Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Review Article - (2016) Volume 7, Issue 1
Pesticide residues with genotoxic potential reach the aquatic environment and constitute a major issue that give rise to concerns at local, regional, national and global scales. Fish serves as an excellent genetic model for the genetic hazard assessment as they are very sensitive to changes in their environment. As there is a close consortium of DNA damage, mutation and induction of various types of genetic disorders, genotoxicity tests like chromosomal aberration test, micronuclei and comet assay are gaining credence and since past few decades many tests have been developed for evaluating genetic alterations in aquatic organisms. These tests rely on the premise that any change to DNA may have enduring and ardent consequences. Thus, the first aim of the genotoxicology is to describe the outcome produced by toxic substances in various test species, but only from the genetic point of view and to draw conclusions that can be extrapolated to man. This review summarizes the genotoxicity tests developed till date and the role of piscine model in genotoxicology. The text also delves the latest knowledge and thinking on these cardinal approaches for the assessment of aquatic environmental health, management and conservation, besides providing useful repository for the researchers especially dealing with aquatic genotoxicity tests. The perspectives for further research on the use of genotoxicology tests were also highlighted.
Keywords: Piscine model; Genotoxicity methods; Chromosomal aberration test; Micronucleus assay; Comet assay; Aquatic ecosystem
The demand for a clean and immune supply of water for drinking, agriculture and recreation has accrued since the last few decades. The global production of chemicals has increased from 1 million ton in 1930 to more than 400 smillion tons during the year 2005 and about 100,000 different substances have been registered in the market, of which 30,000 to 70,000 are in daily use [1]. Most of the chemicals used on land enter the aquatic environment via., wastewater from agricultural, industrial and domestic sources including municipal sewage treatment plants [2]. In some instances, these chemicals are capable of permanently altering the genetic material of exposed organisms, i.e., they are mutagens. In recent years, reports on the occurrences of malignancies and other biological conditions in aquatic organisms following exposure to suspected genotoxins have increased. Such exposure of aquatic fauna not only poses a high risk for non-target organisms including man via food chain in the ecological context, but also may lead to heritable mutations and loss in the genetic diversity with significant implications of the long term survival of exposed population. The studies related to the impact of genotoxic exposure in an organism, therefore, not only cover an attempt to elucidate the mechanism(s) involving direct changes to the genetic material, but also the genotoxic effects and changes at the level of gene pool and population [3].
The uncontrolled presence of genotoxins in aquatic environment poses significant toxicological risks to a myriad of non-target organisms and finds its way to the food chain, threatening the ecological balance and the biodiversity of the nature [4]. This is an unwanted situation and justifies the monitoring of the aquatic environment for the presence of genotoxins. Although it is clear that progress in any field of research depends primarily on the development of its methodology. In this context, recent advances in the development of techniques and diagnostic tools have certainly enhanced the understanding of how an organism interacts with its environment at individual, cellular and molecular level, and substantial progress has been made to assess the genotoxin induced genetic damage in the aquatic biota [3]. The biomonitoring of aquatic genotoxicity is cardinal for many folds. Initially, the detection of genotoxic effects in aquatic environment is needed to assess the health of aquatic organisms. Second, from the ecological perspective, the protection of genetic diversity in natural populations is important for population survival [5]. The exposure to genotoxins can damage the DNA of living cells and if these DNA lesions are not repaired, they can commence a cascade of biological consequences at the cellular, individual, community and finally at the population level [6]. The relevance of detecting the genotoxic risk assessment in contaminated aquatic environment(s) was firstly perceived in the late 1970’s. Since then several tests have been flourished for evaluating genotoxicity in aquatic organisms. The possibility of using changes in DNA integrity to the genetic material as markers of exposure and effect of genotoxicants has been previously investigated [7]. The presence of DNA adducts has been taken as evidence of exposure to specific genotoxins [8]. These tests rely on the premise that any changes to DNA may have chronic and profound consequences [9]. Although some studies have highlighted the state of the art of methodologies from genotoxicological perspective [10-12], this review addresses this question again and to place it in somewhat a broader context. This review addresses the genotoxicity tests developed to date besides delving the latest knowledge and thinking on these cardinal approaches for the assessment of aquatic health, management and conservation.
Genotoxicology is the systematic investigation of the effect(s) that a physical or chemical agent can exert on the genetic system of an organism and when present in our environment, interaction of such chemicals may give rise to cumulative effect(s), with genetic consequences for the future of the affected species. The genotoxicity is often confused with mutagenicity; but while all mutagens are genotoxic, not all genotoxic substances are mutagenic. The alteration can have direct or indirect effects on the DNA: the induction of mutations, mistimed event activation, and direct DNA damage leading to mutations. The primary objective of the genetic toxicology, however, is to detect and analyze the potential hazard of those agents that are highly specific for interaction with nucleic acids and produce alterations in genetic material at sub-toxic concentration. Thus, its first aim is to describe the outcome produced by toxic substances in various test species, but only from the genetic point of view and to draw conclusions that can be extrapolated to man. The next aim is to investigate the mechanism(s) of action of the toxic substances, and on the basis of this knowledge to assess the environmental risks [13]. The beginning of the genotoxicology was marked by H.J. Muller’s discovery of sex linked recessive lethal mutation in Drosophila induced by the x-rays. The impetus in evolving the field of genotoxicity came from the study of Alexander-Hollaender, who led the foundation of the Environmental Mutagen Society (EMS), in USA [14]. The EMS and, in particular, its applied role in genotoxicology, has had a profound positive impact on many disciplines in toxicology and environmental risk assessment [15]. In 1973, after the establishment of International Association of Environmental Mutagen Societies (IAEMS), the beginning of active research in the field of genotoxicology was marked. National and international agencies became increasingly interested in mutagenesis, recognizing its importance not only for genetic diseases but also for carcinogenesis [13]. In the United States, agencies like National Institute of Health (NIH), National Toxicology Programme (NTP), Environmental Protection Agency (EPA), Food and Drug Administration (FDA) have supported the basic research in mutagenesis.
Agents that induce alterations in genetic material at sub-toxic exposure level are classified as genotoxic. These agents usually have common chemical or physical properties facilitating their interaction with genetic material. Most genotoxic agents damage nucleic acids at high exposure concentrations evoking acute and nonspecific cytotoxic effects in wide range of cellular processes. Thus, genetic toxicology has evolved to play a dual role in safety evaluation programmes. One role is the implementation of mutagenicity testing and risk assessment methods to evaluate the impact of genotoxic agents found in the environment, more specifically of those whose presence may alter the integrity of gene pool. The second role is the application of genetic test methods to the detection and mechanistic understanding of carcinogenic chemicals [16]. However, the ecological impacts of many genotoxins still remain poorly understood. The history of genotoxicology and electronic data collections may be reviewed online as the internet offers a wide range of digital resources for the field of genotoxicology. The popular Web based resources are TOXNET, GENE-TOX, TOXLINE, MEDLINE, PUBMED, Gateway, Entrez, Hazardous Substances Data Bank, CHEMIDPLUS at NLM, etc., As the molecular biology and computational tools improve, new areas such as structural activity relationship analysis, mutational spectra databases and toxicogenomics, which are the emerging areas within genotoxicology, have online resources as well [17].
Research on the genotoxic effects of different pollutants to fish has been primarily directed at determining the potential for chemical mutagenesis in these organisms. However, clastogenic activity may be a risk factor for genetic diseases in fish population, in addition to carcinogenesis. There is also continuing interest in developing in vivo genotoxicity assays with piscine model systems for assessing health risk to higher vertebrates [12]. Aquaculture is one of the most rapidly expanding food industries since fish have always been an important source of food for humans and, for large proportion of the population they represent a major source of protein [18]. On the other hand, the aquatic environment is a sink for many chemicals, as illustrated by the occasionally high pollution levels and frequencies of chemical spills, and fish have been the target of overt chemical pollution [1]. Fish have received particular attention as monitor system in the surveillance of aquatic ecosystem and indirectly the human health. For example, marine fish and shell fish, which are a major source of protein in many countries, are often contaminated with high concentrations of methyl mercury. Induction of chromosomal damage has been found in the lymphocytes of persons exposed to methyl mercury through consumption of contaminated fish [19].
In conventional eco-toxicity testing strategies, fish has been regarded as an indispensable component of integrated toxicity testing strategies for the aquatic organisms. The current guidelines at the OECD level are a good example of this with fish-targeted guidelines covering acute toxicity [20], early life stage toxicity [21] and juvenile growth test [22]. In particular, fish acute toxicity tests play an important role in environmental risk assessment and hazard classification, because they allow for first estimates of the relative toxicity of various chemicals in various species [23]. They have become widely required for the aquatic hazard assessment of new and existing synthetic substances as well as for use in the evaluation of complex effluents. Thus, in the “base set” of data requirements for all substances which are produced or imported in quantities of one ton or more per year, the acute toxicity for freshwater fish [20] is a mandatory component. Furthermore, no single species is universally sensitive to all chemicals. The single species LC50 data are questionable with respect to accuracy and, in more general terms, to toxicological relevance. Thus, it seems largely accepted that there is no most susceptible species that could be used in a possible conservative testing approach [24]. The retreat of the toxicity tests with the single most sensitive species is to ignore any knowledge of the complexity of ecosystems. Therefore, the susceptibility and tolerance of most fish populations may be hard to predict from single standard acute toxicity test. Multiple tests on various species are necessary to get an appropriate estimate of the range of susceptibilities to chemical substances.
The current spurt in the fish cytogenetical studies had its origin in the standardization of the newer techniques and in the realization of an immense applied value of the cytogenetic data of fish. The fish genotoxicity indeed stands on the threshold of a golden era with great prospects of its being used in applied research, such as monitoring of genotoxic agents [25]. Since most of the chemicals and pesticides used terrestrially end up finally in the aquatic system, it is the fish that are affected and should be monitored for these subtle but genetically damaging changes [26]. Induction of cytogenetic damage in fish could serve to monitor not only selected genotoxic agents in the laboratory, but also the presence of genotoxicants in surface waters and different aqueous ecosystems [27]. Several testing protocols have been developed earlier which permit evaluation of the mutagenic properties of various chemicals and physical agents. Short term tests include acute toxicity, sister chromatid analysis, gross chromosomal changes, formation of micronuclei and effects on DNA. Long term studies can be carried out on mutagenicity and general toxicity. The modes of administration may be oral, intraperitoneal or in aqueous environment [28]. Exposure of fish to genotoxic agents may occur either by the digestive route or by the respiratory route following absorption of water borne chemical through the gills [12]. The fish is able to convert xenobiotics to active metabolites. It has also been successfully used to demonstrate the presence of genotoxic agents in the aquatic environment [12,27].
The genotoxicity of various chemical agents in fish and other aquatic organisms have been assessed by several assays. These assays differ in their end points and specificities to explore the genotoxic effects induced by various toxicants at different levels in biological systems, but most of them are having their own limitations.
Chromosomal aberrations (CA) are one of the important biological consequences of exposure of the target organism to genotoxic agents. Chromosomal studies have received considerable attention in recent years, in part from a growing interest in the evaluation of genotoxicity of environmental toxicants and carcinogens. Chromosomal changes leading to mutations were first described in Oenothera [29]. During 1930-1940 the CA along with their mechanism, behaviour and fate, following irradiation was studied in some insects like Drosophila and in plant systems, i.e., Allium and Tradescantia. The protocols gradually developed following the discovery of colchicine which was able to arrest cells at metaphase. However, studies of chemical effects of chromosomes were mainly limited to academic exercises till 1970’s, when concern over the deteriorating environmental situation led to an increasing interest for effective short term assays to screen mutagens and carcinogens. The cytogenetic tests emerged as a very important component of these systems [30].
CA may be defined as a missing, extra, or irregular portion of chromosome. A karyotype refers to a full set of chromosomes from an organism that can be compared to a normal karyotype for the species via genetic testing. CA usually occurs when there is an error in cell division following meiosis or mitosis. Cytogenic effects can be studied either in whole animals (in vivo) or in cells grown in culture (in vitro). In both cases, the animal or cell is exposed to the test substance and then afterwards treated with a metaphase-arresting Colchicine or Colcimide. Following suitable staining the metaphase cells are analysed microscopically for the presence of aberration. There are many types of CA and can be organized into two basic groups, structural and numerical aberrations.
The pioneering studies of Howard and Pelc [31] demonstrated that between two mitoses, the intervening interphase has three phases designated as G1 (pre-DNA synthetic), S (DNA synthesis) and G2 (post-synthetic). Ionising radiation induced chromosome type of aberrations in G1, chromatid type of aberrations in G2 and most of the S phase. True radiomimetic chemicals, such as bleomycin and neocarcinostatin induce similar pattern of aberrations as ionising radiation. However, chemical mutagens, which do not induce directly DNA strand breaks but cause other lesions, were shown to induce only chromatid type aberrations irrespective of the DNA synthetic stage treated. An intervening S phase is necessary to visualize the aberrations. Short wave UV induces aberrations in a similar way as chemical mutagens. On this basis, chromosome breaking agents (clastogens) can be classified as S dependent and independent agents [32].
The morphological and numerical alteration of chromosome carrying DNA (genes) results in a quantitative alteration or rearrangement of genes and has been substantially studied in various research areas. The mammalian in vivo chromosomal aberration test has been successfully used for the detection of structural aberrations induced by the test substance in the bone marrow cells of animals, usually rodents [33,34]. In fact the chromosomal aberration test using cultured mammalian cells is one of the sensitive methods to predict environmental mutagens or carcinogens, and considered as complementary test to the Salmonella microsome assay (Ames test) by some workers [35].
Of the estimated 20,000 fish species, a mere 500 have been studied for complete karyotype. In Indian context, the figure is far too less; the fish cytologists have determined the karyotypes of only about 125 species to a satisfactory extent [25]. Genotoxic studies in different fish species using cytogenetic analysis have been reported by a number of workers [27]. Exposure of fish to various toxicants for a prolonged period, even at low levels, leads to chromosomal aberrations including gene alterations [25,36,37]. An advantage of chromosomal studies is that they reveal a measure of sub-lethal effects of xenobiotics in vivo.
Most fish typically have high chromosome numbers, greater than 48 with small acrocentric chromosomes [38]. Given to this nature, different techniques have been used to perform cytogenetic studies; direct in vivo and indirect in vitro. Direct techniques are based on the use of colchicine to block quickly proliferating cell population of kidney, spleen and gill epithelia at the metaphase stage of the cell cycle. Just after the fish is sacrificed, cell samples are taken and treated for slide preparations [39]. The indirect test technique involve tissue cultures [40], that allow for the application of perfect doses of colchicine and the use of mitotic stimulants such as phytohaemeagglutinin (PHA). Each of this procedure has been optimized to obtain large number of well spread metaphases and used regularly for karyotypic analysis, and to assess the impact of genotoxic agents.
Fish as an in vivo model can be quite promising for screening the mutagenic properties of chemicals and environmental pollutants specially the agricultural runoff that routinely pollute our water bodies [12]. Kligerman is credited to develop a cytogenetic fish model (Umbra limi) in which chromosome damage was later accurately detected using physical and chemical agents [39]. The toxicity of different pesticides has been associated with changes in replication of DNA and DNA aberration that leads to mutation and hyperproliferation of cells due to local irritation [41]. In fish DNA repairing takes place at a much lower speed than in mammals [42], which render fish as a sentinel organism as far as bio-monitoring studies are concerned [4,12]. There is rich documented literature witnessing research on molecular level of different fish species showing ill effects of pesticides on genes and DNA levels [43-45].
The genotoxicity of contaminated water in the field and laboratory, for various in vivo and in vitro monitoring systems have successfully employed the fish cells. CA has been observed after exposure of fish species to different chemicals. Exposure of Channa punctatus to Dichlorvos (0.01 ppm) caused chromatid gaps, centromeric gaps, attenuation, chromatid breaks, extra fragments, and stubbed arm in kidney cells [25]; while exposure of the same species to fenvalerate caused chromatid separation, chromatid break, deletion, fragments, gaps, and ring type chromosomes [46]. Interestingly longer exposure was associated with lower frequency of DNA aberrations. Cypermethrin caused changes in nucleic acids (RNA and DNA) in gonadal tissue of Colisa fasciatus. This test was equally effective in cultured cells and embryos in some parallel studies of fish and as well as on cell lines of Umbra limi [2,43,47].
There are several studies demonstrating the suitability of conventional cytogenetic techniques like chromosomal aberration test for the assessment of genotoxic effects of various toxicants using fish as bioindicator. Table 1 presents a list of some important studies, using fish as a model, to evaluate genotoxicity by chromosomal aberration test.
| Fish | Chemical(s)/pollutants | Reference |
|---|---|---|
| Notobranchus rechowi | Ethylmethane-sulphonate and benzo(a)pyrene | [48] |
| Boleophthalmus dussumieri | Mitomycin-C, mercury, selenium and chromium | [49] |
| Oreochromis mossambicus | Malathion | [50] |
| Cyprinus carpio | Cadmium; nitrate | [51] |
| Clarius lazera | Ethylmethane sulfonate | [52] |
| Channa punctatus | Dichlorvos | [25] |
| Heteropneustus fossilis | Pentachlorophenol | [53] |
| Oreochrochromis mossambicus | Fenvalerate | [54] |
| Channa punctatus | Heavy metals | [55] |
| Etroplus suratensis | Methyl parathion and Phosphamidon | [56] |
| Oreochromis niloticus and Clarias lazera | Agricultural and industrial waste water | [57] |
| Oreochromis mossambicus | Ethylmethane sulfonate | [58] |
| Channa punctatus | Cypermethrin | [59] |
| Mystus gulio | Lambda-cyhalothrin | [60] |
| Clarias gariepinus | 2,4-dichlorophenoxyacetic acid | [61] |
| Boleophthalmus dussumieri | Bleomycin, Mitomycin-C and Doxorubicin | [62] |
| Oreochromis mossambicus | Carbamate pesticide methomyl | [63] |
| Oreochromis niloticus | Copper sulphate and lead acetate | [64] |
| Channa punctatus | Paper mill effluent | [65] |
| Clarias batrachus | Fluoride | [66] |
| Oreochromis niloticus and Tilapia zillii | Sewage discharge | [67] |
| Channa punctatus | Fenvalerate | [46] |
| Cirrhinus mrigala | Butachlor | [68] |
| Channa punctatus | Cypermethrin | [37] |
| Cirrhinus mrigala | Butachlor | [69] |
| Cirrhinus mrigala | Dyeing industry effluent | [70] |
| Carassius carassius | Endosulfan | [12,71] |
Table 1: Summary of some landmark genotoxicity studies in fish using chromosomal aberration test: a review.
The MNT is recognized as one of the most simple, reliable and successful assays for the in vivo evaluation of genotoxicity of various toxicants. Micronuclei (MN) were first described in the cytoplasm of erythrocytes more than a century ago and were called “fragment of nuclear material” by Howell or “intraglobularies corpuscules” in the terminology of Jolly in the late 18th century and early 1900. These structures are known by the haematologists as “Howell-Jolly bodies” [72]. It is now well-established that MN mainly originate from acentric chromosome fragments, acentric chromatid fragments or whole chromosomes that fail to be included in the daughter nuclei at the completion of telophase during mitosis because they did not attach properly with the spindle during the segregation process in anaphase [73]. These displaced chromosomes or chromosome fragments are eventually enclosed by a nuclear membrane and, except for their smaller size, are morphologically similar to nuclei after conventional nuclear staining. Scoring MN in interphase nuclei is technically easier and faster than scoring of chromosomal aberration in metaphase nuclei [27].
Salvadori highlighted that, whatever the type of DNA damage, MN is originated during cell division [74]. The DNA damage due to exposure to mutagenic agents is expressed in micronucleus form just after one cycle of cell division and is dependent on the proportion of cells under division. The increased interest in environmental genotoxicity studies went ahead with the development of a great number of tests to evaluate genotoxic effects in aquatic environments. Considering this, MNT, one of the most popular and promising tests on ecotoxicology, represents a cytogenetic indicator of DNA damage for over 30 years [73].
The MNT is a simple and sensitive assay for “in vivo and in vitro” evaluation of genotoxicity in aquatic environment as a part of the biomonitoring programs for several days followed by microscopic analysis of erythrocytes, gill, liver, kidney and fin cells [27]. Fish serves as useful genetic model for the assessment of pollution in aquatic ecosystems [75]. Fish respond to toxic agents similar to higher vertebrates and can allow the assessment of substances that are potentially hazardous to humans. Fish can, in fact, be the sentinel organisms that can indicate the risk of human exposure to drinking water contaminated with genotoxicants [72].
In India, studies on MNT have been initiated by Manna who reported presence of micronuclei in erythrocytes and chromosomal aberrations in Oreochromis mossambicus treated with aldrin, cadmium chloride and x-rays [76]. The percentage of micronuclei was low but chromosomal aberrations were higher in x-rays treated specimens. Das and Nanda noted an increase in micronuclei frequency in the erythrocytes of Heteropneustes fossilis exposed to mitomycin C and paper mill effluent [77]. Barat employed the MNT to screen the genotoxic effect of the pesticide Malathion on Channa punctatus at concentration of 0.033 mL/L for periods of 5,11 and 16 days and reported significant increase in the frequency of MN than that of the control [78]. MNT have been reported to be sensitive for detecting the genotoxic potential of the low levels of malathion pesticide in two fish species namely Mystus vittatus and C. punctatus in aqueous medium and there was also dose dependent increase in MN formation [79]. The study of Ateeq showed a positive dose-response relationship in fish for genotoxic and cytotoxic endpoints when exposed to two herbicides namely 2,4-dichlorophenoxyacetic acid (2,4-D) and butachlor [80]. The genotoxic endpoints in the study were MNT and chromosomal aberration test using C. punctatus as an experimental model. Ahmad studied the effect of pentachlorophenol (PCP) on fish genome using haematoxylin-eosin technique and observed that the frequency of micronucleated erythrocytes increased with the increase in exposure time [81]. Recently, Farah reported the strong genotoxic effects of PCP and 2,4-D in fresh water fish C. punctatus, using MNT and chromosomal aberration test [82]. The ameliorating effect of ethanolic neem leaf extract was also carried out in the study.
MNT has become a very popular tool for assessment of genotoxic potential of various chemical agents by using fish as a model. Table 2 shows some studies on fish, for the evaluation of genotoxicity of various xenobiotic agents, using MNT.
| Fish | Chemical(s)/pollutants | Reference |
|---|---|---|
| Oreochromis mossambicus | Aldrin, cadmium chloride and x-rays | [76] |
| Cyprinus carpio and Tinca tinca | Aflatoxin B1, arochlor 1254, benzidene, benzo(a)pyrene and 20-methylechloanthrene | [83] |
| Heteropneustes fossilis | Mitocycin C and paper mill effluent: allylformate | [77] |
| Esox lucius | Radiocesium | [84] |
| Oncorhynchus mykiss | In situ to heavily polluted tributary of the River Po (Northern Italy) | [85] |
| Carassius auratus gibelio | Selenium, mercury and methyl-mercury | [86] |
| Salmo truttafario | PCB77 | [87] |
| Channa punctatus | Malathion | [78] |
| Oncorhynchus mykiss | A textile industry effluent | [88] |
| Cheirodon interruptus interruptus | Pyrethroid λ-cyhalothrin | [89] |
| Astyanax bimaculatus | Cyclophosphamide, vinblastine sulfate | [90] |
| Channa punctatus | Malathion | [79] |
| Oncorhynchus mykiss | Colchicine, mitomycin, cyclophophamide, acrylamide, methyl-methanesulfonate and N-ethyl-N-nitrosourea | [91] |
| Clarias batrachus | 2,4-dichlorophenoxyacetic acid and butachlor | [80] |
| Heteropneustes fossilis | Pentachlorophenol | [81] |
| Channa punctatus | Pentachlorophenol and 2,4-dichlorophenoxyacetic acid | [82] |
| Anguilla Anguilla, Phoxinus phoxinus and Salmo trutta | Metals, hydrocarbons, pesticides | [92] |
| Cyprinus carpio | Disinfectants (sodium hypochlorite, peracetic acid and chloride dioxide) | [93] |
| Cyprinus carpio, Carassius gibelio, Corydoras paleatus | Cadmium chloride and copper sulphate | [94] |
| Oreochromis niloticus and Tilapia rendalli | Domestic sewage | [95] |
| Scophthalmus maximus | Dialkyl phthalate, bisphenol-A, tetrabromodiphenyl ether | [96] |
| Oncorhynchus mykiss | Mixture of heavy metals | [97] |
| Clarias gariepinus, Oreochromis niloticus and Oreochromis aureus | Heavy metals | [98] |
| Channa punctatus | Chlorpyrifos | [99] |
| Channa punctatus | Malathion | [100] |
| Cnesterodon decemmaculatus | A?cida® (insecticide) | [101] |
| Carassius carassius | Agricultural runoff | [102] |
| Apteronotus bonapartii | Benzene | [103] |
| Labeo rohita | λ-cyhalothrin | [104] |
| Carassius carassius | Endosulfan | [12,71] |
Table 2: Assessment of genetic damage by micronucleus assay in fish after in vivo exposure to genotoxicants.
Since Singh launched the alkaline Single Cell Gel Electrophoresis (SCGE) protocol or comet assay to the scientific community, its uses and applications has been accreting [105]. The thematic areas of its current employment in evaluating the genotoxicology are vast, either in vitro or in vivo, both in the laboratory and in the field, terrestrial or aquatic. The comet assay developed by Singh and his coworkers was successfully acclimatized to fish erythrocytes with few modifications. The authors concluded that the assay as “extremely sensitive and should be useful in detecting DNA damage caused by environmental pollutants.” Since the past two decades, this premonitory statement has been recurrent and increasingly reinforced by an array of scientific publications, exploring a wide diversity of approaches, i.e., in vitro [106], in vivo [107], and as well as surveying wild native specimens [108]. Furthermore, in recent years we have witnessed a greater profusion of publications. For example in the last year (2014), 72 articles were published (according to a literature search on PubMed) evaluating DNA damage by comet assay in fish exposed to various genotoxins (Figure 1). This vast utilization of fish should also be regarded as a sign of primary concern of genetic ecotoxicologists over the health status of aquatic ecosystems.
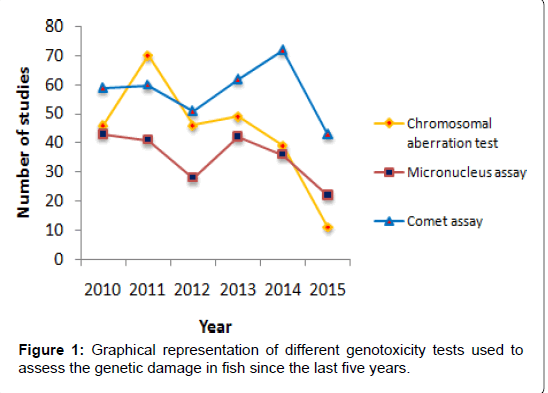
Figure 1: Graphical representation of different genotoxicity tests used to assess the genetic damage in fish since the last five years.
The popularity of this test is due to its sensitivity, simplicity, cost effectiveness and time efficiency due to automatic scoring of the comets by using the image-analysis software. Furthermore, the German Federal Environmental Agency proposes the comet assay as a useful test for a reliable detection of genotoxins in surface waters. Alkaline comet assay is capable of detecting a wide variety of DNA damages such as DNA single-strand breaks, double-strand breaks, oxidatively induced base damages and alkali-labile DNA length: width ratio of the DNA mass in a variety of fish cells (erythrocytes, liver, gill and kidney) exposed to contaminated water samples, and the amount of damage increased with the duration of exposure. It is well-established that comet assay is applicable, virtually, to all species. A clear demonstration of this polyvalence is the finding that, since last two decades, this assay was successfully adapted to more than 90 fish species. This wide range of species includes mostly bony fish (Class Osteichthyes), both ray-finned fish (Subclass Actinopterygii), the overwhelming majority of cases, and lobe-finned fish (Subclass Sarcopterygii) like Arapaima gigas [109]. The jawless fish (Class Agnatha) are represented with an interesting study with sea lamprey (Petromyzon marinus) describing the relationship between sperm DNA damage and fertilizing ability [110], while cartilaginous fish (Class Chondrichthyes) are completely unexplored. Some recent landmark studies on comet assay using piscine model are summarized in Table 3. In what concerns to the type of agent/contaminant tested, the application of comet assay in the field of aquatic genotoxicology has accompanied the evolution of other subareas of environmental toxicology involving piscine models.
| Fish | Cell type | Agent | Exposure | Concentration | Response | Reference |
|---|---|---|---|---|---|---|
| Anguilla Anguilla | Erythrocytes | B(a)P b-BF Arochlor 1254 TCDD | 0.1–50 mg/kg 0.1–50 mg/kg 0.1–50 mg/kg 0.01–2 mg/kg | + | [111] | |
| Carassius auratus | Erythrocytes | PBTA-6 (dye) | 3-9 h | 1–100 mg/kg | + | [112] |
| Danio rerio | Hepatocytesandgill cells | 4-nitroquinoline−1- oxide | 0, 0.1, 0.3, 1.1, and 2.9;μg/l | + - + | [113] | |
| Clarias batrachus | Erythrocytes | Herbicides: 2,4-D Butachlor | 48-96 h | ; 25–75 ppm 1–2.5 ppm | C-R | [114] |
| Channa punctatus | Gill and kidneytissues | Endosulfan (pesticide) | 24-96 h | 4–10 ppb | C-R | [115] |
| Cyprinus carpio | Gill and liver cells | MNNG | 7 days | 0.01–1 ppm (mg/l) | + | [116] |
| C. carpio | Erythrocytes | Lake water (Italy) | 20 days | Seasonal difference | + | [117] |
| Oncorhynchus mykiss | Erythrocytes | Algal extracts (P. fucoides) | 7 days | 0.5% | + | [118] |
| O. mykissO | Erythrocytes | Sediments from Biobio River (Chile) | 21 days | PAHs (2000–7000 ng/g d.w.) | + | [119] |
| C. auratus | Erythrocytes | Leachate from land?ll sites (Japan) | 96 h | + | [120] | |
| C. auratus | Erythrocytes | Glyphosate | 2-6 days | 5–15 ppm | D-R | [121] |
| Plotosuslineatus | Erythrocytes | Diesel water soluble fraction (DWSF) | 6–96 h; 15 days | Diesel oil added to 4 parts water. Water phase dilutedto 50% WSF withdechlorinated water | + | [122] |
| D. rerio | Freshly fertilized eggs | Sediments fromLaguna lake(the Philippines) | 3 h | 430 mg/ml (100%) (1:1); 215 mg/ml (50%); 107.5 mg/ml (25%) | + | [123] |
| Geophagus brasiliensis, Cichla temensis, Hoplias malabaricus, C. carpio and Steindachnerina insculpita | Erythrocytes | Eutrophication | Field study | + | [124] | |
| O. mykiss | Spermatozoa | Cryopreservation | + | [125] | ||
| Labeo rohita | Erythrocytes, liver and gill cells | Phorate | 24-96 h | 0.001, 0.002 and 0.01 ppm | C-R | [126] |
| C. punctatus | Gill cells | Profenofos | 24-96 h | 0.58, 1.16 and 1.74 ppb, | C-R | [127] |
| A. anguilla | Blood andliver | Glyphosate-basedherbicide | 24-72 h | 58 and 116 g l−1 | + | [128] |
| Ictalurus punctatus | Ovary cellline | Water andsediment fromgypsum miningarea | Field study | + | [129] | |
| C.catla, L. rohita | Heart andgillcell lines | Silvernanoparticles | 96 h | Environmental realistic concentration | + | [130] |
(+) = DNA damage; (-) = No DNA damage; (C-R) = Concentration dependent response
Table 3: Genotoxicity studies in fish using the comet assay published during the last twelve years.
For evaluating the environmental samples in the field of genotoxicology, recent advancement has been achieved [131]. A transgenic zebrafish has been established for the detection of genotoxins. It carries plasmids that are having the rpSL gene of Escherichia coli as a mutational target gene [132]. Another transgenic fish, that bikes multiple copies of a bacteriophage lambda vector harboring the cII gene as a mutational target, has been established by a technique originally developed for lambda transgenic rodents [133]. The p53 tumor suppressor gene, which is known to be implicated in cancer development, has been investigated as a possible biomarker for genotoxin in fish cells [134].
The combination of genotoxicity tests with other biomarkers to evaluate the genotoxin(s) based genetic damage has been assessed in many studies (Figure 2). Some of them show a positive correlation between the results given by the genotoxicity tests and other biomarkers. For instance, in one of our previous study [12], a positive correlation between oxidative stress and genotoxicity was observed on endosulfan (persistent organochlorine pesticide) exposure. Our data suggested that chronic exposure to endosulfan resulted in an increased oxidative stress, which was reflected by increase in LPO in erythrocytes, in a time and concentration dependent manner; antioxidative enzymes like glutathione, superoxide dismutase and catalase also fluctuated significantly in the fish blood of all the treatment groups. This type of positive correlation was also supported by many recent studies [37,135]. It is also interesting to note that the different genotoxicity tests also showed positive correlation between them. For instance, the studies performed by Mouchet showed a positive correlation between DNA strands breaks detection and micronuclei induction at most of the times [136,137]. This result was expected since the comet assay reveals primary DNA damages and the micronucleus test measures unrepaired lesions that result from the non-repaired primary DNA damages, which are likely to be inherited by subsequent generations of cells.
In recent years fish also acted as sentinel organism for screening the natural products to evaluate their pharmacological activities [138-140]. Fish, living in different aquatic environments also carry the risk of direct or indirect exposure to various xenobiotics, including pesticide residues [141]. In fish, pharmacological studies are still in preliminary stage and only few reports are available. One such report concerns about the antimutagenic and anticarcinogenic activity of chlorophyllin towards aflatoxin in rainbow trout [142]. The ameliorating effect of vitamin C, β-carotene and azadirachtin (principle compound of neem) against genotoxicity of ethyl methanesulfonate and cadmium chloride has been demonstrated in a fish, Oreochromis mossambicus [58]. Recently the antimutagenic effect of neem leaves extract in freshwater fish, Channa punctatus has been evaluated by cytogenetic tests [143].
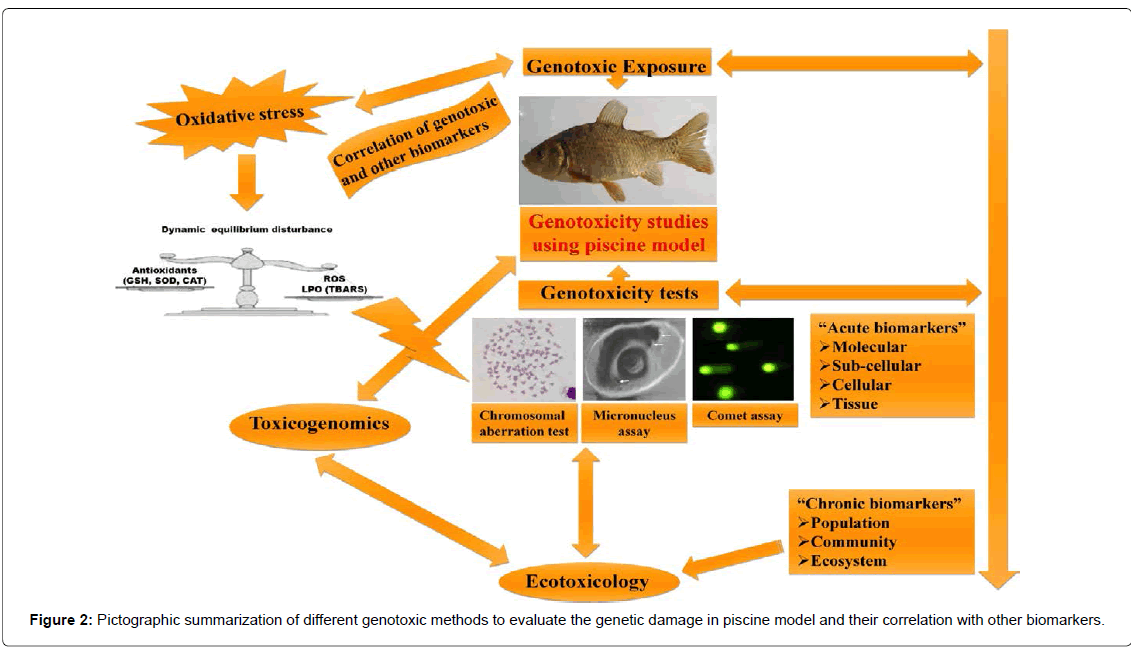
Figure 2: Pictographic summarization of different genotoxic methods to evaluate the genetic damage in piscine model and their correlation with other biomarkers.
In the recent years, as a sign of maturity, intensive attention has been devoted to the interference of non-contamination related factors, i.e., biotic and abiotic, with the genotoxicity expression. This is an important knowledge to permit an accurate assessment of the contribution genotoxins to the measured DNA damage. In this direction, hypoxia and hyperoxia, considered as critical stressors in the aquatic environment, were tested in Cyprinus carpio, revealing that both conditions increase oxidative DNA damage by 25% when compared to normoxic conditions [144]. Another study depicted that acute extreme exercise results in oxidative DNA damage in Leuciscus cephalus, suggesting that fish living in fast flowing contaminated waters are at intense risk [145]. The evaluation of the consequences of germ cell DNA damage on progeny outcomes has been regarded as a strategy to reflect potential chronic impacts of aquatic genotoxins in fish, since genetic damage in such cells, if unchecked, can be carried on to next generations. Therefore, it was concluded that a strong positive correlation exists between the DNA damage in sperm from parental fish (Salmo trutta and Salvelinus alpinus), exposed to the alkylating agent methyl methanesulfonate, and the incidence of skeletal abnormalities in the offspring; clearly indicating that DNA damage had been inherited [146]. The risk assessment of DNA-damaged germ cells in the reproduction is strictly acceptable in fish because of external fertilization or embryo development [125], since both gametes and embryos can be directly exposed to waterborne genotoxicants. This approach can represent an additional contribution to predict the impact of DNA damage on recruitment rate, progeny fitness, and thereby, on the population dynamics. Furthermore, it has been also evaluated that due to the transfer of damaged DNA to the future generation, the offsprings were subjected to elevated teratogenicity and mortality, depicting a strong relationship between genotoxicity and the declining wild populations [147].
The studies reviewed here clearly demonstrate that environmental genotoxicology holds the key to early detection and monitoring of genotoxins in aquatic environments, particularly when piscine model in taken into consideration. Fish serves as useful genetic model and the significance of assessment of genotoxicity in fish lies in the fact that higher vertebrates, including humans feeding on fish, are easily exposed to the genotoxic agents that are trapped in fish body. Ecotoxicological characteristics of fish such as its wide distribution and availability throughout the year, easy maintenance in the aquaria, cost effectiveness and commercial importance make fish an excellent model for genotoxicity studies. Furthermore, fish cells retain important traits like poikilothermic behaviour, unique xenobiotic metabolism, and low rate of repair mechanism [148], thus, making fish as sentinel organism for biomonitoring studies.
Piscine model have been used in several eukaryotic genotoxicity and mutagenicity tests, which include its use in CA test, MNT and above all comet assay. Based on the studies reviewed, MNT has been widely used to screen the genotoxic effects of the pesticides. It has several advantages over CA test such as additional detection of aneugens, technically less demanding, ease of scoring and possibility of automation and micronuclei arise from two important types of genetic damage, i.e., clastogenesis and spindle disruption. Furthermore, MNT is often more sensitive cytogenetic test with the promising results. Comet assay has broad applicability when applied to fish, providing a sensitive, reliable, rapid and versatile system for the study of environmental genotoxicity. However, one of the virtues of this assay is unquestionably its cost-effectiveness, compared to other genotoxicity techniques. Many disciplines in the aquatic genotoxicology are integrated in the task of identifying and comprehending each toxicant mode of activity in organisms, which is responsible for clinical signs of intoxication and death. The combined use of genotoxicity biomarkers, especially the comet assay with other biomarkers as well as standardization and inter-laboratory calibration are recommended to further strengthen its use in environmental assessment studies.
The elucidation of the type of DNA damage that is generated and the critical monitoring of DNA repair through lesion-specific enzymes during the comet assay, will add value to this assay in future genotoxicological studies for monitoring and risk assessment of aquatic organisms. Furthermore, it could also assist to evaluate the potential causes of their declining populations in specific environments. Despite the evidence here highlighted toward a functional association between genotoxicity measured at individual level and a negative impact at population level, so far, DNA damage measured by comet assay in fish has not succeeded to garner enough recognition to be incorporated into national and international risk assessment protocols, even though the comparison between this and other potential biomarkers as already showed higher efficiency in the distinction between impacted and reference sites. The unequivocal and convincing demonstration of its ecological relevance is probably the greatest challenge to comet assay on the next decade.
Overall, these few examples clearly illustrate that the application of the genotoxicity tests, particularly, the comet assay in genotoxicity assessment remains as purposeful as challenging. The swift integration of new methodological improvements to the comet assay protocol like DNA repair enzyme modifications shows that genotoxicologists are constantly improving approaches and protocols. Furthermore, it must be noticed, as showed by the reviewed studies, that ecotoxicology is probably one of the most diversified and complex field of research where genotoxicity assessment is surveyed as routine. As such, one may expect another decade of successful, although constantly improving, application of the genotoxicity.
In spite the fact that the new technologies are emerging, there is still no single ultimate solution for monitoring water quality. Consequently, it is important for the researchers to determine the most suitable genotoxicity tests that suits the needs and the facilities of their research institute, and then to select the available test model, as in research we have to always start with what we have.