Journal of Antivirals & Antiretrovirals
Open Access
ISSN: 1948-5964
ISSN: 1948-5964
Research Article - (2013) Volume 5, Issue 3
Objectives: The present investigation was carried out to test anti-HIV-1 activity of pure compounds isolated from aerial part extracts of Ocimum basilicum and its parasite Cuscuta campestris.
Materials: The anti-HIV-1 activity of these extracts was performed by use of real-time polymerase chain reaction (PCR) assay and high pure viral nucleic acid kit.
Results: The most active fractions were detected by NMR as eugenol and eugenol epoxide respectively in Ocimum basilicum and Cuscuta campestris. The apparent effective concentrations for 50% plaque reduction (EC50) of eugenol and eugenol epoxide were 350 and 80 μg/ml. Time of addition studies demonstrated that the inhibitory effect of eugenol and eugenol epoxide is higher respectively in pre HIV-1 infection and during infection.
Conclusion: These results demonstrate that eugenol and eugenol epoxide may prevent HIV-1 replication with two different mechanisms.
Keywords: HIV-1; Antiviral activity; Eugenol; Real time PCR assay
HIV-1 is an important pathogen and causes the majority of HIV infections globally [1]. Present anti HIV drugs have many disadvantages including resistance, toxicity, limited availability and lake of any curative effect [2]. Currently, many researches have been performed worldwide to isolate the active novel compounds from plants for preventing transmission of HIV and treatment of patients [3]. A diversity of secondary metabolites obtained from these products including alkaloids, polyphenols, flavonoids, sulphated polysaccharides, coumarines and triterpenes [4,5]. These compounds inhibit unique enzymes and proteins critical to the life cycle of HIV, including the reverse transcription process, virus entry, the integrase or protease [6,7]. Screening anti-HIV agents from natural products may be a more effective way for drug discovery. O. basilicum (family Lamiaceae) is an annual plant native to Asia [8]. The extract of O. basilicum has been investigated for several biological activities [9,10]. In recent decades, some bioactive compounds such as caffeic and p-coumaric acid have been isolated from O. basilicum [11]. O. basilicum have been recorded as a common host for Cuscuta campestris in Iran [8]. The parasitic relationship of C. campestris with its hosts has been subjected by numerous investigations [12,13]. Some specious of Ocimum and Cuscuta have been reported to have potent anti-viral activity [14,15]. Therefore, in the present study, anti-HIV-1 activity of Ocimum basilicum and its parasit Cuscuta campestris has been studied.
Plant material
The aerial parts of O. basilicum infected by C. campestris were collected from University of Isfahan herbarium, Iran. The aerial parts of these two plants were separated and dried.
Extraction and isolation of compounds
Methanol extract (98%) of dried parts of O. basilicum and C. campestris were prepared. The extraction was done thrice at 40°C. Then, the resulting liquid was collected, filtered and reduced through evaporation by a rotary evaporator (Stroglass, Italy) at 45°C and dried using a freeze dryer (Zirbus, Germany). Silica-gel column fractionation chromatography was carried out separately with the dried methanol extract of O. basilicum and C. campestris.
Dried methanol extract of O. basilicum (5 g) was eluted with Hexane: Acetone: Methanol (8:2:0 - 0:4:6, v/v/v) and 100% methanol. Fractions 1-13 (0.25, 0.25, 0.31, 0.45, 0.51, 0.36, 0.38, 0.36, 0.45, 0.31, 0.20, 0.31, 0.48 g) were obtained. Fraction 9 was found to have anti- HIV-1 activity and rechromatographed on silicagel column eluted with Ethyl acetate: Me OH (2:1, 1:1, 1:2, v/v) to yield fractions 9a, 9b, 9c (0.12, 0.1, 0.22). Fractions 9c were the most active fraction and were analyzed by NMR as eugenol.
Dried methanol extract of C. campestris (5 g) was eluted with Hexane: Aceton: Methanol: (8:2:0 - 0:4:6, v/v/v) and 100% methanol. Fractions 1-10 (0.45, 0.7, 0.40, 0.46, 0.35, 0.46, 0.48, 0.52, 0.40, 0.64 g) were obtained. Fraction 10 was found to have anti-HIV-1 activity. Fraction 10 was further purified by re-chromatography on silica gel column with Ethyl acetate: Me OH (3:1, 2:1, 1:1 v/v). Fraction 10 was separated to fractions 10a, 10b and 10c (0.2, 0, 0.19, and 0.2). Fractions 10a and 10c were the most active fractions and analyzed by NMR as eugenol and eugenol epoxide respectively.
HPLC analysis
HPLC screening of fraction 9c (isolated from Ocimum basilicum) and fraction 10a and 10c (isolated from Cuscuta campestris) were carried out. HPLC was performed on a HITACHI Series HPLC system equipped with L-7100 pump and an L-7100 UV-Vis detector. Peaks were separated on a reverse phase C18 column using the mobile phase [methanol/acetonitrile (70:30 v/v)]. The flow rate of the mobile phase was 1 .5 ml min-1. The absorption of analytes was detected at 450 nm. Samples were injected to the HPLC bed manually with injection volume as 5 ll. T2000 software was used for peak integration and calculation.
NMR analysis
NMR screening was used to approve structure of active compounds. 1H NMR spectra were recorded on Bruker 500 MHz spectrometer by use of CDCl3 as residual solvent with chemical shifts expressed in parts per million (ppm).
Cells and viruses
Peripheral blood mononuclear cells (PBMCs) were obtained from HIV-1 seronegative donors by lymphodex density centrifugation. The cells were grown in RPMI supplemented with 10% (v/v) Fetal Calf Serum (FCS; Gibco-BRL, Grand Island, NY, USA), 100 U/ml penicillin, 100 mg/ml streptomycin, 2 mM glutamine and 1 mM Na-pyruvate and activated with 5 μg/ml PHA. All of material was purchased from Gibco. A virus stock of HIV-1 subtype B was obtained from Azahra Hospital, Isfahan, Iran. Aliquots of 6×105 cells were infected with a low multiplicity of virus and incubated for 4 days. Then, supernatant of viral stock was harvested every day over a period of 4 days postinfection. Virus titers were determined by the HIV-1 p24 antigen kit (BioMerieux, France). The virus stocks with titers as high as 3×104 infectious units/ml have been obtained. The viruses were stored at -70°C until use.
PBMCs proliferation assay
The proliferation of PHA-stimulated PBMC was determined in the presence of pure compounds isolated from O. basilicum and C. campestris. The compounds were initially dissolved in dimethyl sulfoxide (DMSO) and then diluted with RPMI medium to prepare working concentrations of 2000, 1000, 500, 250, 125, 60 μg/ml. To avoid toxicity or interference by the solvent, the maximum concentration of DMSO in the test medium was 0.019%.
Cellular proliferation of the different concentrations of these compounds on cultured cells was measured using modified 3- (4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. The PBMCs were grown in 96-well plates at a density of 5×104 cells per well. After 72 h of incubation at 37°C, cell viability was examined according to Morgan [16] method with some modification. MTT solution (5 mg/ml) was added to each well, and the plate was incubated for 4 h. Finally, 50 μl of PrOH/HCl/TX (0.04 M HCl in 2-propanol plus 10% Triton X-100) was added to solubilize the formed formazan crystals. The plate was reincubated for 12 h and amount of formazan crystal was determined by measuring the absorbance at A540 nm using a microplate spectrophotometer (Awareness Technology Inc., stat fax 2100).
The PBMC proliferation index (PPI) was calculated as follow:
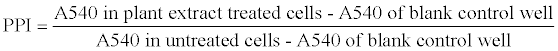
Antiviral activity
Anti-HIV-1 activity of active compounds was investigated via the HIV-1 p24 Antigen kit with 24 h old PBMCs grown in microtitre tissue culture plates. The cell were infected with 10 μl of HIV-1 supplemented with different concentrations of extract (500, 250, 100, 50 μg/ml) and incubated at 37°C for 12 hours. Then infected cells were washed and overlaid with medium at different concentrations of extract. 0.1% DMSO and different concentrations of zidovudine (AZT) (20, 10, 5, and 2.5 μg/ml) were also used as negative and positive controls. After 3 days of incubation, the overlay medium was used to detect and quantify HIV-1 p24 core protein. At the end, supernatant was transferred to the coated 96-well plate for the p24 assay. The protocol was followed as described by the manufacturer, with absorbance measured at 450 nm.
Time of addition study
The time-of-addition effect of eugenol and eugenol epoxide at EC50 concentrations was examined as described by Yang et al. [17]. PBMCs, 2×105 per well, were seeded into 24-well culture plates (Nunc; Nalge Nunc International, Rochester, NY, USA) and incubated for 24 h. Then, PBMCs were treated with 350 μg/ml of eugenol and 80 μg/ml of eugenol epoxide either concurrent with HIV-1 (0 h) or at intervals of 6 and 12 h pre-infection or also post-infection. After incubation at 37°C for 72 h, the reduction in the virus titer was measured by realtime polymerase chain reaction (PCR) assay and infection cultures containing the extracts were compared with that of the control cultures for each treatment.
Quantitative real-time PCR assay for HIV-1
For real-time PCR, 200 ml of supernatant of each treated-infected, or untreated-infected (virus control) wells were collected. RNA was purified from 200 μl of each specimen by High Pure viral nucleic acid kit (Roche Diagnostics, Meylan, France) according to the standard protocol. HIV-1 RNA was detected and quantified by real-time reverse transcription PCR (RT-PCR). As previously described, forward primer NEC152 (5′-CCTCAATAAAGCTTGCCTTGA-3′) and the reverse primer NEC131 (5′-GGCGCCACTGCTAGAGATTTT-3′) and the dually labeled NEC-LTR probe (5′-6-carboxyfluorescein- A G T A G T G T G T G C C C G T C T G T T R T K T G A C T - 6 - carboxytetramethylrhodamine-3′) in the long terminal repeat gene were used [18]. Primers and probes were synthesized by Metabion Co. (Germany). The master mix contained 1x RNA master hybridization buffer, including the Tth DNA polymerase and deoxynucleoside triphosphate mix (containing dUTP instead of dTTP), 2.5 Mn (OAC) 2, and 0.3 μM concentrations of each primer and probe. Cycling conditions were as follows: initial reverse transcription at 61°C for 30 min, denaturation at 95°C for 30 s, and 45 cycles of denaturation at 95°C for 1 s, annealing at 55°C for 15 s and elongation at 65°C for 1 min with a ramp of 5°C/s (with fluorescence acquisition at the end of each elongation stage). The positive control was consisted of culture supernatant of the HIV-1. As a control for cross-contamination, a sample of distilled water was subjected to the RNA extraction procedure, and the resulting extract was amplified in duplicate [19].
Statistical analysis
Data from three independent experiments are presented as mean ± SD. The CC50 and EC50 values were calculated by Microsoft Excel 2007. A selectivity index (SI) was calculated for each viral strain by the ratio of CC50 to EC50 value. The Student’s unpaired t-test was used to assess significance between the test sample and solvent control. P-value <0.05 was considered to be statistically significant.
HPLC analysis
Figures 1A and 1B shows the HPLC chromatograms of the fraction 9c of Ocimum basilicum and fraction 10c of Cuscuta campestris. After purification only one peak, 9c, with high intensity was observed at retention time 2.8. The HPLC chromatogram of fraction 10c was also observed at retention time 3.2 min. the HPLC chromatograms of fraction 10a of Cuscuta campestris was same as 9c of Ocimum basilicum (data not shown).

Figure 1: HPLC analysis of the eugenol (A), eugenol epoxide (B) isolated from Ocimum basilicum and Cuscuta campestris. Chromatographic conditions: C18 column using the mobile phase: methanol/ acetonitrile (70:30 v/v) at 1.5 ml min–1.
NMR analysis
The most active fraction obtained from O. basilicum was fraction 9c, which analyzed by NMR experiment as eugenol. Fractions 10a and 10c were also isolated from C. campestris and determined as eugenol and eugenol epoxide.
Eugenolepoxide
1HNMR (CDCl3, 400 MHz): δ 6.80 (d, 1H, J = 7.9 Hz), 6.75 (d, 1H, J =1.8 Hz), 6.64 (dd, 1H, J = 1.8, 7.9 Hz), 5.60 (s, 1H), 3.85 (s, 3H), 3.02 (m, 1H), 2.80 (dd, 1H, J1 = 5.5, J2 = 14.7 Hz), 2.73 (dd,1H, J1 =2.4, J2 = 4.9 Hz), 2.68 (dd,1H, J1 = 5.6, J2 = 14.8 Hz), 2.50 (dd,1H, J1 = 4.9, J2 = 2.4 Hz).
Eugenol
1HNMR (CDCl3, 400 MHz):δ 7.12 (s, 1H), 6.78 (d, 2H, J =7.2 z), 6.32-6.42 (m, 2H), 5.45-5.65 (m, 2H), 3.85 (s,3H), 3.26 (m, 2H)
13CNMR (CDCl3, 100 MHz) δ 147.23, 145.01, 138.23, 133.14, 122.15, 117.01, 114.52, 112.36, 56.24, 41.03.
PBMCs proliferation assay
Eugenol significantly induced PBMCs proliferation in a dose dependent manner upto 500 μg/ml. The result showed that 500 μg/ml of eugenol can increase the number of cells more than 3 folds and CC50 of this compound was obtained at 1500 μg/ml. Eugenol epoxide didn’t induce PBMCs proliferation and CC50 of this compound was attained at 1700 μg/ml (Figure 2).

Figure 2: Effect of eugenol and eugenol epoxide on PBMCs proliferation assay.
Anti-HIV-1 activity
Antiviral activity of eugenol and eugenol epoxide isolated from O. basilicum and C. campestris were evaluated by the HIV-1 p24 Antigen kit. Results showed that 500 μg/ml eugenol and eugenol epoxide inhibit viral replication >90%. The antiviral activity of these two compounds was further examined by different concentrations of 250, 100, 50 μg/ ml (Figure 3).

Figure 3: Effect of eugenol and eugenol epoxide isolated from O. basilicum, C. campestris and AZT on HIV-1 replication in Lymphocyte cells. The 50% inhibitory concentration (EC50) of each extract was calculated using regression line. Each bar represents the mean ± SD of three independent experiments.
The effective dose to reduce virus titers by 50% ( EC50) of eugenol and eugenol epoxide were observed as 350 and 80 μg/ml, respectively. Also the calculated selective index (SI) for eugenol and eugenol epoxide were 4.2 and 21.25 respectively.
Mechanism of action on HIV replication
A time-of-addition experiment was performed by measuring the viral RNA yields in infected culture supernatants, by means of a realtime PCR assay. Eugenol and eugenol epoxide were added before (-12 and -6 h), during (0 h) and after (6 and 12 h) virus absorption in the culture medium until the end of experiment. The curves generated during PCR reactions showed a prominent drop in HIV-1 DNA amounts in treated cultures compared to those of virus controls. The strongest HIV-1 proliferation in cells treated with eugenol and eugenol epoxide was achieved when the extracts were added respectively before and during the initial stages of infection. The results showed that the amount of viral DNA in eugenol treated cells at 12 and 6 h before infection was 104 to 105 folds lower compared to untreated control (Figure 4A). In contrast, the amount of DNA in cells treated with eugenol epoxide was 104 lower during infection compared to untreated control (Figure 4B).

Figure 4: Time of addition effect of eugenol (A) and eugenol epoxide (B) isolated from O. basilicum and C. campestris on HIV-1 replication. The extracts was added either before (-12 and -6), during (0) and after (6 and 12) virus infection. The extracts added before virus infection was rinsed off prior to the virus exposure. Each value is the result of mean ± SD of three independent experiments.
These studies suggest that there is a relationship between the compounds present in O. basilicum and its parasit C. campestris. In the present study, eugenol has been isolated from O. basilicum and its parasit C. campestris. But eugenol epoxide has been isolated only from C. campestris. According to literature, some epoxides such as lutein- 5,6-epoxide has been isolated from C. reflexa [20]. The present results confirm the occurrence of enzymatic epoxidation in C. campestris. Here, the anti-HIV-1 activity of eugenol and eugenol epoxide was evaluated for the first time. The results showed that eugenol can increase PBMCs proliferation in a dose dependent method and decrease viral replication. Eugenol epoxide potently inhibits HIV-1 replication without affecting PBMCs proliferation. Eugenol and its derivatives are natural flavonoids which have been identified in various plants, such as basil, cinnamon, lemon balm and clove [21]. There are reports on biological activities of eugenol including antifungal, antibacterial, antioxidant and anti-inflammatory effects [22-24]. Anti- HSV activity of eugenol against HSV-1 and HSV-2 viruses with EC50 of 25.6 μg/ml and 16.2 μg/ml were reported previously [25]. Some studies reported that eugenol could increase lymphocyte proliferation in a dose dependent method [26]. Therefore the ability of lymphocyte proliferation of eugenol may be account for anti-HIV-1 activity. The time of addition study revealed that eugenol was most effective when added 12 and 6 hours pre HIV infection. Pre-incubation of the PBMCs with eugenol may activate proliferation of lymphocyte and prevent HIV-1 replication. Some results demonstrated that stimulation of T cells in vivo can be associated with a decrease in viral replication. Additional evidence suggested that potent T-cell stimulation may enhance production of chemokines and other antiviral cytokines [27-29]. Eugenol epoxide potently cannot affect PBMCs proliferation but inhibits HIV-1 replication. The time of addition study revealed that eugenol epoxide was most effective when added concurrently with HIV- 1 infection. These observations indicated that eugenol epoxide could inhibit the first stage of HIV-1 infection which included attachment of the virus to the cell membrane, diffusion of the virus through the cell membrane and the reverse transcription process. Nevertheless, further studies are needed to verify the mechanism of isolated compounds of C. campestris extract.
Based on the findings of this study, it can be concluded that eugenol and eugenol epoxide isolated from O. basilicum and C. campestris have an inhibitory effect on the initial stage of HIV-1 infection.
This work was supported by the grant from University of Isfahan, Iran. We thank Faculty staff for their research efforts and comments on the study design.
Our work is supported by a Project Grant (189114) in University of Isfahan, Iran.