Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2016) Volume 5, Issue 5
Juglans regia L. (walnut) is a plant traditionally used for the treatment of diarrhea. We investigated the antidiarrheal effects of a hydroethanolic leaf extract (HLE) of J. regia against castor oil-induced diarrhea in rats after screening phenolic compounds in the extract. J. regia HLE (0.5, 1, and 2 g/kg by gavage) produced a dose-dependent delay in the onset of diarrhea compared to the vehicle. It also significantly reduced the number of diarrheal feces, fresh weight and water content of the feces, body weight loss and pain at the two highest doses. Loperamide was effective in inhibiting diarrhea and body weight loss but failed to reduce pain. Our findings show that J. regia HLE has a beneficial effect on diarrhea symptoms and associated pain suggesting that it could be an interesting alternative to standard anti-motility drugs. Flavonoids and hydroxycinnamic acids detected in the extract might explain the effects observed.
Keywords: Juglans regia L.; Walnut extract; Diarrhea; Castor oil; Rats
HLE: Hydroethanolic Leaf Extract; SEM: Standard Error of the Mean; UHPLC: Ultra-high Performance Liquid Chromatography.
Diarrheal disease is a significant cause of morbidity and mortality worldwide, especially in children [1]. Although relatively few patients die from diarrhea in industrialized countries, it remains an important cause of morbidity associated with substantial health care costs [1]. Diarrhea can be defined by increased stool frequency, liquidity or volume and may be accompanied by tenesmus, fever and abdominal pain [2]. Treatment of diarrhea is generally non-specific and is usually aimed at reducing the discomfort and inconvenience of frequent bowel movements. Oral rehydration therapy remains the major treatment for diarrhea although it does not reduce the volume or duration of diarrhea [3]. The major anti-motility drugs for acute diarrhea are loperamide and diphenoxylate [4]. Post-treatment constipation is frequent with loperamide. In addition, it is not recommended for children due to the potential for central nervous system side effects and the possibility of respiratory depression [5]. Diphenoxylate contains atropine, which has no antidiarrheal effectiveness and may produce objectionable side effects. Treatment with antibiotics is controversial unless diarrhea is severe or due to cholera [5]. Side effects and expenses associated with the use of standard medicines have generated in the last decades renewed interest in medicinal plants [3].
Walnut (Juglans regia L.) is a plant of the family Juglandaceae and is the most widespread tree nut in the world [6]. The useful parts of walnut tree are leaves, second peel and fleshy part of green fruit and its wood. J. regia leaves have been widely used in traditional medicines because of their anti-microbial, anti-helminthic, astringent, antidiarrheal and hypoglycemic activity [6]. It was recently demonstrated that the aqueous and ethanolic leaf extracts possess peripheral analgesic properties and activity against acute and chronic inflammation [7].
The aim of the present study was to provide support for the traditional use of J. regia as an anti-diarrheal agent. Anti-diarrheal properties of a hydroethanolic leaf extract (HLE) of J. regia were evaluated by using the castor oil-induced diarrhea model in rats after characterization of the extract.
Preparation of the HLE of J. regia
The HLE of J. regia was obtained according to patented process WO2001056584A1. J. regia fresh leaves were extracted with 20 to 70% (v/v) ethanolic leaching. Extracts were then mixed and concentrated under reduced pressure to evaporate ethanol (100 mbar absolute pressure, controlled temperature at 40-50°C). Glycerol was added to the aqueous solution to obtain a dry material with a final concentration of 2.1% (w/w). The HLE of J. regia (marketed under the brand name EPS Walnut, PhytoPrevent, France) was stored at room temperature before use.
Analysis of the HLE of J. regia
Chromatographic analyses using ultra-high performance liquid chromatography (UHPLC) of the extract were performed on a Ultimate 3000 RSLC UHPLC system (Thermo Fisher Scientific Inc., MA, USA) coupled to a quaternary rapid separation pump (Ultimate autosampler) and a rapid separation diode array detector. Phenolic compounds were separated on a Kinetex EVO C18 column (2.1 × 100 mm; 1.7 μm; Phenomenex, CA, USA), which was controlled at 30°C. The mobile phase was a mixture of 0.1% (v/v) formic acid in water (phase A) and 0.1% (v/v) formic acid in acetonitrile (phase B). The gradient of phase A was 100%-80% (0-10 min), 80%-73% (10-35 min), 73%-0% (35-40 min) and then was held at 0%. The flow rate was 0.8 mL/min, and the injection volume was 5 μL. The UHPLC system was connected to an Orbitrap (Thermo Fisher Scientific Inc., MA, USA) mass spectrometer, operated in negative electrospray ionization mode. Source operating conditions were: 3 KV spray voltage; 320°C heated capillary temperature; 400°C auxiliary gas temperature; sheath, sweep and auxiliary gas (nitrogen) flow rate 50, 10 and 2 arbitrary units, respectively; and collision cell voltage between 10 and 50 eV. Full scan data were obtained at a resolution of 70,000 whereas MS2 data were obtained at a resolution of 17,500. Data were processed using Xcalibur software (Thermo Fisher Scientific Inc., MA, USA).
Compounds present in the HLE were characterized according to their retention times, mass spectral data and comparison with authentic standards when available or with the related literature [8- 10]. The amount of flavonoids was estimated according to the French Pharmacopoeia [11].
Animals
Male Wistar rats (Charles River Laboratories, L’Arbresle, France) weighing 225-250 g were housed in groups of two per cage in a room with 12-hour inverted light/dark cycle (9:00 pm/9:00 am) and controlled temperature (22 ± 2°C) and hygrometry (50 ± 20%). Food (M20, SDS Dietex, Argenteuil, France) and water were available ad libidum. Animal care protocols were used in accordance with guidelines of the European Communities Directive 2010/63/EU and the ASAB Ethical Committee. The study received the approval by the French Ministry of Higher Education and Research (Agreement no. 1012.01 from March 2015).
Castor oil-induced diarrhea in rats
Castor oil-induced diarrhea was performed according to the method developed by Karim and colleagues [12]. After 12 days of acclimatization, the rats were randomized into five groups: a negative control, a positive control and three test groups, each containing eight rats. Treatment was administered by gavage. The administration volume was 10 mL/kg in all cases. Rats received vehicle (spring water) in the negative control group and loperamide hydrochloride (Sigma- Aldrich, Saint-Quentin Fallavier, France) at the dose of 5 mg/kg in the positive control group. In the three test groups, rats received the HLE of J. regia at the doses of 0.5, 1, or 2 g/kg. One hour after this treatment, diarrhea was induced by the administration of castor oil by gavage at the dose of 10 mL/kg (Sigma-Aldrich, Saint-Quentin Fallavier, France). Each animal was thereafter placed in an individual cage and observed for 4 hours. The following parameters were monitored: time to initial evacuation (onset time; min), total number of feces and diarrheal feces, fresh weight and water content of the feces (g), and body weight loss (g). Behavioral parameters i.e., eye closing, abnormal posture, fur aspect and activity were used as pain indices. Eye closing, abnormal posture and fur aspect were adapted from published studies [13,14]. Eye closing was scored 0 for complete opening (normal eyes), 1 for half-closed eyes and 2 for complete closing. Animal posture was scored 0 for normal posture, 1 for slightly arched back, 2 for arched back and 3 for very arched back. Fur aspect was scored 0 for normal fur, 1 for slightly ruffled fur, 2 for ruffled fur and 3 for very ruffled fur. Animal activity was scored 0 for intense activity (four-leg standing with no lying position; permanent movement and frequent rearing behavior), 1 for normal activity (four-leg standing with frequent movement and moderate rearing behavior), 2 for moderate activity (mainly on four legs with sporadic movement and low rearing behavior), 3 for little activity (partly four-leg standing and lying position with rare movement and no rearing behavior) and 4 for very little activity (mainly in lying position with very rare movement and no rearing behavior). The global behavioral score was calculated by adding the scores obtained for each behavioral parameter. The investigator was blind to the experimental status of the rats.
Statistical Analysis
Results are presented as mean ± standard error of the mean (SEM). The significance of differences between groups was evaluated using the Kruskal-Wallis one-way analysis of variance followed when significant, by the Mann-Whitney U-test for inter-group comparisons. For all statistical analyses, the level of significance was set at p<0.05. All statistical analyses were carried out using the software StatView®5 statistical package (SAS, Institute Inc., USA).
The UHPLC analysis of the HLE of J. regia revealed the presence of nine flavonoids mainly, quercetin, myricetin, kaempferol and taxifolin derivatives. Four hydroxycinnamic acids were also identified: chlorogenic, neochlorogenic, 3- and 4-p-coumaroylquinic acids (Figure 1 and Table 1). Content in flavonoids expressed as hyperoside equivalent was 2.94 mg/mL.
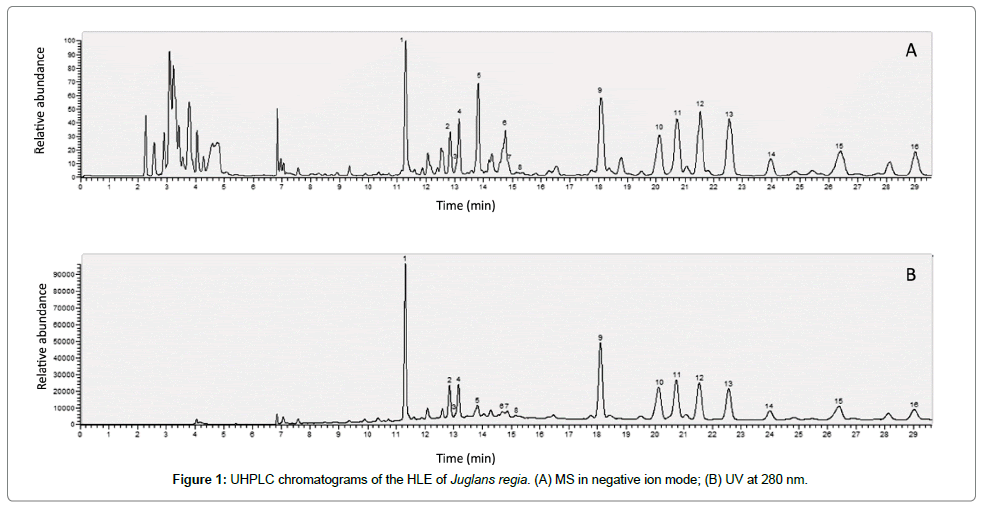
Figure 1: UHPLC chromatograms of the HLE of Juglans regia. (A) MS in negative ion mode; (B) UV at 280 nm.
| Peak | Retention time (min) |
Molecular ion [M-H]- (m/z) |
Formula | MS² (m/z) | Compounds | Reference |
|---|---|---|---|---|---|---|
| 1 | 11.29 | 353.0878 | C16H18O9 | 191/179/135 | Neochlorogenic acid | Standard |
| 2 | 12.86 | 337.0932 | C16H18O8 | 163/119/191 | 3-p-coumaroylquinic acid | [8] |
| 3 | 13.09 | 289.0721 | C15H14O6 | 109/125/203/151/123/245/205/221/137/149/161/97/179 | Catechin | Standard |
| 4 | 13.14 | 353.0877 | C16H18O9 | 191/161/89/59/115/ | Chlorogenic acid | Standard |
| 5 | 13.83 | 339.1084 | C23H16O3 | 159/161 | Unknown | [8] |
| 6 | 14.74 | 175.0393 | C10H8O3 | 175/174/176/147/131 | Unknown | |
| 7 | 14.85 | 337.0928 | C16H18O8 | 173/163 | 4-p-coumaroylquinic acid | [7] |
| 8 | 15.30 | 479.0838 | C21H20O13 | 316/479/271/165 | Myricetin-3-O-glucoside | [8] |
| 9 | 18.13 | 463.0883 | C21H20O12 | 271/300/255/243/151/179 | Hyperoside | Standard |
| 10 | 20.12 | 435.0932 | C20H20O11 | 151/285/435 | Taxifolin-pentoside derivative | [8] |
| 11 | 20.74 | 433.0771 | C20H18O11 | 300/271/433/255 | Quercetin-pentoside derivative | [8] |
| 12 | 21.53 | 433.0778 | C20H18O11 | 300/271/433/255 | Quercetin-pentoside derivative | [8] |
| 13 | 22.56 | 447.0933 | C21H20O11 | 271/300/255/151/243/179/163 | Quercitrin | Standard |
| 14 | 23.96 | 417.0828 | C20H18O10 | 284/417/255/227/285 | Kaempferol-pentoside derivative | [9] |
| 15 | 26.43 | 417.0829 | C20H18O10 | 284/285/417/255/227/256/327 | Kaempferol-pyranarabinoside | [9] |
| 16 | 29.01 | 501.1402 | C25H26O11 | 179/281 | Unknown |
Table 1: Identification of phenolic compounds in the HLE of Juglans regia in negative ions with UHPLC-MS and MS2.
The HLE of J. regia protected the rats against diarrhea induced by castor oil (Table 2). The onset time of diarrhea was three times higher in the HLE 2 g/kg group (186.1 ± 20.1 min) than in the vehicle group (61.3 ± 4.6 min, p<0.05). The increase in the onset time of diarrhea also observed in the HLE 1 g/kg group was not statistically significant compared with the vehicle. In addition, the total number of feces and diarrheal feces, fresh weight and water content of the feces, and body weight loss decreased significantly in both the HLE 1 g/kg and the HLE 2 g/kg groups compared with the vehicle group (Table 2). It is worth noting that a highly significant anti‑nociceptive activity was observed with HLE at the doses of 1 and 2 g/kg: the global behavioral scores were respectively reduced by 49.4% and 67.0% compared with the vehicle (p<0.001; Table 3). This was the result of a significant decrease in the scores of all behavioral parameters (activity, closing of the eyes, fur aspect, and posture).
| Doses* | Vehicle | Loperamide | Juglans regia HLE | ||
|---|---|---|---|---|---|
| 10 | 5 | 0.5 | 1 | 2 | |
| Onset time (min) | 61.3 ± 4.6 | 236.8 ± 3.3a | 87.0 ± 13.1 | 123.5 ± 23.4 | 186.1 ± 20.1a |
| Incidence of diarrhoea | 8/8 | 0/8 | 8/8 | 5/8 | 1/8 |
| Total number of feces | 14.3 ± 1.0 | 0.1 ± 0.1a | 12.4 ± 1.1 | 6.4 ± 1.8a | 3.5 ± 1.3a |
| Inhibition of defecation (%) | - | 99.1 | 13.2 | 55.3 | 75.4 |
| Total number of diarrhoeal feces | 9.3 ± 1.1 | 0.0 ± 0.0a | 6.0 ± 0.8b | 2.5 ± 0.9a | 0.5 ± 0.5a |
| Inhibition of diarrhoea (%) | - | 100.0 | 35.1 | 73.0 | 94.6 |
| Fresh weight of feces (g) | 5.61 ± 0.61 | 0.02 ± 0.02a | 4.26 ± 0.72 | 1.97 ± 0.61a | 0.91 ± 0.46a |
| Water content of feces (g) | 4.16 ± 0.42 | 0.01 ± 0.01a | 3.19 ± 0.59 | 1.31 ± 0.45a | 0.43 ± 0.23a |
| Body weight loss (g) | 12.1 ± 1.2 | 5.1 ± 0.4 a | 9.5 ± 0.9 | 7.9 ± 0.5 b | 5.5 ± 0.6 b |
Values are expressed as means ± SEM (n=8); * vehicle: mL/kg body weight, loperamide and J. regia HLE: mg/kg body weight ap<0.005, bp<0.05 when compared with the vehicle group.
Table 2: Anti-diarrheal activity of the HLE of Juglans regia in Wistar rats with castor oil-induced diarrhea.
The standard drug, loperamide, significantly reduced diarrhea and body weight loss (Table 2) but failed to reduce pain: the global behavioral score was not significantly different from that observed with the vehicle (p>0.05; Table 3). Compared to the loperamide group, the global behavioral score observed with the HLE was significantly reduced by 68.2% at the dose of 2 g/kg (p<0.001), 51.1% at 1 g/kg (p<0.005) and 18.9% at 0.5 g/kg (p<0.05).
| Doses* | Vehicle | Loperamide | Juglans regia HLE | ||
|---|---|---|---|---|---|
| 10 | 5 | 0.5 | 1 | 2 | |
| Activity (0-54) | 3.8 ± 0.2 | 3.8 ± 0.2 | 3.4 ± 0.2 | 2.3 ± 0.3 a | 1.1 ± 0.4 b |
| Posture (0-43) | 2.0 ± 0.2 | 1.9 ± 0.1 | 1.8 ± 0.2 | 1.1 ± 0.1 a | 0.9 ± 0.1 a |
| Fur (0-43) | 1.8 ± 0.2 | 1.6 ± 0.3 | 1.9 ± 0.1 | 0.9 ± 0.1 a | 0.8 ± 0.2 a |
| Eye (0-32) | 1.0 ± 0.3 | 1.5 ± 0.2 | 0.4 ± 0.2 | 0.0 ± 0.0 a | 0.0 ±0.0 a |
| Global behavioral score (0-12) | 8.5 ± 0.5 | 8.8 ± 0.6 | 7.4 ± 0.4 | 4.3 ± 0.4 b | 2.8 ± 0.5 b |
Values are expressed as means ± SEM (n=8); *Vehicle: mL/kg body weight, loperamide and J. regia HLE: mg/kg body weight; ap<0.005, bp<0.001 when compared with the vehicle and loperamide groups.
Table 3: Effect of the HLE of Juglans regia on behavioral parameters in Wistar rats with castor oil-induced diarrhea.
This study provides evidence that supports the traditional use of J. regia in diarrheal conditions. We demonstrated that our HLE of J. regia produced a dose-dependent and significant delay in the onset of diarrhea compared to the vehicle. The highest doses (1 and 2 g/kg) also significantly reduced the number of diarrheal feces, fresh weight and water content of the feces and body weight loss. Interestingly, unlike loperamide, the HLE of J. regia also showed anti-nociceptive activity. By being effective in resolving both acute diarrhea and associated pain, the HLE of J. regia could be an interesting alternative to standard antimotility drugs.
The anti-diarrheic and anti-nociceptive activities of our HLE may be due to a synergistic effect between hydroxicinnamic acids and flavonoids, which were detected in our extract. Hydroxycinnamic acids such as chlorogenic, neochlorogenic, 3- and 4-p-coumaroylquinic acids were previously identified in other types of J. regia leaf extracts [9,15,16]. Neochlorogenic and chlorogenic acids exhibit antioxidant and anti-inflammatory activities [17-19], which might protect the intestinal mucosa against castor oil. The effect of castor oil is mediated by ricinoleic acid, a hydroxylated fatty acid released from the substance by intestinal lipases. Ricinoleic acid causes local irritation and inflammation of the mucosa leading to the release of inflammatory mediators and an increase in the net secretion of water and electrolytes [20]. Chlorogenic acid was also found to alleviate pain in several animal models [21] and to attenuate diarrhea in a model of colitis in mice [22].
The anti-diarrheal activity of flavonoids was attributed to their ability to inhibit hydroelectrolytic secretions and intestinal motility [3]. In vitro and in vivo experiments showed that flavonoids can decrease the intestinal secretory response induced by prostaglandin E2 and the contractions induced by spasmogens [23]. Quercetin, liberated after the hydrolysis of quercetin-derived glycosides in the digestive tract, has anti-spasmodic [24] and anti-nociceptive activities [25]. It affects smooth muscle fibers as a calcium-antagonist agent [26] and exerts its anti-nociceptive effect by inhibiting pro-nociceptive cytokine production and the oxidative imbalance mediation of inflammatory pain [25].
Further studies will be needed to elucidate the exact mechanisms underlying the anti-diarrheal effects of our J. regia HLE and its phytochemical compounds with regard to anti-motility, anti-secretory and anti-inflammatory properties. Also, it would be very interesting to evaluate the possible mechanisms involved in its anti-nociceptive action.