Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2013) Volume 1, Issue 3
Bacillus bacteria have attracted the attention of scientists as promising probiotics because of their versatile antimicrobial activity and established health benefits on the host. In this study, seven Bacillus strains were identified and analyzed for antagonistic activity against broad spectrum of food borne pathogens. All strains were identified as B. subtilis, based on the results of morphological, biochemical characterization and 16S rDNA sequence analysis. B. subtilis strains demonstrated antagonistic activity against test-cultures of pathogens, including multiresistant strains. Reference Bacillus strains, derived from the commercial probiotics did not show antagonistic activity against tested strains of pathogens. Three the most active cultures were studied for production of biosurfactants. Crude biosurfactants were isolated and analyzed by oil spread test and inhibition activity against Salmonella, Shigella and Staphylococcus cultures. Biosurfactants from three tested B. subtilis strains gave positive oil spread test. Inhibition activity of biosurfactants was found only against Staphylococcus strains. Production of biosurfactants depended on the incubation conditions of Bacillus culture. Best results were obtained after cultivation of bacilli in starch broth at 30°C. The concentration of produced biosurfactant increased in time with growth of bacteria and reached the maximum at 30 hours of incubation.
Keywords: Bacillus subtilis; Antagonistic activity; Food borne pathogens; Biosurfactants
MRSA: Methicillin Resistant Staphylococcus aureus; SA: Starch Agar; TBS: Trypticase Soy Agar; NA: Nutrient Agar; CFU: Colony Forming Unit; OD600: Optical Density at a wavelength of 600 nm
Foodborne pathogens are among the most significant problems in maintaining the health of the population. In 2011 the CDC estimates that each year roughly 1 in 6 Americans (or 48 million people) gets sick, 128,000 are hospitalized, and 3,000 die of foodborne diseases. The leading causes of foodborne illnesses in the United States are Salmonella and Shigella [1,2]. Staphylococcus aureus is among top five pathogens contributing to domestically acquired foodborne illnesses. Staphylococcal food poisoning is estimated to account for 241,148 foodborne illnesses per year in the United States, according to the CDC information (http://www.cdc.gov/foodborneburden/2011-foodborneestimates.html). Foodborne illnesses are routinely treated with several classes of antibiotics. However, the use of these antibiotics has become problematic as over the years there have been numerous reports of cases of multi-antibiotic resistant food borne pathogens, worldwide [3-5]. In the United States, the proportion of methicillin-resistant S. aureus (MRSA) isolates from patients in intensive care units increased from 1992 to 2003 by 3% per year. Moreover, there is a great concern that the continued use of these drugs will result in the emergence of new resistant strains of these bacteria [6-9]. Colonization of the intestinal tract with MRSA may have important clinical implications, such as development of antibiotic-associated diarrhea, environmental dissemination, subsequent risk of infections and toxic shock syndrome [10-14]. Since foodborne infections have a dramatic impact on morbidity and mortality, particularly of infants and children, new approaches for cost effective and easy-to-deliver prophylaxis and treatment of these infections are highly desirable.
One of the growth areas in the control of foodborne infections is the use of probiotics [7]. Probiotic prophylaxes and therapies are gaining wider acceptance as more scientific data emerge regarding the interaction between pathogen and beneficial microbes in the human intestinal tract and molecular mechanisms of probiotics’ action. Probiotic bacteria which confer beneficial effect for the host and have pronounced antagonistic activity against these pathogens are expected to present a clear alternative in the prevention and treatment of foodborne infections.
Bacteria of the Bacillus genus possess a great potential as probiotic cultures. Bacillus bacteria are among the most widespread microorganisms in nature. These bacteria are known to be producers of more than 200 antibiotics. Bacillus antibiotics differ in their structure, as well as spectrum of activity [15]. Some strains of Bacillus synthesize bacteriocines, which are only effective against bacteria of the same species, others produce antibiotics against Gram-negative bacteria and still other strains have a wide spectrum of antibiotic activity (including antifungal and antiprotozoan) [16]. Thus, it is possible to find strains with unique spectrum of activity among Bacillus bacteria. The aim of this work was to isolate and characterize Bacillus strains with pronounce activity against food borne pathogens.
Bacterial strains
Seven bacterial strains (16k, M1-1, 11-89, M2-3, 101, BSB, 105), isolated from environment, were used in this study. Morphological characterization of the cultures was done with high resolution CitoViva microscope [17,18]. Gram reaction and catalase activity were analyzed in tested strains. Bacterial cultures were identified using API 50 CHB tests (bioMerieux, Marcy-l’Etoile, France). For further characterization, the 16S rRNA gene was PCR amplified using universal 16S primers that correspond to positions 0005F and 0531R. Products of sequencing reactions were analyzed with an ABI 3100- AVANT Genetic Analyzer in MIDI Labs (Newark, DE). Sequence analysis was performed using BLAST and Sherlock® DNA microbial analysis software and database.
Probiotic strains
Bacillus cultures from commercial probiotics were studied as the reference strains (Table 1).
| Bacillus strain | Probiotic | Producer |
|---|---|---|
| B. cereus IP 5832 | Bactisubtil | Cassenne Marion, Paris, France |
| B. cereus DM-423 | Cereobiogen | Keda Drugs Trade Co Ltd under Dalian university of Medical Sciences, China |
| B. clausii | Enterogermina | Sanofi -Synthelabo, Milan, Italy |
Table 1: Characterization of Bacillus probiotic strains.
Test-cultures
Test-cultures of Salmonella, Shigella and Staphylococcus were obtained from the culture collection of Auburn University (Auburn, AL). Stock cultures were maintained at -20°C in NZY medium, supplemented with 25% (v/v) glycerol.
Antagonistic activity of Bacillus strains
Activity of Bacillus strains against pathogens were studied by the method of delayed antagonism in solid nutrient medium [19]. Briefly, Bacillus strains were inoculated as a line on the surface of a nutrient media. After 72 h of growth at 30 or 37°C overnight test-cultures were inoculated as a perpendicular line to the Bacillus culture. The plates were incubated for 24 h at 37°C. The antagonistic activity was detected as a zone of pathogens’ growth inhibition. Different media were tested to assess the antagonistic activity of Bacillus strains-NZY, starch agar (SA), trypticase soy agar (TBS) and nutrient agar (NA). Starch agar composed of starch (10 g/L), peptone (5 g/L), NaCl (0.5 g/L), agar (15 g/L) was used previously for cultivation of Bacillus probiotic strain [20]. Test-cultures of pathogens were grown overnight in NZY medium at 37°C.
Biosurfactant evaluation
For preparation of the inoculum, Bacillus strains were grown in NZY medium overnight at 37°C on shaker-incubator (200 rpm). Seed cultures were inoculated into nutrition media (1 mL of overnight culture into 250-mL flack with 50 mL of tested medium). Two media were used for cultures cultivation: SA and a fermentation media for biosurfactant production by B. subtilis natto, composed of 5.0 g/L sucrose, 20.0 g/L peptone, 0.5 g/L yeast extract, 0.02 g/L MgSO4•7H2O, 1.4 g/L Na2HPO4•12H2O, 0.4g/L KH2PO4 [21]. Bacillus strains were incubated 24 hours at 30° and 37°C. After incubation the bacterial cells were precipitated by centrifugation at 10,000 x g at 4°C for 10 min. Cell free supernatant was acidified to pH 2.0 with 1N HCl and left overnight at 4°C to precipitate. The resulting precipitate was collected by centrifugation at 10,000 x g for 20 min. The supernatant was discarded and the remaining pellet was resuspended in 5 mL of methanol and left for extraction for 4 hours at room temperature. Methanol extract was centrifuged at 10,000 x g for 30 min and supernatant was transferred into a preweighed 50 mL glass and evaporated overnight under the vaccum in an exicator with silicagel. The glass was weighed again to determine the net weight of the crude biosurfactant. Obtained samples were diluted in deionized sterile water (pH 8.0) for further testing.
Biosurfactant production during Bacillus cultivation
Bacillus strain was cultivated in 250 mL flasks with 50 mL of starch medium at 30°C for 32 hours. At different time intervals, the fermentation medium was sampled for determination of biomass and biosurfactant concentration.
Surface activity of biosurfactants was measured by an oil spreading test [22,23]. Briefly, 20 μL of crude oil was added to a Petri dish (90 mm diameter) with 50 mL of distilled water to form a thin membrane. Ten microliters of sample was put onto the center of the oil membrane. The diameter of the oil-displaced circle area was measured. Each sample was tested in triplicate.
Antimicrobial activity of biosurfactants was evaluated by an agar well diffusion method [20]. Prepared suspensions of test-cultures in PBS (108 CFU/mL) were inoculated onto the surface of agar medium (100 μL of suspension on each plate). Wells (6 mm diameter) were made with a sterile cork borer. 50 μL of the test solutions were added to each well. Plates were incubated for 24 hours at 37°C. Zones of testcultures growth inhibition were measured.
Statistics
Statistical analyses (t- Test and ANOVA) were performed using Microcal™ Origin® version 6.0 (Northhampton, MA) to validate the signification of the results. The data are presented as means (± SD) of at least three replicates.
Identification of bacillus strains
The microscopic study of bacterial cultures showed these strains to be Gram-positive rods, less than 1 μm in diameter. All strains sporulated aerobically without swelling of the cell and produced catalase. These data indicated that tested strains belong to Bacillus genus. Additional testing with API 50CHB kit resulted in identification of all cultures as B. subtilis. Partial sequence of 16S rRNA gene confirmed the obtained results of biochemical identification.
Antagonistic activity
Antagonistic activity of B. subtilis cultures was tested on different nutrient media at two temperatures: 30° and 37°C. All cultures showed prominent growth on selected media at both temperature, but no antagonistic activity was indicated at 37°C. Bacillus cultures inhibited the growth of pathogenic bacteria only after growth on SA at 30°C (Table 2).
| Bacillus subtilis strain/Medium* | Zone of test-cultures growth inhibition, mm | |||||
|---|---|---|---|---|---|---|
| Staphylococcus aureus ATCC12600 | Salmonella typhimurium ATCC 13311 | S. dublin SA 2424 |
S. enteritidis | Shigella sonnei | Shigella flexneri | |
| BSB3 | ||||||
| NZY; NA; TSA | 0 | 0 | 0 | 0 | 0 | 0 |
| SA | 28.3 ± 0.3 | 29.6 ± 0.6 | 25.6 ± 0.3 | 20.3 ± 0.8 | 23.3 ± 0.8 | 21.3 ± 0.3 |
| M1-1 | ||||||
| NZY; NA; TSA | 0 | 0 | 0 | 0 | 0 | 0 |
| SA | 14.6 ± 1.4 | 6.7 ± 1.7 | 5.0 ± 0 | 7.0 ± 0 | 11.7 ± 0.3 | 11.0 ± 0.6 |
| 16k | ||||||
| NZY; NA; TSA | 0 | 0 | 0 | 0 | 0 | 0 |
| SA | 29.3 ± 0.3 | 25.6 ± 0.3 | 16.0 ± 0.6 | 22.0 ± 0.6 | 21.6 ± 0.3 | 19.6 ± 0.8 |
| 11-89 | ||||||
| NZY; NA; TSA | 0 | 0 | 0 | 0 | 0 | 0 |
| SA | 20.3 ± 0.6 | 17.7 ± 0.7 | 15.3 ± 0.3 | 13.6 ± 0.6 | 15.7 ± 0.7 | 14.6 ± 0.3 |
| M2-3 | ||||||
| NZY; NA; TSA | 0 | 0 | 0 | 0 | 0 | 0 |
| SA | 19.7 ± 0.9 | 23.7 ± 0.8 | 15.7 ± 1.2 | 21.7 ± 0.9 | 20.3 ± 0.3 | 21.0 ± 0.6 |
| 101 | ||||||
| NZY; NA; TSA | 0 | 0 | 0 | 0 | 0 | 0 |
| SA | 17.3 ± 0.7 | 6.2 ± 0.6 | 4.3 ± 0.3 | 18.6 ± 0.9 | 19.7 ± 0.3 | 15.3 ± 0.6 |
| 105 | ||||||
| NZY; NA; TSA | 0 | 0 | 0 | 0 | 0 | 0 |
| SA | 34.1 ± 1.2 | 25.7 ± 0.8 | 16.2 ± 0.3 | 23.6 ± 0.6 | 24.3 ± 0.3 | 21.2 ± 0.3 |
Table 2: Antagonistic activity of B. subtilis strains on different media.
Strains BSB, 16K and 105, showed the highest antagonistic activities, were used in further experiments with broad spectrum of Salmonella and Staphylococcus strains, including clinical isolates. Bacillus cultures from commercial probiotics were tested as reference strains. Antagonistic activity of bacilli was studied after growth on SA at 30°C. B. subtilis strains were highly effective against all tested strains of Salmonella and Staphylococcus (Table 3, Figures 1 and 2). Commercial Bacillus strains showed no antagonistic activity against test-cultures.
| # | Test -cultures | Zone of test-cultures inhibition (mm) by B. subtilis strains | ||
|---|---|---|---|---|
| 105 | BSB3 | 16 k | ||
| 1 | S. typhimurium Health 9491 | 0 | 23.8 ± 0.7 | 22.7 ± 0.3 |
| 2 | S. typhimurium DT 104 Dairy | 25.6 ± 0.3 | 25.8 ± 0.8 | 24.2 ± 0.6 |
| 3 | S. diarisonae | 25.3 ± 0.8 | 27.1 ± 0.9 | 26.3 ± 0.6 |
| 4 | S. panama SA 3583 | 22.6 ± 0.3 | 25.3 ± 0.3 | 23.1 ± 0.7 |
| 5 | S. indica SA 4401 | 23.1 ± 0.2 | 25.6 ± 0.7 | 25.2 ± 0.3 |
| 6 | S. derby SARB 10 | 30.2 ± 1.3 | 31.4 ± 0.8 | 27.3 ± 0.6 |
| 7 | S. typhimurium LT2 | 30.1 ± 0.6 | 32.3 ± 0.7 | 28.6 ± 0.3 |
| 8 | S. mission | 26.6 ± 0.3 | 25.8 ± 0.6 | 24.3 ± 0.7 |
| 9 | S. montevideo | 25.0 ± 0.0 | 25.3 ± 0.7 | 23.0 ± 0.6 |
| 10 | S. typhimurium 6787 | 22.3 ± 0.3 | 24.3 ± 0.7 | 19.7 ± 0.8 |
| 11 | S. typhimurium Heath 1390 | 21.7 ± 0.3 | 22.0 ± 0.6 | 21.7 ± 0.3 |
| 12 | S. bongori SA 4910 | 21.7 ± 0.9 | 25.0 ± 0.6 | 23.7 ± 0.3 |
| 13 | S. typhimurium Nal 1x fecal | 19.7 ± 0.8 | 20.7 ± 0.3 | 18.0 ± 1.1 |
| 14 | S. minnesota | 30.7 ± 0.3 | 31.0 ± 0.7 | 27.7 ± 0.9 |
| 15 | S. salamae SA 41106 | 10.0 ± 0.7 | 23.6 ± 0.3 | 18.7 ± 0.9 |
| 16 | S. typhimurium 520-96 | 22.3 ± 0.6 | 24.0 ± 0.6 | 21.6 ± 0.7 |
| 17 | S.Thompson 265-4 | 25.1 ± 0.3 | 26.3 ± 0.9 | 23.3 ± 0.6 |
| 18 | S. infantis SARR 27 | 31.1 ± 0.9 | 30.3 ± 0.6 | 27.6 ± 0.3 |
| 19 | S. paratyphimurium | 30.2 ± 0.3 | 30.3 ± 0.6 | 29.6 ± 0.7 |
| 20 | S. typhimurium DT 104 Swine | 21.1 ± 0.2 | 24.6 ± 0.3 | 22.6 ± 0,3 |
| 21 | S. typhimurium 9693 | 22.3 ± 0.3 | 25.7 ± 0.6 | 23.3 ± 0.1 |
Table 3: Anti-Salmonella activity of Bacillus strains.
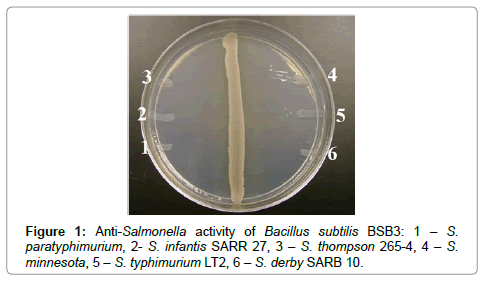
Figure 1: Anti-Salmonella activity of Bacillus subtilis BSB3: 1 – S. paratyphimurium, 2- S. infantis SARR 27, 3 – S. thompson 265-4, 4 – S. minnesota, 5 – S. typhimurium LT2, 6 – S. derby SARB 10.
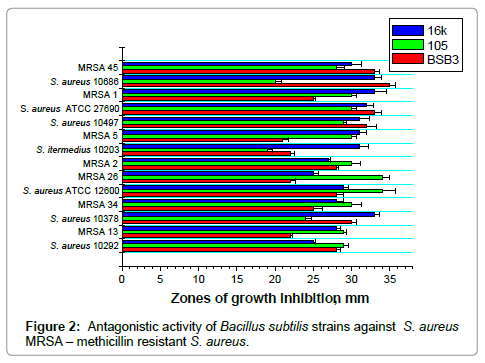
Figure 2: Antagonistic activity of Bacillus subtilis strains against S. aureus MRSA – methicillin resistant S. aureus.
Biosurfactant production
Biosurfactant production was tested in two media-starch broth and fermentation medium, used for surfactant production by B. subtilis natto [21]. B. subtilis strains were incubated in two medium at 30°C and 37°C. Production of biosurfactant was assessed by the oil spreading technique and by inhibition of test-cultures growth. The best conditions for biosurfactant production for all Bacillus cultures were cultivation in starch broth at 30°C. Results in Figure 3A indicate that oil spread test for biosurfactants, produced by B. subtilis BSB3 and 16k at 30°C gave higher results than for Bacillus cultures, grown at 37°C. Study of inhibition activity of these biosurfactants against test-cultures, showed that activity of biosurfactant from B. subtilis BSB3, cultivated at 30°C was more pronounced, than at 37°C (Table 4; Figure 3B). Biosurfactants from B. subtilis 16k, incubated at different temperatures, demonstrated more consistent results. B. subtilis 105 produced biosurfactant, as it was confirmed by the oil spreading test, but this biosurfactant had lack of inhibition activity. Biosurfactants, produced by Bacillus strains demonstrated inhibition activity only against Staphylococcus cultures. Tested cultures of Salmonella and Shigella were resistant to these biosurfactants.
| Test-cultures | Zone of test-cultures growth inhibition (mm) by biosurfactants from B.subtilis | |||||
|---|---|---|---|---|---|---|
| BSB3 (30°C)* | BSB3 (37°C)** | 16k (30°C) | 16k (37°C) | 105 (30°C) | 105 (37°C) | |
| S. aureus 10292 | 12 | 10 | 17 | 15 | 0 | 0 |
| MRSA 13 | 10 | 9 | 13 | 11 | 0 | 0 |
| S. aureus 10378 | 15 | 12 | 20 | 19 | 0 | 0 |
| MRSA 34 | 11 | 8 | 0 | 0 | 0 | 0 |
| S. aureus ATCC 12600 | 16 | 12 | 20 | 20 | 0 | 0 |
| MRSA 26 | 15 | 10 | 13 | 13 | 0 | 0 |
| MRSA 2 | 12 | 10 | 14 | 14 | 0 | 0 |
| S. itermedius 10203 | 16 | 9 | 25 | 25 | 0 | 0 |
| MRSA 5 | 12 | 9 | 15 | 14 | 0 | 0 |
| S. aureus 10497 | 13 | 11 | 17 | 17 | 0 | 0 |
| S. aureus ATCC 27690 | 13 | 11 | 15 | 15 | 0 | 0 |
| MRSA 1 | 10 | 7 | 15 | 14 | 0 | 0 |
| S. aureus 10686 | 15 | 12 | 18 | 18 | 0 | 0 |
| MRSA 45 | 13 | 10 | 18 | 16 | 0 | 0 |
Table 4: Activity of biosurfactants against Staphylococcus test-cultures.
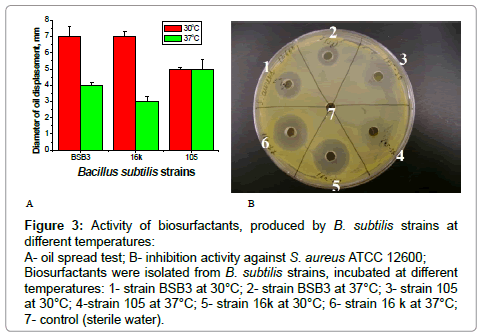
Figure 3: Activity of biosurfactants, produced by B. subtilis strains at different temperatures:
A- oil spread test; B- inhibition activity against S. aureus ATCC 12600; Biosurfactants were isolated from B. subtilis strains, incubated at different temperatures: 1- strain BSB3 at 30°C; 2- strain BSB3 at 37°C; 3- strain 105 at 30°C; 4-strain 105 at 37°C; 5- strain 16k at 30°C; 6- strain 16 k at 37°C; 7- control (sterile water).
Production of biosurfactant during B. subtilis BSB3 cultivation
Production of biosurfactant by B. subtilis BSB3 increased in time and corresponded to the bacterial growth curve. The maximum production of biosurfactant was detected at 30 hours of bacteria cultivation (Figure 4).
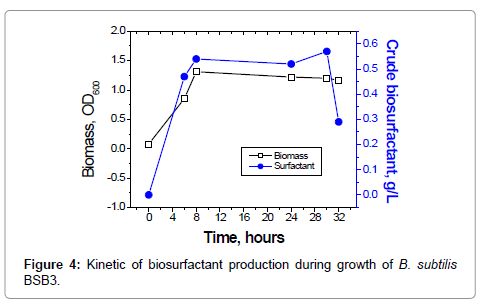
Figure 4: Kinetic of biosurfactant production during growth of B. subtilis BSB3.
Bacillus bacteria are known to be effective antagonists of different pathogens [15,24]. In recent years bacilli were extensively studied as probiotics, due to their health benefits on the host [25,26]. A search for new Bacillus strains with pronounced antagonistic activity against food borne pathogens opens up promising expectations for treatment of these infections.
In the present study, newly isolated Bacillus strains were analyzed. All strains, identified as B. subtilis, were tested for their activity against Salmonella, Shigella and Staphylococcus strains. Antagonistic activity of Bacillus strains was detected only after cultivation on starch agar at 30°C. Incubation of bacilli on starch agar at 37°C, as well as on NZY, NA and TSA at 37°C and 30°C did not result in antagonistic effect. These outcomes are in accordance with our previous findings about conditions for production of antimicrobial compounds by Bacillus cultures [27]. Three B. subtilis strains, showed the highest activity against tested pathogens, were studied with broad spectrum of Salmonella and Staphylococcus cultures, including clinical multiresistant strains. As reference strains, commercial Bacillus probiotic cultures from Bactisubtil, Cereobiogen and Enterogermina were used. None of the reference strains were active against tested pathogens. B. subtilis isolates demonstrated high activity of test-cultures’ inhibition. Antagonistic activity was detected against all strains of Salmonella and Staphylococcus, including MRSA. Inhibition of MRSA by Bacillus cultures was shown by other authors [28-30], but no anti-Salmonella effect was found in the same strains.
Bacteria of the Bacillus genus (predominantly, B. subtilis) produce various biosurfactants, which have a high potential for biotechnology and pharmacology [31]. These compounds vary in structure and spectrum of activity and usually are responsible for antimicrobial effects of Bacillus bacteria [21,32]. In our study B. subtilis strains produced biosurfactants after cultivation in starch broth at 30°C. Incubation of these cultures in fermentation medium, used for B. subtilis natto [21], resulted in lack of biosurfactants production. Presence of biosurfactant in cultivation medium was tested by the oil spread test and by inhibition activity against Salmonella, Shigella and Staphylococcus strains. It was shown elsewhere, that the oil spread test correlates with the biosurfactant production [23]. Biosurfactants from three tested B. subtilis strains gave positive oil spread test, showing the diameter of oil displacement from 3 to 7 mm. These results are in accordance with data for crude biosurfactants from B. subtilis natto [21] and from B. subtilis and B. licheniformis [33]. Inhibition activity of biosurfactants was found only against Staphylococcus strains and depended on the incubation temperature of Bacillus culture. Biosurfactant from B. subtilis BSB3, incubated at 30°C, demonstrated higher activity against Staphylococcus strains, as opposed to bacteria incubated at 37°C. Activity of biosurfactant, produced by B. subtilis 16 k did not changed with the change of incubation temperature. Biosurfactant from B. subtilis 105 showed no activity against tested pathogens. The inhibitory activity of B. subtilis may be the cumulative result of different antimicrobials, known for this bacteria [16]. Identified here biosurfactants play an important role in the anti-Staphylococcus activity for at least two B. subtilis strains - BSB3 and 16 k. Kinetic of biosurfactant production was similar to those reported for Bacillus cultures by other authors [33]. The concentration of biosurfactant increased in time with growth of bacteria and reached the maximum at 30 hours of incubation.
In the present study seven Bacillus strains were characterized for their activity against Salmonella, Shigella and Staphylococcus pathogens. Three strains showed pronounced antagonistic activity against broad spectrum of pathogenic cultures, including multiresistant strains. Inhibitory effect on S. aures and MRSA strains was caused by production of biosurfactant, identified for two B. subtilis strains.
Further study of antimicrobial compounds, produced by Bacillus bacteria, will result in better understanding of the mechanisms of antagonistic activity of bacilli and selection of new strains, promising for biotechnology and pharmacology.
Authors are grateful to members of Department of Anatomy, Physiology and Pharmacology and Department of Biological Sciences of Auburn University for support and encouragement.