density Lipoprotein (HDL), and β-cyclodextrines promote sperm capacitation acting as acceptors of cholesterol by removing it from the plasma membrane [48]. As a result of this process, decrease ratio of cholesterol/ phospholipid consequently contributes to an increased membrane fluidity promoting increase of ion permeability [32,49-51].
Bicarbonate and sterol depletion: Albumin acts in synergy with bicarbonate by mediating efflux of sterols from the sperm surface [52,53]. Flesch et al. [47] observed that the addition of albumin causes cholesterol efflux (Figure 2), but only in bicarbonate-responding cells that exhibited virtually no filipin labelling in the sperm head area. In the absence of bicarbonate, albumin had no effect on other lipid components and no affinity to cholesterol. Bicarbonate also induces sperm surface oxysterol formation by activation of signalling pathway of the ROS, which can be inhibited or blocked by addition of antioxidants as vitamin E or vitamin A [38]. These sterols oxidation products (oxysterols), which are more hydrophilic, can be extracted using albumin [53] or can facilitate an oxysterol dependent scavengersensitive transport of free sterols to albumin [54].
Bicarbonate and sperm plasma membrane potential: Under normal conditions, spermatozoa maintain intracellular ion concentration markedly different from extracellular environment and these differences provide the resting membrane potential [55]. When spermatozoa are exposed to different environments during transport through the male and female genital tracts, they find different extracellular ion concentration. For example, the epididymal fluid contains high K+, low Na+, and even lower concentrations of HCO3 -. After ejaculation, there will be a drastic change in the concentrations of these ions in the seminal fluid and finally into the female tract, where the concentrations of low potassium and high HCO3 - are present [56,57]. As a result of changes in extracellular ion concentrations, there will be changes in intracellular concentrations of these ions leaving alterations in membrane potential [58,59] which consequently occurs in the hyperpolarisation of sperm plasma membrane [60]. It has been shown in mouse sperm that changes in membrane potential do not occur in BSA or HCO3 - absence [59]. These results suggest that HCO3 - present in capacitation media as well as cholesterol efflux may have a direct or indirect function of events allowing hyperpolarisation of the sperm plasma membrane [55]. Arnoult et al. [61] showed that only hyperpolarized sperm populations are capable of undergoing the acrosome reaction in presence of solubilised zona pellucida material.
Calcium and sperm capacitation
In 1915, Loeb [62] was the first to demonstrate that Ca2+ is required in the extracellular medium for fertilization to occur in invertebrates. Of all intracellular signalling mechanisms, perhaps the most studied and best characterized one is the mobilization of Ca2+. This pathway involves transitory increase of intracellular calcium concentrations produced by multitude intercellular messengers.
One of the most important consequences of cholesterol efflux from membranes is a massive influx of extracellular Ca2+, which is considered a prerequisite for the acrosome reaction process [63]. This Ca2+ influx may be due to changes occurring in the membrane fluidity. The intracellular Ca2+ increase in sperm can activate one or more enzymatic pathways (Figure 2). For example, the Adenylate Cyclase (AC) increases during capacitation in response to Ca2+, this enzyme will catalyse the conversion of ATP to cAMP (revised by [48]).
In 1998, Visconti and Kopf [8] suggested a cooperative effect of Ca2+ and HCO3 -in modulating sperm capacitation requiring the presence of both as well as increase in cAMP levels and the subsequent phosphorylation of different proteins. In swine, both Ca2+ and HCO3 - appear to be required for capacitation and their roles are synergistic, since it has been shown that theHCO3 -will stimulate the entry of Ca2+ in this species [64]. However, in mouse spermatozoa, Tateno et al. [65] showed that Ca2+ ionophore A23187 can make spermatozoa capable of fertilizing in vitro without activation of cAMP-dependent phosphorylation pathways in media bicarbonate free.
Ca2+ is important to sperm hyperactivation during capacitation. CatSperm 1 and 2 are voltage dependent calcium channels that are located in the tail of the sperm. Sperm from mice deficient in these calcium channels are infertile and do not exhibit hyperactivation during capacitation despite having PY [66].
Another aspect that influences capacitation related to calcium is intracellular pH. Sperm not capacitated maintain an acidified intracellular pH [67]. This fact acts as a regulator of calcium influx [68] preventing capacitation and acrosome reaction. Intracellular pH becomes more alkaline during capacitation [69]. Today it is believed that increasing intracellular calcium, bicarbonate and the pH during sperm capacitation produce sAC activation and consequently cAMP [9,51,70,71].
In addition, calmodulin, which is a protein binding Ca2+ considered to be an important transducer of calcium signals, appears to be diminished during capacitation. This mechanism could be based on inhibition of Ca2+-ATPase plasma membrane by increasing cAMP levels through PDE1 inhibition (reviewed by [72]).
Tyrosine phosphorylation of sperm proteins
Protein phosphorylation or de-phosphorylation is controlled by activity of protein kinases and protein phosphatases, which provide cells a “switch” through which they can activate function of various proteins [73]. Phosphorylation occurs in serine, threonine, and tyrosine. PY is related to capacitation process and sperm hyperactivation in many mammal species (human [74], bovine [75], murine [8] or porcine [76]). In opposite, it has been shown that protein phosphatases decrease their activity during capacitation [73].
Increasing PY during capacitation is regulated by a cAMPdependent pathway which involves PKA [77]. cGMP-PKG pathway is also involved in this process [78]. In 2002, Visconti et al. [79] described the possible mechanisms, which could regulate the PY dependent signalling pathway cAMP/PKA: a) the direct or indirect stimulation of a tyrosine kinase by PKA, b) the direct or indirect inhibition of a tyrosine phosphatase, and c) direct or indirect phosphorylation of proteins by PKA on serine or threonine residues to prepare these proteins for subsequent phosphorylation on tyrosine residues.
PY is specific for each species. For example, in man PY during sperm capacitation requires the presence of BSA, and HCO3 -but no calcium [80]. In the case of stallion PY during capacitation requires HCO3 -but neither BSA nor calcium [81]. Another factor to consider in PY is time. PY in boar sperm occurs close to 1 hour after the addition of bicarbonate [82], whereas in bull sperm it occurs 4 hours after addition of heparin [75].
Although PY is an important key in capacitation, it is not yet entirely clear how the phosphorylation of these proteins is involved in sperm-zona recognition, gamete interaction, or exocytosis of acrosomal content [83]. The level of PY in human sperm correlates strongly with the sperm-zona-binding capacity [84] and alterations in PY have been found in subfertile subjects [85] indicating its physiological role in fertilization. In pigs ejaculated spermatozoa selected in the oviduct adhere to the epithelial cells and suppress PY of sperm proteins. This modulation by the oviductal epithelium on PY and, therefore capacitation could help synchronize sperm functions to the time of ovulation [86].
Nitric oxide (NO) and sperm capacitation
NO, a highly ROS, has been found in several physiological systems and regulated manifold functions in male and female reproductive systems [87].
NO is a very small lipophilic molecule that can rapidly diffuse through biological membrane barriers and acts as an intracellular and extracellular biological messenger in a variety of physiological processes. NO is synthesized in vivo from L-arginine by the action of NOS (Figure 3), an enzyme existing in three isoforms: neuronal NOS (nNOS or NOS1), endothelial NOS (eNOS or NOS3), also referred to as constitutive NOS, responsible for the continuous basal release of NO, and both require calcium/calmodulin for activation [88,89]. A third isoform is an inducible calcium-independent form (iNOS or NOS2). NOS activity is dependent on substrate availability and the co-factors NADPH, Flavin Mononucleotide (FMN), Flavin Adenine Dinucleotide (FAD) and tetrahydrobiopterin (BH4). The availability of these factors determines the cellular rates of NO synthesis [90].

Figure 3: Nitric oxide synthase produces NO from L-arginine and different co-factors (NADPH, FMN, FAD and BH4). NO activates three different pathways. A) Activation of Soluble Guanylate Cyclase (sGC); B) Protein tyrosine phosphorylation and C) Modulation of protein function by S-nitrosylation. This figure represents calcium dependent isoforms (nNOS and eNOS); in the case of calcium independent isoform (iNOS) the calmodulin is not present.
Different NOS isoforms were detected in mammalian spermatozoa such as mouse [91], bull [92], human [93,94] and boar spermatozoa [95] activating the biosynthesis of NO. NO was able to affect sperm motility [94,96,97], acrosomal reaction [98,99], acts on PY of sperm proteins [100,101] and enhancement of sperm-zona pellucida binding ability [102].
NO has different functions in the spermatozoa, acting on different pathways that result in sequential and parallel processes (Figure 3). The main actions of NO are:
A. Activation of soluble guanylate cyclase (sGC)
The most important intracellular signalling role for NO in the spermatozoa is its capacity to activate the soluble isoform of Guanylate Cyclase (sGC) [103]. Activation of sGC (Figure 4) leads to increase in intracellular levels of cGMP, which has been implicated in several sperm signalling pathway functions, such as capacitation, acrosome reaction, chemotaxis, and sperm–egg interaction [20,27,99,104].
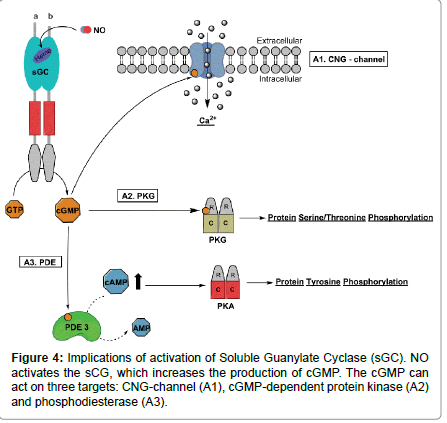
Figure 4: Implications of activation of Soluble Guanylate Cyclase (sGC). NO activates the sCG, which increases the production of cGMP. The cGMP can act on three targets: CNG-channel (A1), cGMP-dependent protein kinase (A2) and phosphodiesterase (A3).
There are at least three targets of cGMP: Cyclic Nucleotide- Gated (CNG) channels, cGMP-dependent Protein Kinase (PKG) and Phosphodiesterase (PDE), involved in several physiological events in mammalian spermatozoa. All of these targets result in increased levels of intracellular calcium and the phosphorylation of different proteins causing sperm hyperactivation and acrosome reaction. Calcium influx together with increased protein phosphorylation brings about the capacitation response (Figure 4).
A1: Cyclic Nucleotide-Gated Channels (CNG): CNG channels have been expressed in mammalian sperm [105] mainly along the length of the flagellum [105,106]. They are activated by cGMP and have been proposed to mediate the influx of Ca2+ to the cytoplasm during capacitation in mammalian spermatozoa controlling sperm motility [106]. This signalling pathway involving CNG channel activation using cGMP is one of the first events that occurred during capacitation in the mouse sperm (Figure 4A1) [78].
A2: cGMP-dependent Protein Kinase (PKG): PGK is a major cellular receptor of cGMP and plays important roles in cGMPdependent signal transduction pathways. Previous studies have identified in mammals two forms of PGK (I and II) [107,108], that are encoded by distinct genes and two different isoforms of PGK-I (designated Ia and Ib) that are produced by alternative splicing [109]. PGK-I seem to play an important role in mediating the acrosome reaction [20,99], modulating several sperm motion patterns and sperm chemotaxis.
The increase in cGMP in the cytoplasm and the subsequent activation of PKG [107,108] results in protein serine/threonine phosphorylation (Figure 4A2) [110], which might also indirectly, mediate a new calcium entry [78]. This promotes sperm capacitation and acrosome reaction [111].
A3: Phosphodiesterase (PDE): Evidence was provided for the involvement of PDE in sperm motility and capacitation [112]. cGMP and cAMP compete for catalytic sites of PDEs that hydrolyse both cyclic nucleotides [113,114]. A rise in intracellular levels of cGMP could inhibit cAMP degradation via cyclic nucleotide phosphodiesterase type 3 [115], which increase intracellular cAMP levels and, consequently, cause an activation of PKA [116] and indirectly increase protein (PY) (Figure 4A3).
B. Protein tyrosine phosphorylation
NO appears to be involved in PY through different mechanisms, acting on two essential pathways for sperm capacitation: on cAMP/ PKA or Extracellular Signal Regulated Kinase (ERK) pathway (Figure 5). These mechanisms for the control of PY are not mutually exclusive neither excluding, both pathways act in parallel [28].
NO can influence the cAMP/PKA pathway by activation of sGC (see above) it could also modulate directly sperm Adenylate Cyclase Activity (AC). The activation of PKA represents the point of convergence for these two pathways. Low concentration of NO could stimulate AC with a subsequent increase in cAMP levels [117] to increase PY by activation of PKA. However, high concentrations of NO can inhibit AC [118]. McVey et al [118] also demonstrated that the effects of NO on AC activity are reversible, suggesting S-nitrosylation of AC as a possible mechanism of action of the NO (Figure 5).
ERK pathway is a chain of many proteins (Shc, Grb2, Sos, Ras and ERK1/2 module, which includes three kinases sequentially activated: Raf, MEK, and ERK1/2). NO intervenes in the middle of the pathway, modify the Ras structure [119], reacting with cysteine residues [28] and inducing its activation. Ras interact with Raf by activating it [119], leading to MEK activation. MEK phosphorylates Thr and Tyr are residues within the Thr-Glu-Tyr motif, which are located at the active site of ERKs 1 and 2 (Figure 5). All this process is necessary for the subsequent PY [28] and is involved in the acquisition of sperm motility [120].
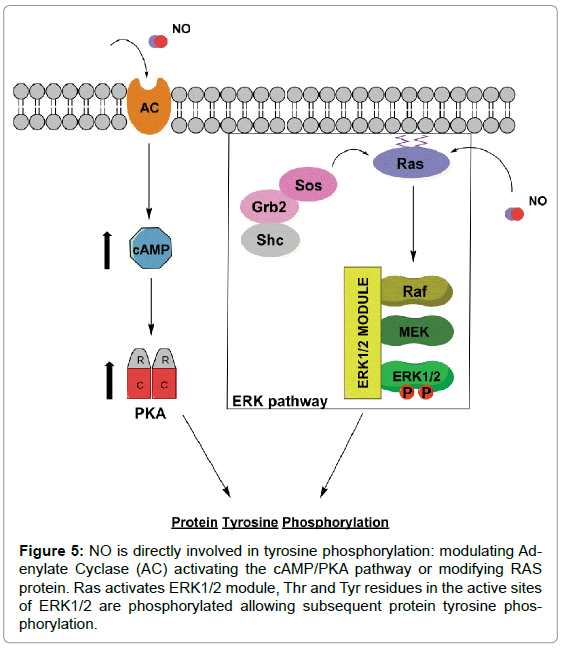
Figure 5: NO is directly involved in tyrosine phosphorylation: modulating Adenylate Cyclase (AC) activating the cAMP/PKA pathway or modifying RAS protein. Ras activates ERK1/2 module, Thr and Tyr residues in the active sites of ERK1/2 are phosphorylated allowing subsequent protein tyrosine phosphorylation.
C. Direct modulation of protein function by S-nitrosylation of exposed cysteine residues
Mature sperm lack the necessary machinery for the transcription or protein modification and thereby require post-translational modifications to control the activity of proteins. NO participates in protein regulation, which acts directly on protein targets (exposed cysteine residues) via S-nitrosylation [121,122]. S-nitrosylation is a regulated post-translational protein modification (Figure 6), analogous to phosphorylation and acetylation [123,124], which involves the covalent incorporation of a NO into thiol groups (-SH), to form S-nitrosothiol (S-NO). This modification is selective, reversible and stabilizes NO in a uniquely bioactive form. Lefièvre et al. [125] described numerous sperm proteins that can be nitrosylated in human sperm. Some of the S-nitrosylated proteins are involved in processes related to sperm capacitation as energy generation, sperm motility [125] or hyperactivation [126,127]. However, how some of nitrosylated proteins perform their function remains unknown.
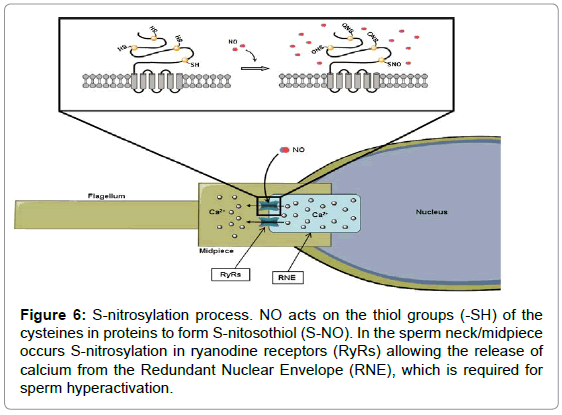
Figure 6: S-nitrosylation process. NO acts on the thiol groups (-SH) of the cysteines in proteins to form S-nitosothiol (S-NO). In the sperm neck/midpiece occurs S-nitrosylation in ryanodine receptors (RyRs) allowing the release of calcium from the Redundant Nuclear Envelope (RNE), which is required for sperm hyperactivation.
For sperm, hyperactivation is necessary mobilisation of stored Ca2+ in the sperm neck/midpiece [127]. The calcium store in the neck of the sperm takes place in the Redundant Nuclear Envelope (RNE) [128]. To mobilize calcium from these stores is necessary to enable ryanodine receptors (RyRs); intracellular calcium release channels involved in regulation of cytosolic calcium levels [129]. These proteins contain a large number of thiol groups and are thus subject to S-nitrosylation by NO [130,131]. S-nitrosylation can potentiate opening of RyR [132- 137], probably by generations of the membrane permeant product cys-NO [138]. A NO-induced Ca2+ elevation was accompanied by an increase in S-nitrosylation levels of endogenous RyR [139,140] and also inhibition of these Ca2+ channels can occur under strongly nitrosylating conditions or at high doses of NO (Figure 6) [134,137,140].
Progesterone acts synergistically with NO (by S-nitrosylation) to mobilise Ca2+ at the sperm neck/midpiece (by activation of RyRs) [126] contributing to the hyperactivation that is vital for penetration of the egg vestments.
The sperm from being deposited in the female genital until it reaches the place of fecundation undergoes a series of changes known classically as capacitation. This process involves modifications membrane lipids, loss of cholesterol, activation of cAMP/PKA pathway, increase Ca2+ uptake and pH (pHi), hyperpolarisation of membrane potential and tyrosine phosphorylation. Among the most studied molecules associated with this process include BSA as cholesterol acceptor, bicarbonate as an activator of the cAMP-PKA-tyrosine phosphorylation, and calcium as an activator of channels voltagedependent and hyperactivity motility process. However, there are other pathways in relation to capacitation as NO/sGC/cGMP, which have some common steps to cAMP-PKA-tyrosine phosphorylation pathways. Another pathway recently described relating to capacitation process is the protein nitrosylation. Nevertheless, this new way of signalling involves numerous proteins whose functions are yet to be determined and that can be important to understand the complex process of sperm capacitation.
Supported by Spanish Ministry of Economy and Competitiveness and FEDER, Grant AGL2012-40180-C03-01