Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2016) Volume 5, Issue 2
The protective role of lichens are getting more important. In this study, the antigenotoxic and antimutagenic activity of the methanol extracts of four lichen species (Anaptychia ciliaris, Bryoria fuscescens, Parmotrema chinensa and Xanthoria candelaria) were investigated. Sister Chromatid Exchange (SCE) test were used for determining the genotoxic contamination and the results showed that four lichen extracts inhibited the mutagenic effects of AFB1. Three different concentrations used and it was determined that the most effective concentration is 20 μg/mL againts the mutagenic effects of AFB1. In addition, the antioxidant effects of A. ciliaris, B. fuscescens, P. chinensa and X. candelaria were determined by measuring the activities of superoxide dismutase (SOD), glutathione peroxidase (GPx) and glutathione (GSH) and level of malondialdehyde (MDA) against the oxidative stress of AFB1 in human lymphocytes in vitro. It was observed that the levels of antioxidant enzymes were decreased dramatically and level of malondialdehyde (MDA) increased after treatment with AFB1 but the antioxidant enzymes were increased and MDA levels decreased in case of applying the lichen methanol extracts. We also determined that our lichen samples have strong antioxidative and antigenotoxic character and their antigenotoxic quality may be relation to the mechanisms of strong antioxidative property.
Keywords: Antioxidant; Antigenotoxicity; Lichens; Genotoxicity; AFB1; in vitro
Lichens are a form of symbiont between a fungus (mycobiont) and an alga and/or a cyanobacterium (photobiont), which contains a wide variety of organic compounds with certain secondary metabolite classes typical of these organisms [1-3]. Although classically described as symbiotic associations between photosynthetic microorganisms and fungi, lichens are actually extremely complex microbial communities [4]. They are known to be an excellent source of low molecular weight secondary compounds. More than 1000 secondary compounds have already been identified from various species of lichens and their cultured symbionts and their natural products have been used as cosmetics, decorations, dyes, foods, and medicines [5,6]. Lichens are known to have therapeutic effects on various diseases in traditional system of medicine of many countries. A number of factors such as specific and extreme habitat, slow growth and long life are the basis for the production of diverse bioactive compounds having protective functions against several physical and biological influences [6-8]. They have been attracting the attention of many researchers because of their diverse pharmaceutical potentials as shown in their antioxidant [9,10], antiviral [11], anti-proliferative [12,13], anti-inflammatory, anti-tumor [14-16], and insecticidal [17] activities. Also, lichens are considered to be a source of natural antioxidants [18,19]. Various lichen compounds, either as crude extracts or in purified form, have been screened against various cancer cell lines from solid tumors [20]. For ages, lichens have long been investigated popularly for biological roles, mainly antitumor, antimicrobial and antioxidant activities [21].
Reactive oxygen species (ROS) are important class of damage agents for cellular macromolecules. ROS, such as O2 −, OH− and H2O2, are highly genotoxic/ mutagenic and harmful to cellular macromolecules such as DNA, proteins and lipids and may be an important etiologic factor in degenerative diseases such as cancer, cardiovascular and cerebrovascular pathologies, as well as in the aging process [22,23]. However; H2O2 not only is a source of oxidative stress but also acts as an essential second messenger in signal transduction networks of normal, healthy cells, wherein growth factors, cytokines and a variety of other ligands trigger its production through the activation of their corresponding receptors [24]. Antioxidants are compounds which can impede the oxidation process by reacting with free radicals, chelating catalytic metals and scavenging oxygen in biological systems. So, antioxidants are of prime importance in preventing various pathophysiological dysfunctions and diseases [25-27]. Lichens and their metabolites have manifold biological activity, such as: antiviral [11], antioxidant [8,25-29], antimicrobial, anticancer [27,29,30]; antimutagenic [31]; antigenotoxic [29,30,32] and plant growth inhibitory, antiherbivore, ecological roles and enzyme inhibitory [29]. Hence, they have also, for hundreds of years, been used in many contry as a cure for diseases of humans. For example, Lobaria pulmonaria and Parmelia sulcata have been used in the treatment of pulmonary and cranial diseases, respectively. Similarly, Xanthoria parietina was used to cure jaundice and Letharia vulpina in stomach diseases. The usage of some lichens for many years in the traditional medicine was later justified by numerous researches that confirmed their various biological activities [29,33-35].
Antioxidant activity of various lichens are known, such as: Cladonia furcata, Lecanora atra, Lecanora muralis [29], Parmelia sulcata, Flavopamelia caperata, Evernia prunastri, Hypogymnia physodes, Cladonia foliacea [8], Evernia prunastri, Pseudoevernia furfuraceae [4]. So far, only few research demonstrated that lichen extracts have antigenotoxic role, such as: Rhizoplaca chrysoleuca, Lecanora muralis [32], Cladonia foliacea [28], Lasallia pustulata [30]. Moreover, no previous work has been encountered regarding the antigenotoxic and antioxidant activities of the lichens A. ciliaris, B. fuscescens, P. chinensa and X. Candelaria in the literature (Figures 1-4).
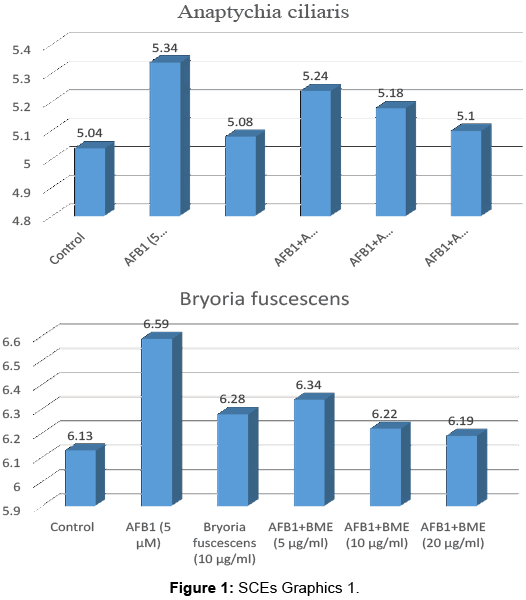
Figure 1: SCEs Graphics 1.
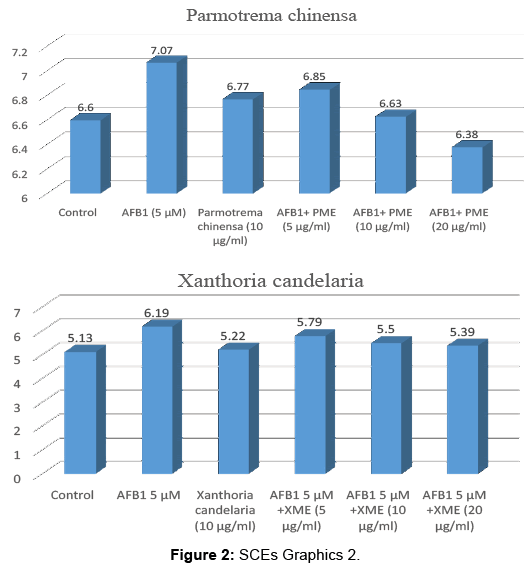
Figure 2: SCEs Graphics 2.
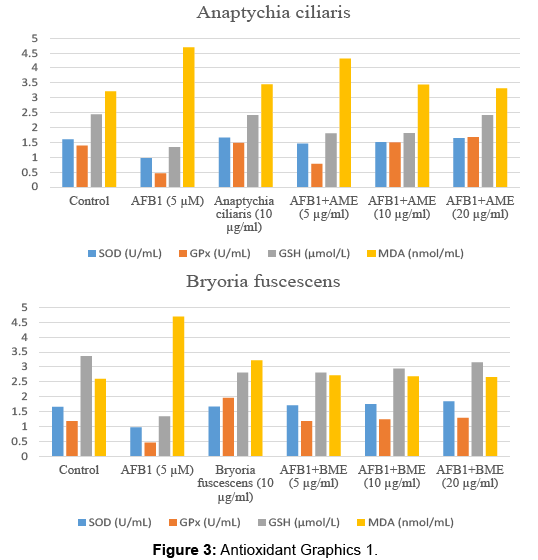
Figure 3: Antioxidant Graphics 1.
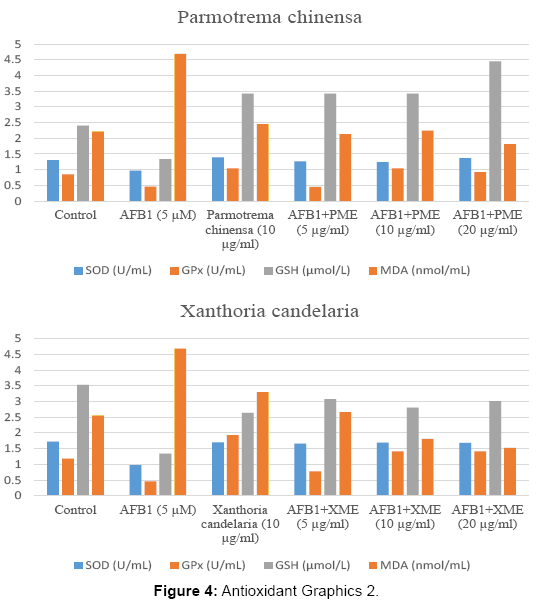
Figure 4: Antioxidant Graphics 2.
Within the frame of the aforementioned reasons, we previously aimed to examine the in vitro antioxidant and antigenotoxic activities of the methanol extracts of the lichens A. ciliaris, B. fuscescens, P. chinensa and X. Candelaria. Antigenotoxic activity was determined by SCE assay and aflatoxin B1 (AFB1), was used as a positive control. The antioxidant potency of the lichens was investigated by measuring the SOD, GPx activities and the levels of GSH, MDA.
Samples and preparation of methanol exracts
Lichen samples were collected from Artvin and Erzurum provinces between 10-20 August 2011, Turkey. This samples were identified using different lichen flora books and papers [28-30] by Ali Aslan. Samples are stored in the herbarium of Ali Aslan laboratory with sample labels (ATA-KKEF 1839; ATA-KKEF 1840; ATA-KKEF 1841; ATA-KKEF 1842), Ataturk University, Erzurum, Turkey.
Lichen samples were made into powder. 10 g powder were extracted with 250 mL of methanol using the Soxhlet extractor (Isopad, Heidelberg, Germany) for 24 hours at room temperature [31]. After extraction, methanol extract was filtered with paper (Whatman filter) and evaporation. The extract was lyophilized and kept in the dark at +4°C.
Microscopic evaluation
Human lymphocytes were collected from healthy voluntary donors (ages range 24-28). Lymphocyte cultures (obtained from peripheral blood) cultivated into RPMI-1640 chromosome medium. Different concentration of sample doses of four lichen species were added this medium with AFB1 (5 μM concentration).
For SCE experiment, heparin at a ratio of 1/10 was added to the 1 ml peripheral blood samples of donors. The blood samples were added to the 5 ml chromosome medium B supplemented with 6 μg/ ml 5’-bromo-2’-deoxyuridine in sterile conditions. The cultures were incubated at 37°C for 72 hours. 0.06 μg/ml colchicine was added at 2 h before the harvesting of the culture. The SCE tests were performed as described by Perry and Evans [36], but with some modifications [37]. Next, slides were stained with 5% Giemsa (pH = 6.8) prepared in Sorensen buffer solution, for 20–25 min; washed in distilled water; dried at room temperature [32]. At the end of the experiment, 20 suitable methaphases recorded and well-spread second division metaphases containing 42–46 chromosomes in each cell were scored, and the values obtained were calculated as SCEs per cell.
Biochemical analysis for SOD, GPx, GSH and MDA assays
The peripheral blood supernatant were collected for immediately assayed for enzyme activities. All samples were centrifuged at 3000 rpm for 10 min at 4°C. After that Cu, Zn-SOD and GPx activity and GSH and MDA levels were measured by the method of (with some modifications) Paglia and Valentina [35], Sun [36] and Ohkawa [35], respectively. All samples were measured in sixfold.
SOD assay: For SOD assay, supernatant was treated with 2.45 mL of assay reagent 100 μL of the sample. Xanthine oxidase was determined by measuring the absorbance at 560 nm. Cu, Zn-SOD activity in the cell culture supernatant was detected by the method of Ref. [34].
GPx assay: For GPx assay, supernatant was treated with 100 μL of 8 mM NADPH, 100 μL of 150 mM reduced GSH, 20 μL of glutathione reductase (30 units/mL), 20 μL of 0.12 M sodium azide solution and 2.65 mL of 50 mM potassium phosphate buffer (pH 7.0, 5 mM EDTA). Atfer the treatment, the solition was mixed with NADPH to NADP and measured measured spectrophotometrically for 5 min at 340 nm. GPx activity in the cell culture supernatant was measured by the method of Paglia and Valentine [33].
GSH assay: For GPx assay, supernatant was treated with 3 mL cuvette and then 750 μL of 10mM 5-5’-dithio-bis-2-nitrobenzoic acid (DTNB) solution. 150 μL of 1.47mM β-NADPH was added the cuvette and was measured spectrophotometrically for 2 min at 412 nm. GSH levels in the cell culture supernatant were assessed according to the method of Tietze and Anderson [36,37].
MDA assay: For GPx assay, supernatant was treated with 8.1% sodium dodecyl sulphate, 20% acetic acid, 0.9% thiobarbituric acid. After, samples were centrifuged at 4000 X g for 10 min and this solution was measured at 532 nm. MDA levels in the cell culture supernatant were determined spectrophotometrically according to the method described by Ref. [35], and protein concentrations were determined by Bradford method [38]. All photometrical measurements were performed with a DU 530 spectrophotometer (Beckman Instruments, Fullerton, California, USA) in a quartz cuvette.
Statistical analysis
SCE values and biochemical parameters were analysed Oneway ANOVA and Mann-Whitney U test. For SCE statistical analysis, a value of p less than 0.05 was acceped as statistically significant. For biochemical parameters statistical analysis, a value of p less than 0,001 was acceped as statistically significant. Results were expressed as mean ± SD. All data were evaluated with SPSS 16.0 version for Windows (SPSS Inc, Chicago, Illinois, USA).
AFB1 significantly increased the SCE frequence on peripheral lymphocytes when compared with the control as seen in Tables 1-4. Such an increase was found to be statistically significant (p<0.05). It was obverved that treatment group with different concentrations of AME, BME, PME and XME together with AFB1 decreased the SCE frequence compared with intoxicated with AFB1. All concentrations are reduced SCE frequencies especially 20 μg/mL is the most effective dose.
| Metaphase | SCE Frequence | SCE counted |
SCE/cell ±S.S | |
|---|---|---|---|---|
| Control | 60 | 2-8 | 303 | 5.04 ± 0.88a |
| AFB1 5 µM | 60 | 5-11 | 321 | 5.34 ± 6.77d |
| Anaptychia ciliaris (10 µg/ml) | 60 | 3-9 | 305 | 5.08 ± 1.20ab |
| AFB1 5 µM+AME(5 µg/ml) | 60 | 4-9 | 315 | 5.24 ± 4.15cd |
| AFB1 5 µM+AME (10 µg/ml) | 60 | 4-10 | 311 | 5.18 ± 3.84bc |
| AFB1 5 µM+AME(20 µg/ml) | 60 | 4-9 | 306 | 5.10 ± 3.52ab |
Table 1: Anaptychia ciliaris.
| Metaphase | SCE Frequence |
SCE counted | SCE/cell ±S.S | |
|---|---|---|---|---|
| Control | 60 | 4-7 | 368 | 6.13 ± 2.88a |
| AFB1 5 µM | 60 | 6-13 | 396 | 6.59 ± 7.77c |
| Bryoria fuscescens (10 µg/ml) | 60 | 4-9 | 377 | 6.28 ± 4.97ab |
| AFB1 5 µM +BME(5 µg/ml) | 60 | 3-10 | 381 | 6.34 ± 4.09bc |
| AFB1 5 µM +BME(10 µg/ml) | 60 | 5-10 | 373 | 6.22 ± 6.69ab |
| AFB1 5 µM +BME(20 µg/ml) | 60 | 4-11 | 371 | 6.19 ± 2.18ab |
Table 2: Bryoria fuscescens.
| Metaphase | SCE Frequence |
SCE counted | SCE/cell ±S.S | |
|---|---|---|---|---|
| Control | 60 | 4-9 | 396 | 6.60 ± 2.11a |
| AFB1 5 µM (84ml) | 60 | 6-10 | 424 | 7.07 ± 3.37ef |
| Parmotrema chinensa (10 µg/ml) | 60 | 2-8 | 406 | 6.77 ± 2.17ab |
| AFB1 5 µM +PME (5 µg/ml) | 60 | 5-10 | 411 | 6.85 ± 3.01de |
| AFB1 5 µM +PME (10 µg/ml) | 60 | 5-11 | 398 | 6.63 ± 3.16d |
| AFB15 µM +PME (20 µg/ml) | 60 | 4-9 | 383 | 6.38 ± 3.07c |
Table 3: Parmotrema chinensa.
| Metaphase | SCE Frequence |
SCE counted | SCE/cell ±S.S | |
|---|---|---|---|---|
| Control | 60 | 2-9 | 308 | 5.13 ± 1.82a |
| AFB1 5 µM | 60 | 5-10 | 371 | 6.19 ± 2.17d |
| Xanthoria candelaria (10 µg/ml) | 60 | 4-8 | 313 | 5.22 ± 1.97ab |
| AFB1 5 µM+XME(5 µg/ml) | 60 | 5-10 | 347 | 5.79 ± 2.01cd |
| AFB1 5 µM+XME(10 µg/ml) | 60 | 4-11 | 330 | 5.50 ± 1.69bc |
| AFB15 µM+XME(20 µg/ml) | 60 | 6-11 | 323 | 5.39 ± 1.42b |
Table 4: Xanthoria candelaria.
Tables 5-8 represent us the control and experimental groups of enzyme activities such as SOD, GPx and GSH, MDA levels. AFB1 significantly decreased the activities of SOD, GPx and GSH level and MDA level increased (p<0.001). Nonetheless after treatment of different concerations of AME, BME, PME and XME, the activities of SOD, GPx and GSH level increased and MDA level decreased.
| SOD (U/mL) | GPx (U/mL) | GSH (µmol/L) | MDA (nmol/mL) | |
|---|---|---|---|---|
| Control | 1.60 ± 0.05 | 1.39 ± 0.02 | 2.44 ± 1.14 | 3.21 ± 0.57 |
| AFB1 5 µM | 0.97 ± 0.11a | 0.46 ± 0.16a | 1.34 ± 0.11a | 4.69 ± 2.03a |
| Anaptychia ciliaris (10 µg/ml) | 1.66 ± 0.03b | 1.48 ± 0.03b | 2.41 ± 0.18b | 3.45 ± 0.02b |
| AFB1 5 µM+AME(5 µg/ml) | 1.46 ± 0.04b | 0.78 ± 0.31abc | 1.80 ± 0.77abc | 4.31 ± 0.15b |
| AFB1 5 µM+AME (10 µg/ml) | 1.51 ± 0.02b | 1.50 ± 0.11abd | 1.81 ± 0.11abc | 3.44 ± 0.01bcd |
| AFB1 5 µM+AME(20 µg/ml) | 1.64 ± 0.01b | 1.68 ± 0.11abd | 2.42 ± 0.51bde | 3.31 ± 0.11abcd |
Table 5: Anaptychia ciliaris.
| SOD (U/mL) | GPx (U/mL) | GSH (µmol/L) | MDA (nmol/mL) | |
|---|---|---|---|---|
| Control | 1.66 ± 0.01 | 1.18 ± 0.01 | 3.36 ± 1.57 | 2.60 ± 0.22 |
| AFB1 5 µM | 0.97 ± 0.11a | 0.46 ± 0.16a | 1.34 ± 0.11a | 4.69 ± 1.03a |
| Bryoria fuscescens (10 µg/ml) | 1.67 ± 0.01b | 1.96 ± 0.13ab | 2.81 ± 1.57ab | 3.22 ± 2.40a |
| AFB1 5 µM+BME(5 µg/ml) | 1.71 ± 0.01bc | 1.18 ± 0.10bc | 2.81 ± 0.25ab | 2.72 ± 0.50a |
| AFB1 5 µM+BME(10 µg/ml) | 1.75 ± 0.01b | 1.24 ± 0.12abc | 2.94 ± 1.41ab | 2.68 ± 0.50ab |
| AFB1 5 µM+BME(20 µg/ml) | 1.84 ± 0.02abc | 1.29 ± 0.01abcde | 3.15 ± 0.38abcd | 2.66 ± 1.44abc |
Table 6: Bryoria fuscescens.
| SOD (U/mL) | GPx (U/mL) | GSH (µmol/L) | MDA (nmol/mL) | |
|---|---|---|---|---|
| Control | 1.30 ± 0.02 | 0.85 ± 0.24 | 2.40 ± 0.61 | 2.22 ± 1.57 |
| AFB1 5 µM (84ml) | 0.97 ± 0.11a | 0.46 ± 0.16a | 1.34 ± 0.11a | 4.69 ± 2.03a |
| Parmotrema chinensa (10 µg/ml) | 1.39 ± 0.01ab | 1.04 ± 0.37ab | 3.42 ± 0.59ab | 2.45 ± 1.05ab |
| AFB1 5 µM +PME (5 µg/ml) | 1.26 ± 0.02bc | 0.45 ± 0.34abc | 3.42 ± 0.41ab | 2.13 ± 0.18b |
| AFB15 µM +PME (10 µg/ml) | 1.24 ± 0.02abc | 1.04 ± 0.84abd | 3.42 ± 0.59ab | 2.24 ± 0.01b |
| AFB15 µM +PME (20 µg/ml) | 1.37 ± 0.02abde | 0.92 ± 0.05bd | 4.45 ± 0.59abcde | 1.81 ± 0.18abc |
Table 7: Parmotrema chinensa.
| SOD (U/mL) | GPx (U/mL) | GSH (µmol/L) | MDA (nmol/mL) | |
|---|---|---|---|---|
| Control | 1.72 ± 0.02 | 1.18 ± 0.12 | 3.53 ± 0.50 | 2.56 ± 0.32 |
| AFB1 5 µM | 0.97 ± 0.11a | 0.46 ± 0.16a | 1.34 ± 0.11a | 4.69 ± 2.03a |
| Xanthoria candelaria (10µg/ml) | 1.70 ± 0.02b | 1.93 ± 0.43ab | 2.64 ± 0.41b | 3.30 ± 1.51ab |
| AFB1 5 µM+XME(5 µg/ml) | 1.66 ± 0.01b | 0.77 ± 0.10abc | 3.08 ± 0.17abc | 2.66 ± 1.02bc |
| AFB1 5 µM+XME(10 µg/ml) | 1.69 ± 0.02b | 1.41 ± 0.16abcd | 2.81 ± 0.01abc | 1.81 ± 0.18abc |
| AFB1 5 µM+XME(20 µg/ml) | 1.68 ± 0.04b | 1.41 ± 0.16abc | 3.01 ± 0.01abc | 1.52 ± 1.46abcde |
Table 8: Xanthoria candelaria.
According to the literature, many of the lichen species have important biological properties including antioxidant, anticancer, antigenotoxic and antiproliferative [10,29,31]. Current report collects data related to antioxidant activities of more than 75 lichen species (from 18 botanical families) and 65 isolated metabolites. Much information comes from in vitro investigations, such as chemical assays evaluating radical scavenging properties, lipid peroxidation inhibition, and reducing power of lichen species and compounds; similarly, research on cellular substrates and animal models generally measures antioxidant enzymes levels and other antioxidant markers, such as glutathione levels or tissue peroxidation [38,39]. For example, Gulcin et al. found that the aqueous extracts of Cetraria islandica had a strong antioxidant activity [39]. Similar results were reported by Behera et al. for different extracts from the lichen Usnea ghattensis [40]. Kekuda et al. find an antioxidant activity for the extracts of the lichen Parmotrema pseudotinctorum and Ramalina hossei and Manojlovic et al. explored antioxidant properties of Laurera benguelensis [41]. In this study we determined that methanol extracts of the lichens A. ciliaris, B. fuscescens, P. chinensa and X. Candelaria had a strong antioxidant capacity by measuring the activities of superoxide dismutase (SOD), glutathione peroxidase (GPx) and glutathione (GSH) and level of malondialdehyde (MDA) against the oxidative stress of AFB1 in human lymphocytes in vitro. It was observed that the levels of antioxidant enzymes were decreased dramatically and level of malondialdehyde (MDA) increased after treatment with AFB1 (Tables 5-8). Based on these results, it could be concluded that antioxidative nature of the extracts might depend on their phenolics. Phenolic components are potential antioxidants, free radical terminators [25]. Many researches found high correlations between antioxidative activities and phenolic content such as epigallocatecthin gallate, quercetin, gallic acid, curcumin, eugenol, usnic acid, polysaccharide Ci-3, lichestrerinic acid, protolichesterinic acid and organic acids such as oxalic, fumaric, malic and lactic acids [42-45]. An oposite finding of Odabasoglu et al. with methanol extracts of Lobaria pulmonaria and Usnea longissima, were explained by the participation of other, non-polar components, insoluble in methanol in this activity [46].
In order to elucidate the antigenotoxic activities of the lichens A. ciliaris, B. fuscescens, P. chinensa and X. Candelaria, SCE system was performed in this study and aflatoxin B1 (AFB1), was used as a positive control. Aflatoxin B1 represent food-contaminating mutagens, as found in some nuts/cereals and cooked meat [47]. This agent is known to stimulate the release of free radical, including reactive oxygen species, which leads to chromosomal aberrations [48]. Its mutagenic effects have been well documented in a number of in vitro and in vivo models, where the presence of DNA adducts, DNA breaks, gene mutations, induction of DNA synthesis and inhibition of DNA repair have been determined, as well as increases in the rate of chromosomal aberrations, micronuclei and sister chromatid exchanges (SCE) [48,49]. According to our results, the extracts significantly exhibited antigenotoxic potential against AFB1 at with applications of increasing doses. The antigenotoxic activity may be explained with inhibitor activities of the lichen extracts on the formation of free radicals. The results of this study also showed that AFB1 increased SCE frequencies and decreased the activities of enzymes SOD, GPx, and GSH level, but increased MDA level (Tables 1-8). Our results indicate that the antigenotoxic effects of the four lichens species could be related to its antioxidant potential.
As a result, it can be stated that tested lichen extracts have a strong antioxidant and antigenotoxic activity in vitro. On the basis of these results, the lichens which are tested in this study could be good and safe natural antioxidant and anticancer agents and also, could be of indispensible in human therapy, animal and plant diseases. Further studies should be concentrated to search new compounds from lichens that exhibit strong antioxidant, anticancer and antigenotoxic activity for anticancer drug development.