Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2017) Volume 5, Issue 3
Synbiotics are a combination of probiotic bacteria and the growth promoting prebiotic ingredients that purport “synergism”. Probiotics are well known for their ability to restore a healthful balance of commensal microbes supporting the repair of hyper-permeable epithelial barriers and interfering with infection. The present research was designed to evaluate the anti-virulence role of cell-free supernatants of synbiotic cultures. The effect of prebiotics fructo-oligosaccharide, inulin and isomaltose on probiotic Lactobacillus acidophilus, Lactococcus lactis, Lactobacillus casei, Lactobacillus reuteri and Saccharomyces boulardii growth, fermentation products has been investigated. Cell-free supernatants of synbiotics cultures have been studied for their antimicrobial, antibiofilm and anti-adhesion properties. Results highlight that prebiotics can enhance the antimicrobial and anti-virulence activities of probiotics against potential pathogens Staphylococcus aureus and Escherichia coli. The present research supports, for the first time, the value of anti-virulence activity of metabolites produced by synbiotics suggesting their use as a suitable adjuvant in antibacterial treatment.
Keywords: Probiotics; Prebiotics; Anti-biofilm; Anti-adhesion; Staphylococcus aureus; Escherichia coli; Synbiotics
The word "probiotic" means “for life” (from the Greek προ β?ος, pro bios) and it was coined in 1960 to name substances produced by microorganisms that promoted the growth of other microorganisms [1]. Several definitions followed one another until the most recent proposed by Food and Agriculture Organization of the United Nations (FAO), that defines probiotics as “Live microorganisms, which, when administered in adequate amounts, confer a health benefit on the host” [2].
Interest in probiotics and probiotic-based functional foods has grown enormously during the last few years, primarily due to immense health potentials.
The action of prebiotics, which are non-digestible food ingredients such as Fructo- Oligosaccharide (FOS) and inulin, is strictly linked to probiotics since they selectively stimulate the growth of beneficial bacteria and/or suppress that of harmful bacteria in the colon, affecting beneficially the host [3].
Probiotics and prebiotics are used in synergistic combinations called synbiotics [4].
Scientific work on the properties and functionality of living microorganisms in food have suggested that probiotics play an important role in digestive and respiratory functions, suppression of mutagenesis, tumorigenesis, peroxidation, hypercholesterolemia or intestinal putrefaction [5]. Probiotics could also have a significant effect on the alleviation of infectious diseases in children and other high-risk groups. The major factors affecting the composition of the microbiota are related to changes in physiological conditions of the host (aging, stress, health status and ethnical environment), diet, medication, illness and infections. This could result in a decline of the beneficial bacteria and in an increase in potentially harmful bacteria (dysbiosis). Most experts now recognize that intestinal dysbiosis, the imbalance in the structural and/or functional properties of the gut microbiota that can disrupt host-microbe homeostasis, is integral to the pathogenesis of Inflammatory Bowel Disease. Therefore, there is great interest in modulation of the gut microbiota to provide protection from disease. Interventions that modulate the beneficial enteric microbes include diet, probiotics, prebiotics and more recently the emergence of experimental therapies such as faecal microbiota transplant or phage therapy [6].
The administration of beneficial microbes was reported to maintain a proper balance of the intestinal microbiota (eubiosis) that, actually, represents an attractive option for therapeutic or preventive applications in such medical conditions.
Probiotics showed an antimicrobial activity carried out through many mechanisms, one of which is the production of bacteriocins. Bacteriocins are a variety of antagonistic factors that include metabolic end products, antibiotic-like substances and bactericidal proteins. Bacteriocins are involved in inhibition of Gram-positive, Gram-negative bacteria, fungi and yeasts. It was shown that Lactobacillus salivarius produces ABP-118, a small bacteriocin known for its broadspectrum antimicrobial activity, since it is capable of inhibiting a number of food-borne and medically significant pathogens, including Bacillus, Listeria, Enterococcus and Staphylococcus species, without an apparent antagonistic activity towards related Lactobacillus strains, with the exception of Lactobacillus fermentum KLD [7]. Lactococcus lactis can produce nisin, a cationic peptide active against Clostridium difficile [8]. The inhibition of Listeria monocytogenes and enterohaemorrhagic E. coli in mice by the bacteriocin-producing strain Lactobacillus casei has been reported [9]. Cotter et al. suggested that, on the basis of all these properties, many bacteriocins could be of particular value in clinical settings [10].
Moreover, among all the beneficial effects of probiotics, it has been seen that Lactobacillus strains can inhibit Helicobacter pylori growth in a dose-dependent manner by mean of short chain fatty acids (SCFA) production, particularly acetic acid [11,12]. Bacteriocins and fatty acids are just two examples of pharmabiotic substances which may contribute to probiotic functionality within the mammalian gastrointestinal tract [13].
Moreover, Lactobacilli are known to produce biosurfactants with antimicrobial activity against several pathogens in the intestinal tract interfering with biofilm formation and adhesion to the epithelial cells surfaces [14].
This study was specifically undertaken with the objective of assessing the health benefits of prebiotics (FOS, inulin and isomaltose) on the growth, antimicrobial and anti-adhesion properties of five potential probiotics (L. acidophilusi , L. lactis , L. casei , L. reuteri and S. boulardii ). Cell-free supernatants of probiotics (CFS) and cell-free supernatants of synbiotics (prebiotic-probiotics) (CFS-S) cultures have been tested in in vitro models with the aim to evaluate the kinetic growth, the SCFA production, antimicrobial, anti-biofilm and antiadhesion activities.
Bacterial strain and culture conditions
Lactobacillus acidophilus ATCC 4356, Lactococcus lactis ATCC 11454, Lactobacillus casei ATCC 334, Lactobacillus reuteri ATCC 55148 and Saccharomyces boulardii ATCC MYA- 796 (Sb48) were purchased from the American Type Culture Collection-ATCC, LGC Standards S.r.L., Milan, Italy. One day before the experiment, a colony of each strain has been isolated from each culture and inoculated, separately, onto a 7 mL of fresh De Man, Rogosa and Sharpe (MRS) broth (Sigma-Aldrich, Ottawa, Canada, USA). Microbial suspensions have been incubated for 24 h at 37°C.
Staphylococcus aureus ATCC 29213 and Escherichia coli ATCC PTA-7296 (American Type Culture Collection-ATCC, LGC Standards S.r.L., Milan, Italy) were used to test antimicrobial and antibiofilm activity of our probiotic strains. Bacteria were initially streaked from -80°C glycerol stock onto a Muller Hinton Agar (MHA, Sigma-Aldrich, Ottawa, Canada, USA) plate and a fresh single colony was inoculated into 15 ml of Muller Hinton Broth (MHB, Sigma-Aldrich, Ottawa, Canada, USA) and cultured overnight at 37°C. Cell growth was determined by measuring optical density at 600 nm using a spectrophotometer (Infinite® m200 PRO reader, Tecan Group Ltd., Männedorf, Switzerland Tecan).
Kinetic growth of probiotics in prebiotics-containing media
The inocula have been adjusted to a final concentration of 105 CFU/mL in simple MRS or media containing 1% w/v of different prebiotics (FOS, inulin, isomaltose alone or in combination). FOS was purchased from Natur-BioCare (San Pietro di Legnano, Italy), inulin and isomaltose from Bio-line Laboratories (Canaro, Italy).
The cultures have been incubated at 37°C in Infinite® m200 PRO reader (Tecan) and each sample has been read at 600 nm every 60 minutes for 24 h.
CFS production
106 CFU/mL of probiotics were inoculated in a volume of 15 mL of simple MRS and MRS containing FOS, inulin and isomaltose alone or in combination and incubated for 24 h at 37°C with periodic mixing. After incubation, samples were centrifuged at 3000 g for 10 minutes and supernatants were sterilized through a 0.22 filter. CFS were stored at -20°C until use.
Determination of SCFA produced by probiotics
106 CFU/mL of probiotics were inoculated in a volume of 15 mL of simple MRS and MRS containing FOS, inulin and isomaltose alone or in combination and incubated for 24 h at 37°C with periodic mixing. To test acetic acid production, probiotics were grown in 15 mL of Roswell Park Memorial Institute 1640 medium [RPMI 1640] in presence or absence of different prebiotics in order to avoid the interference observed for MRS in gas chromatography (GC) analysis. After incubation, samples were centrifuged at 3000 g for 10 minutes and supernatants were recovered in 15 mL tubes. CFS were acidified with 20% sulphuric acid and filtered using 0.22 μm cellulose filters (Phenomenex Italia, Castel Maggiore, Italy). Standards (acetic acid, propionic, butyric and isobutyric acids) and sulphuric acid, used to acidify our samples, were obtained from Fluka (Sigma-Aldrich, Ottawa, Canada, USA). SCFA have been determined using GC (Trace- GC Thermo) equipped with a flame ionisation detector and a VF-WAX column (30 m length, 0.25 mm internal diameter and 0.25 μm film thickness).
Minimal Inhibitory Concentration (MIC) assay
The Minimal Inhibitory Concentration (MIC) was determined by micro-broth dilution method according to the Clinical and Laboratory Standards Institute/National Committee for Clinical Laboratory Standards (CLSI/NCCLS) Approved Standard M100-S21 [15]. Gentamicin (Sigma-Aldrich, Ottawa, Canada, USA) solution (2 mg/ml) was prepared by dissolving the agent in endotoxin free water. Supernatants of probiotics grown in presence of prebiotics were recovered as described above. Briefly, to determine the MIC of probiotics supernatants against S. aureus and E. coli , supernatants were serially diluted 1:2 in U bottom 96-well plates starting from a dilution of 1:2 up to 1:4096. Gentamicin has been used as a control. The plates were incubated at 37°C for 24 h. The MIC of each probiotic supernatants was defined as the lowest amount of supernatant able to inhibit the visible growth of the microorganisms.
Biofilm formation
The in vitro static biofilm assay was performed using a 96-well microtiter plate as previously described with some modification [16,17]. To grow biofilms, overnight cultures of S. aureus and E. coli were diluted 1:100 into 15 ml of MHB supplemented with 2% w/v sucrose (Sigma-Aldrich, Ottawa, Canada, USA), in presence or in absence of different probiotic supernatants. Cultures were incubated at 37°C for 24 h under static conditions. After incubation, the biofilm developed in each well was washed twice with 200 μL of distilled water and then dried for 45 min. In each well, 100 μL of 0.4% crystal violet (Merck KGaA, Frankfurter Strasse 250, 64293 Darmstadt, Germany) were added for 30-45 min. After this procedure, the wells were washed four times with distilled water and immediately discoloured with 200 μL of 95% ethanol (Sigma-Aldrich, Ottawa, Canada, USA). After 45 minutes, 100 μL of discoloured solution was transferred to a well of a new plate and the crystal violet was measured at 570 nm in a microplate reader. The amount of biofilm formed was measured comparing the absorbance values of the CFS or CFS-S-treated wells versus untreated control wells. Biofilm formation bioassays were performed in quadruplicate in at least two individual experiments for each concentration.
Propagation and maintenance of cell lines
HT-29 human mucus secreting adenocarcinoma cell line (ATCC HTB-38) was cultured in Roswell Park Memorial Institute 1640 medium [RPMI 1640] (Sigma-Aldrich, Ottawa, Canada, USA), supplemented with 10% (v/v) heat-inactivated (56°C/30 min) fetal bovine serum (FBS, Sigma), 100 U penicillin/mL and 100 μg streptomycin/mL (cRPMI) in 25 cm2 culture flask at 37°C in an atmosphere of 5% CO2. For the adhesion assay, cells were seeded into 96-well culture plates at a concentration of 2 ×105 cells per well and incubated until monolayer formation.
Bacteria labelling
For adhesion assay, cells were harvested by centrifugation, washed three times in 0.01 M phosphate buffer saline (PBS, pH 7.2) containing 0.15 M NaCl (PBS) and then heat-killed at 65°C for 1 h. Cells were stored at 4°C in PBS containing 0.05% sodium azide. After heatinginactivation, killing action was tested by seeding 100 μl of bacterial suspension in MHA agar. To label cells with fluorescein, cells were resuspended to 2 × 108 CFU/ml in either 0.01 mg of Fluorescein isothiocyanate (FITC)/ml of PBS (suspension assays). After incubation for 15 min at room temperature in the dark, FITC-labeled bacteria were washed twice with Hanks balanced salt solution containing 0.25% Bovine Serum Albumin (HBSA) and resuspended at the concentration of 5 × 107 CFU/ml to the appropriate concentration in HBSA.
Adhesion assay
In vitro adhesion assay was performed as described by Bianchi et al. [18] with some modifications. The HT29 cell monolayer was washed twice with 1 ml of PBS for removing red phenol of the medium. Suspension of S. aureus or E. coli labelled with FITC was added to each well (1 × 106 CFU/20 μl) for 30 minutes at 37°C in an atmosphere of 5% CO2. After incubation monolayers with adhered bacteria are washed twice with 200 μl of PBS at room temperature. After adding 100 μl of PBS in each well the plate was read in a microplate reader fluorometer at an Excitation of 485 nm and an Emission of 530 nm (Tecan). A calibration curve was obtained for each bacterium, two fold dilutions of the working bacterial suspension starting from 106 bacteria/ml were prepared and added to monolayer before reading by fluorimeter (TECAN). The fluorescence intensity of supernatants was plotted against the corresponding CFU. Three replicates were run for each strain in two different experiments.
Statistical analysis
Results are given as mean ± standard deviation (SD). Significance was tested by means of a Students two-tailed t test. P< 0.05 was considered significant.
Kinetics growth of probiotics in presence of different prebiotics
The kinetics growth of probiotics has been evaluated to determine the most efficient prebiotics stimulating growth and metabolism of selected microorganisms (Figure 1). FOS, inulin and isomaltose, alone and in combination stimulated significantly the growth of L. acidophilus starting from the 16th hour of incubation with respect to MRS medium alone. L. lactis showed an increased growth in medium containing isomaltose or the combination of prebiotics after 24 h of incubation with respect the control medium.
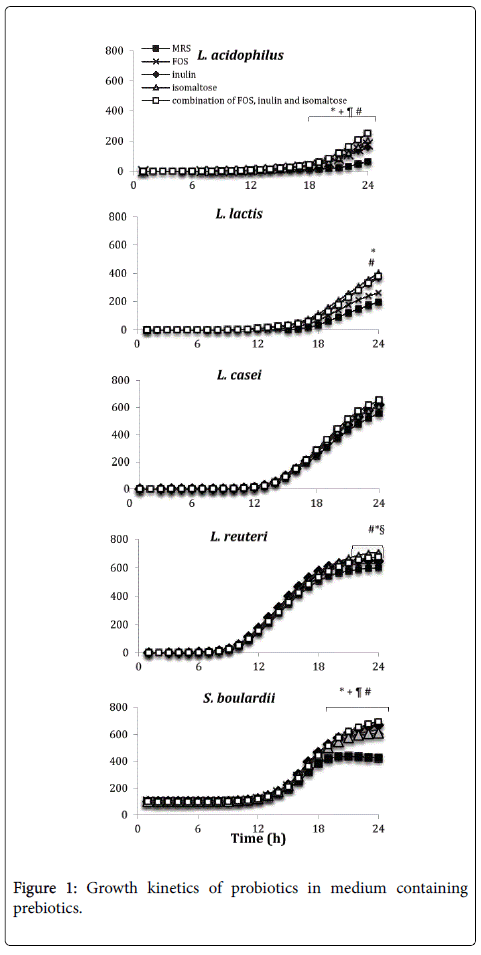
Figure 1: Growth kinetics of probiotics in medium containing prebiotics.
A significant increase of L. reuteri growth has been observed in presence of isomaltose and medium containing the prebioticscombination. Moreover, the growth of L. reuteri in presence of prebiotic-combination resulted significantly higher than in presence of MRS containing FOS. The growth of S. boulardii , after 18 h of incubation, was significantly increased in each medium containing prebiotics when compared with the control MRS. No different kinetics have been observed when L. casei has been incubated in different media. L. casei kinetic was not affected by prebiotics addition. In general, it can be affirmed that probiotic growth is improved by the presence of a combination of prebiotics rather than by individual oligosaccharides.
SCFA production
SCFA are produced by bacterial fermentation of carbohydrates, which occurs in the colon. Colonic-derived SCFA provide from 5 up to 15% of the total energy requirement by humans, depending on nutrient intake and the composition of the microbiota [19]. The major products of the saccharolytic fermentation are butyrate (C4) and isobutyric (iC4), propionate (C3) and acetate (C2). They are involved in several physiological processes such as blood pressure regulation, inflammation, lipid metabolism, mineral and ammonia absorption [20,21]. They are readily absorbed by large intestinal mucosal cells and act as energy sources for colonocytes.
In this study, we determined the concentration of C2, C3, C4 and iC4 SCFA in CFS and CFS- S by GC analysis (Figure 2).
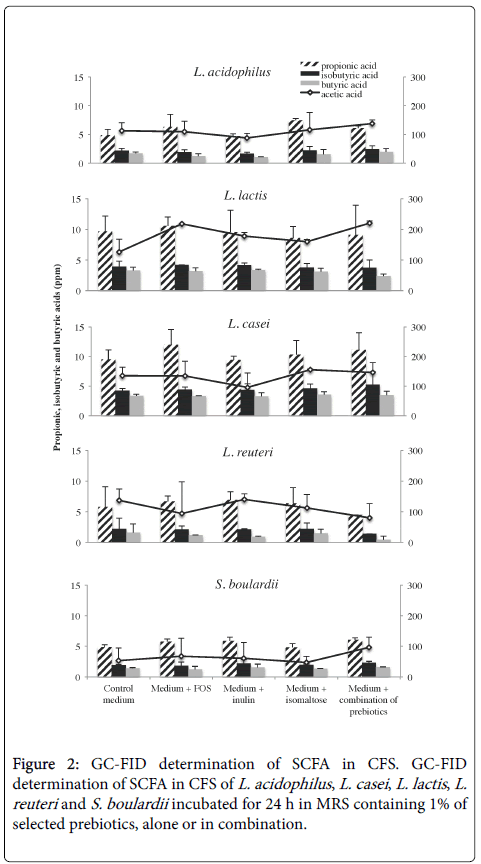
Figure 2: GC-FID determination of SCFA in CFS. GC-FID determination of SCFA in CFS of L. acidophilus , L. casei , L. lactis , L. reuteri and S. boulardii incubated for 24 h in MRS containing 1% of selected prebiotics, alone or in combination.
A higher production of C2 in supernatants of L. lactis (~200 ppm) when compared with other bacteria (~100 ppm) was detected. CFS of S. boulardii contained a lower concentration of C2 (~60 ppm). The combination of FOS, inulin and isomaltose resulted the best condition to stimulate C2 production for all probiotics tested, with the exception of L. reuteri . As regard the production of C3, iC4 and C4, the concentration, determined in CFS of probiotics, was around 12 ppm, 2.5 ppm and 1.5 ppm respectively. Instead, the production of C3, iC4 and C4 in L. lactis and L. casei CFS was higher than that observed in other CFS. Although a difference in the induction of SCFA production by different probiotics has been observed, prebiotic addition did not affect the SCFA release.
Antimicrobial activity
Antimicrobial activity of supernatants of probiotics growth in absence or in presence of prebiotics versus S. aureus and E. coli is shown in Figure 3. The MIC value was normalized for the number of bacteria; the MIC is expressed as μl of supernatants of a bacterial culture at a concentration of 108 bacteria/ml. The positive control for both Gram-positive S. aureus and Gram-negative E. coli was gentamicin with MIC values of 0.97 and 0.48 μg/ml respectively. All probiotics grown in MRS showed an antimicrobial activity against S. aureus , the addition of all prebiotics tested, alone or in combination, to the culture of L. acidophilusi , L. lactis and L. casei reduced significantly the MIC value when compared with MRS medium. As regard L. reuteri CFS, only the prebiotic-combination improved the antimicrobial activity. Instead, the anti-Staphylococcus property of S. boulardii was lower than that observed for the other probiotics and the addition of prebiotics did not affect the activity. Generally, the antimicrobial activity exhibited by probiotics versus Gram-positive bacteria was different from that observed for Gram-negative bacteria. In fact, only L. acidophilusi and L. lactis CFS showed a comparable antimicrobial activity against E. coli with respect to S. aureus . L. casei , L. reuteri and S. boulardii CFS showed high MIC values (70, 50 and 40 μl, respectively).
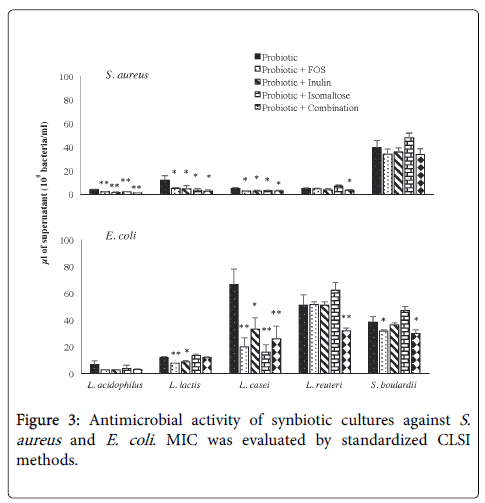
Figure 3: Antimicrobial activity of synbiotic cultures against S. aureus and E. coli . MIC was evaluated by standardized CLSI methods.
The addition of prebiotics to L. acidophilusi cultures did not influence antimicrobial activity, while FOS and inulin improved L. lactis effect. As observed for S. aureus , the antimicrobial activity of L. casei has been increased by the addition of prebiotics alone or in combination whereas for L. reuteri only the addition of prebioticscombination was effective. Finally, for S. boulardii , unlike that observed for S. aureus , the addition of FOS or prebiotic combination has improved the performance of the yeast. In general, the prebiotics tested improve the antimicrobial activity of probiotics against the Gram positive S. aureus to greater extent than versus the Gram negative E. coli , suggesting that the metabolites involved in antimicrobial activity are different or act differently.
Antibiofilm activity
To analyse in depth the antimicrobial properties of CFS and CFS-S, we examined the ability of S. aureus and E. coli to form biofilm in the absence or presence of different supernatants (Figure 4). Biofilm formation was measured by determining the mass of biofilm using crystal violet staining. Biofilms were grown in static conditions in the presence of CFS and CFS-S at concentrations lower than MIC. In particular, we used a concentration of 0.5 MIC and 0.1 MIC for the bacterial strains tested in order to find an effect on biofilm formation independent on microbiostatic or microbicidal properties.
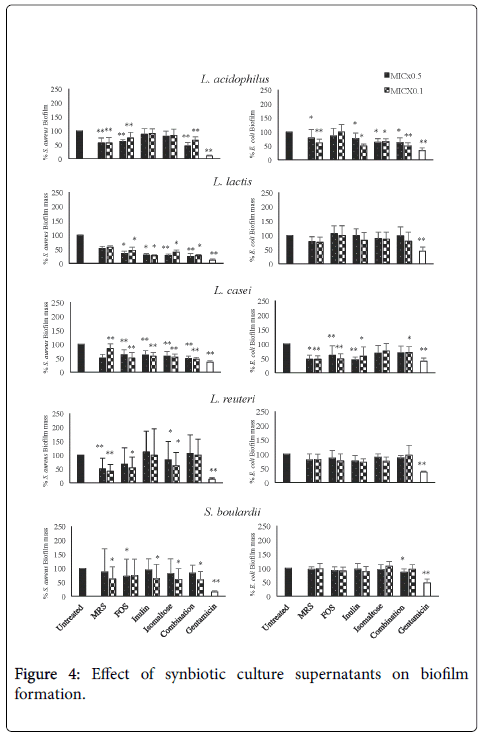
Figure 4: Effect of synbiotic culture supernatants on biofilm formation.
Supernatant of L. acidophilusi , grown in MRS, was able to reduce the biofilm mass of both S. aureus and E. coli . The addition of FOS, alone or in combination, did not affect the anti-biofilm property of L. acidophilus metabolites, whereas the adding of inulin and isomaltose abrogated the anti-biofilm activity against S. aureus . On the contrary, FOS abrogated anti- biofilm activity against E. coli while inulin and isomaltose did not alter the observed antimicrobial effect.
These data suggest that prebiotics may interfere with the production of metabolites with antimicrobial activity produced by L. acidophilusi .
L. lactis supernatant was not able to reduce significantly the biofilm mass against the Gram- positive bacteria S. aureus and Gram-negative E. coli . However, the addition of prebiotics, alone or in combination, has prompted the L. lactis to produce metabolite(s) with anti-biofilm activity against S. aureus but not against E. coli . Supernatants of L. casei was able to reduce the biofilm mass of S. aureus at a concentration of 0.1 MIC but not at 0.5 MIC. The addition of prebiotics, alone or in combination, did not affect the antibiofilm properties of the L. casei metabolites obtained with 0.1 MIC, but it decrease the biofilm mass observed with 0.5 MIC. L. casei products, obtained in absence or in presence of prebiotics, showed an antibiofilm activity against E. coli . The prebiotic isomaltose seems down-regulate the antimicrobial activity observed, this effect is probably due to its interference with the bacterial metabolism.
Metabolite(s) produced by L. reuteri showed antibiofilm activity only versus S. aureus . The addition of prebiotics did not affect the activity against E. coli and even worsened the activity against S. aureus .
S. boulardii metabolites were able to reduce only the biofilm mass of S. aureus at 0.1 MIC and prebiotics did not affect this property. No activity was observed against E. coli
In parallel experiments, the effect of MRS plus prebiotics (alone or in combination) on S. aureus and E. coli biofilm formation was analyzed. Prebiotics did not affect the mass of the biofilm (data not shown) suggesting that the inhibition of biofilm formation observed was due to probiotic metabolites produced in presence of prebiotics.
Anti-adhesion activity
To evaluate the influence of prebiotics on the production of probiotic metabolites able to interfere with the pathogen colonization, we analysed the adhesion of S. aureus and E. coli to the intestinal cell line HT29 in presence of CFS (Figure 5). Supernatants of L. acidophilus in absence or in presence of FOS and isomaltose were able to down-regulate the adhesion of S. aureus to HT-29 monolayer. Indeed isomaltose, alone or in combination with FOS and inulin, reduced the adhesion of Gram negative E. coli . CFS of L. lactis was not able to affect the adhesion of both potential pathogens with the exception of prebiotic combination on S. aureus . CFS of L. casei cultured in media containing FOS and isomaltose alone or in combination showed anti-adhesive effect on S. aureus . Interestingly, CFS of L. casei in presence of inulin decreased significantly the adhesion of E. coli respect to untreated monolayers and versus epithelial cells treated with L. casei metabolites produced in absence of prebiotics. Inulin addition to the culture medium of L. reuteri was also able to reduce the adhesion of S. aureus to HT-29 respect to control (RPMI 1640) and L. reuteri CFS treated monolayers. The same effect was observe for E. coli adhesion when HT-29 were treated with CFS of L. reuteri grown in presence of the combination of prebiotics. No effect on the microbial adhesion has been observed with S. boulardii CFS. The metabolites of probiotics grown in RPMI 1640, by themselves, are unable to regulate microbial adhesion to epithelial cells. We observed that, the addition of prebiotics has modified the metabolite profile of probiotics, prompting the anti-adhesion activity, which is an important virulence factor.
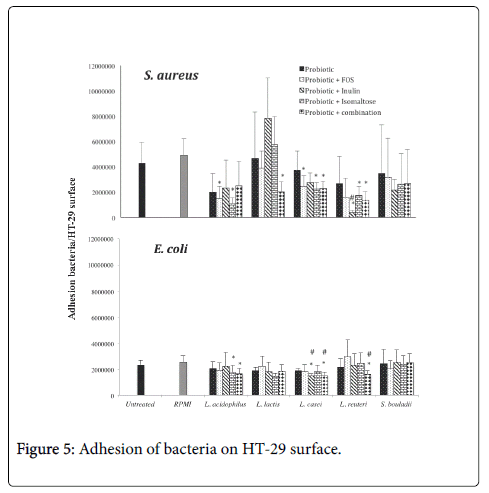
Figure 5: Adhesion of bacteria on HT-29 surface.
In recent years, there have been considerable interests in the use of probiotic live cells for nutritional and therapeutic purposes. Nevertheless, some limitations, such as survival of live cell during the gastrointestinal-transit and their effective delivery to target tissues upon ingestion, have to be considered. Several attempts have been made to overcome these limitations such as their microencapsulation, spray drying and lyophilisation.
In this work, CFS of probiotics and CFS-S, as a suitable alternative strategy for the preparation of probiotic-products, were investigated for their anti-microbial, antibiofilm activities and anti-adhesion properties.
The main aims of this study was to find out the beneficial activity of metabolic products secreted by CFS and CFS-S, moreover, to evaluate the effect of selected prebiotics on kinetic growth of our bacteria strains.
It is well known that some oligosaccharides could promote selectively the growth of lactobacilli and bifidobacteria in the gastrointestinal tract.
To date, only a few carbohydrates have been documented as prebiotics, namely β-galacto- oligosaccharides (GOS), lactulose, fructo-oligosaccharides and inulin [22]. Several candidate prebiotics have been proposed, but there is a need for additional studies that document their selective utilization by beneficial microbes.
The stimulation of growth rate of probiotics by prebiotics was the main goal of this first analysis. Our results clearly indicate that cell growth is enhanced with the addition of polysaccharides FOS, inulin and isomaltose. Belenguer et al. proposed FOS and inulin as prebiotics that selectively stimulate bacterial growth [23].
In our knowledge there are not studies concerning the use of isomaltose in combination with probiotics. However, our data reveal that isomaltose promotes the growth of L. acidophilusi , L. lactis , L.reuteri and S. boulardii , suggesting a potential prebiotic effect. This result is in line with the opinion of Roberfroid et al. which proposed to expand the original idea of the prebiotic concept, that should be translated in ‘prebiotic effects’, defined as: ‘The selective stimulation of growth and/or activity (ies) of one or a limited number of microbial genus (era)/species in the gut microbiota that confer (s) health benefits to the host [22].
One of the their major metabolic functions of probiotics is the fermentation of undigested carbohydrates and proteins and the relative production of organic acids and molecules, which may have important intestinal trophic effects and represent an additional energy source for the host, as well [24]. Therefore, we analysed the capacity of chosen probiotics to produce SCFA in presence of stimulating prebiotics FOS, inulin and isomaltose to the medium. A different response to prebiotics and a higher production of C2 with respect to C3, iC4 and C4 has been observed. Metabolic products, including SCFA, produced from dietary prebiotics by one bacterial species may then provide substrates to support growth of other populations and this is termed cross-feeding.
Specifically, the cross-feeding clarifies the conversion of C2, the major fermentation product, to C3 and C4 by the intestinal microbiota [24] (Figure 6).
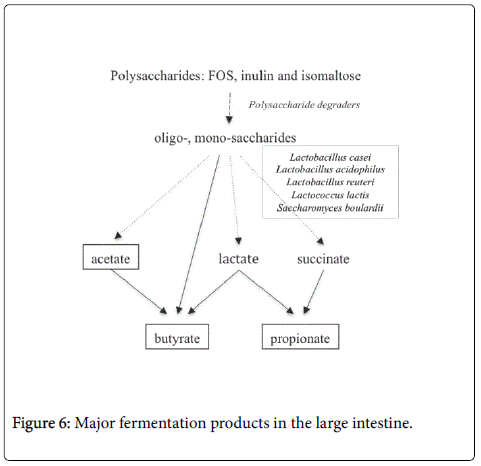
Figure 6: Major fermentation products in the large intestine.
SCFA are considered pivotal in the interaction between gut microbiota and the human host. An additional property of SCFA is the antimicrobial effect on pathogenic microorganism. Fatty acids can inhibit the growth of bacteria, protozoans, viruses and fungi. Fatty acid sensitivity is a characteristic of Gram-positive bacteria respect to Gram-negative species [25], confirming the better antimicrobial and antibiofilm activity observed for S. aureus suggesting that among active metabolites SCFA could play a role.
Indeed, pathogenic E. coli O157:H7 regulate flagellar expression and motility based on relative composition and concentration of SFCA by an unknown mechanism [26]. Probiotics CFS could also contain bacteriocins that are involved in their antimicrobial activity, in particular subtilosin by Bacillus subtilis prevents biofilm formation by inhibiting bacterial quorum sensing [27]. L. acidophilusi is already known for its antimicrobial activity against S. aureus and E. coli [28], but the antibiofilm property has been demonstrated only in co-culture with S. aureus [29]. Unlike S. boulardii , the antimicrobial effects of L. acidophilus , L. lactis , L. casei and L. reuteri , mediated by direct cell competitive exclusion, as well as the production of acids or bacteriocin-like inhibitors against S. aureus , are already extensively studied [30]. The capacity of L. acidophilusi CFS to inhibit the production of the virulence factor lipase by planktonic and sessile S. aureus has been reported, but no information are available about the biofilm mass. Esopolysaccharide produced by L. acidophilusi showed anti-biofilm activity against a wide range of Gram-positive and Gramnegative bacteria by affecting the expression of genes involved in chemotaxis, auto-aggregation and co-aggregation [31]. Nisin A, a class I bacteriocin, is produced by L. lactis and it is able to inhibit and kill strains of methicillin- resistant Staphylococcus aureus [32].
Moreover, Meng et al. demonstrated that L. acidophilusi strains possessed murein hydrolase activities that are sublethal to E. coli cells [33]. Lipocalin-2 (Lcn2), a multifunctional innate immune protein derived from L. lactis inhibited the growth of E. coli and reduced the bioactivity of enterobactin (E. coli -derived siderophore) in vitro [34].
In contrast with our study, Sharma et al. observed that L. casei CFS does not exhibit inhibitory activity against four pathogens, namely S. aureus , L. monocytogenes , E. coli , and Klebsiella pneumoniae [35]. For L. reuteri , instead, a strong antibacterial potential was observed against E. coli by means of organic acids, ethanol, and reuterin production [36]. Concerning S. boulardii , the antimicrobial capacity has been studied in terms of adhesion of pathogenic bacteria to its surface as a mechanism for trapping [37].
In our knowledge, it is the first time that CFS of probiotics are used to inhibit E. coli biofilm formation. Gómez et al. used preformed probiotic biofilms (including L. lactis and L. casei ) as alternative approach to contrast the formation of pathogenic biofilms (L. monocytogenes , E. coli O157:H7 and Salmonella typhimurium ) in food industries [38].
We confirm that all probiotic CFS tested were able to reduce the biofilm mass of S. aureus at a concentration lower than MIC. The addition of prebiotics did not affect the anti-biofilm activity observed with the exception of L. casei CFS-S whose showed anti-biofilm property only in presence of prebiotics. A possible molecule involved could be biosurfactants produced by L. casei and L. reuteri [39]. Biosurfactants from L. casei strains exhibited considerable antioxidant and anti-proliferative potencies and were able to inhibit biofilm formation of oral S. aureus strains [40]. Only L. acidophilusi and L. casei were able to reduce the biofilm mass of Gram-negative E. coli . As regard the yeast S. boulardii , its antibiofilm activity is not yet demonstrated. The reduction of biofilm mass of S. aureus observed in our experimental condition is probably related to molecule (s) involved in quorum sensing since the inhibition was significant only at 0.1 MIC and not at 0.5 MIC. This phenomenon could be related to the sigmoidal dependence of the biological effect of metabolites-receptor interaction.
Adherence of bacteria to the epithelium is an important factor of virulence as it is the first step in the colonization of a tissue. The antiadhesion activity of probiotics is often associated with their competition with pathogens for binding sites. To date, there are few studies that analyse the anti-adhesion activity of probiotic CFS. A study published in 2012 has demonstrated the anti-adhesion activity of Lactobacillus gasseri and Lactobacillus plantarum supernatants versus Helicobacter pylori on Caco 2 cells [41].
In our knowledge, the present study analysed for the first time the ability of metabolites produced by synbiotics to inhibit the adhesion of Gram positive and Gram negative bacteria to intestinal epithelial cells.
In conclusion, this study highlights the possible use of synbiotics as adjuvants in antimicrobial strategies. Future work will be focused on the mechanisms of action of synbiotics and on understanding which metabolite(s) are involved in the antibiofilm and anti-adhesion activities.