Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2016) Volume 3, Issue 3
Despite the extensive use of dwarfing rootstocks in commercial apple cultivation worldwide, it is presently unknown how different growing environments may modify the first expression of rootstock-induced dwarfing of the scion. To address this, we measured the growth and architectural development of compound ‘Royal Gala’ apple trees grafted onto three different vigour clonal rootstocks (‘M27’, ‘M9’ and ‘M793’), grown in three locations of New Zealand. Detailed architectural measurements were made over the first year of growth after grafting and the study was repeated for a second year. Within each site and year, trees on ‘M793’ had the greatest primary axis length and node number, trunk cross-sectional area, number of sylleptic shoots and final tree dry weight, while those on ‘M27’ had the smallest. Trees on dwarfing rootstocks generally had a higher frequency of flowering than those on ‘M793’. Our results demonstrate that the effect of dwarfing rootstocks on sylleptic shoot growth, flowering, and dry weight is highly plastic in response to the growing environment. Across all rootstock genotypes, the scion bud type had a strong effect on tree growth: trees that developed from a vegetative scion bud (monopodial primary axis) had a final dry weight 15-45% greater than those with a floral scion bud (sympodial primary axis). One rootstock-induced effect that was consistent between sites and years was that dwarfing rootstocks caused earlier termination of primary axis growth. This provides novel information about the physiological processes that apple dwarfing rootstocks modify.
<Keywords: Apple dwarfing; Rootstock; Primary axis growth; Temperature; Principal component analysis (PCA)
The primary axis of a compound apple tree is derived from a scion bud that is grafted onto the rootstock (Figure 1). When the grafted bud is vegetative, the primary axis develops directly from the apical meristem. In the case of a mixed floral bud, the apical meristem terminates in an inflorescence and the axis development is continued by outgrowth of a sub-terminal axillary meristem. In plant architecture, these two types of axis development are referred to as “monopodial” and “sympodial” respectively [1,2]. In apple, a vegetative bud contains 9-11 preformed nodes [3,4] whereas a floral bud has only 5-6 preformed vegetative nodes [5,6], which may affect the onset and vigour of primary axis development. Anecdotally, it is thought that grafting scion wood comprised mixed floral buds limits young tree growth and therefore final tree size in the first year of growth after propagation. However, no previous studies have determined how grafted bud type may quantitatively modify scion architecture and whole tree dry mass gain.
During the first season of growth after grafting, axillary meristems along the primary axis either grow out to form a sylleptic shoot [7], or develop into either a floral bud or a vegetative bud that remains dormant until spring of year two (Figure 1). Apple dwarfing rootstocks reduce tree size by altering the development of axillary meristems, which affects the type of growth units that develop during the following season. The earliest effect of the ‘Malling 9’ (‘M9’) dwarfing rootstock is a decrease in the number, length and node number of sylleptic shoots that develop within the first year after grafting [8-10] and/or an increase in the proportion of floral buds along the primary axis in the second spring [11,12]. More floral buds results in fewer and shorter extension growth units in the next growth cycle [13]. Rootstock-induced dwarfing is manifested within the first year of growth and becomes more pronounced over successive growth cycles. After several years, trees grafted onto ‘M9’ had a canopy size half that of the same scion grafted onto the semi-vigorous ‘MM106’ rootstock [14].
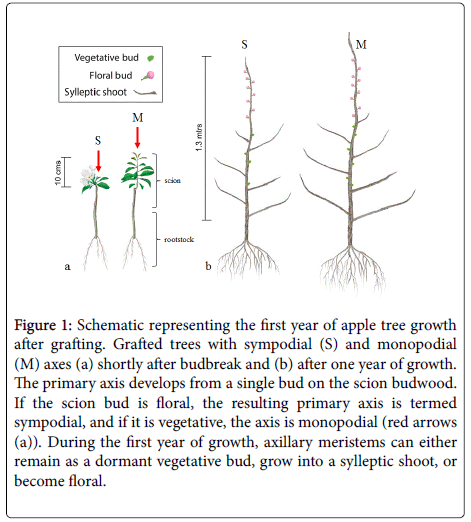
Figure 1: Schematic representing the first year of apple tree growth after grafting. Grafted trees with sympodial (S) and monopodial (M) axes (a) shortly after budbreak and (b) after one year of growth. The primary axis develops from a single bud on the scion budwood. If the scion bud is floral, the resulting primary axis is termed sympodial, and if it is vegetative, the axis is monopodial (red arrows (a)). During the first year of growth, axillary meristems can either remain as a dormant vegetative bud, grow into a sylleptic shoot, or become floral.
For genetically identical apple scion/rootstock combinations, the scion phenotype that develops in the first year after grafting of the compound tree can differ markedly depending on growing season and/or environment. For example, in the humid climate of the Manawatu, ‘Royal Gala’ scions on ‘M9’ typically developed a primary axis that was shorter because of fewer nodes, and developed fewer sylleptic shoots than scions on more vigorous rootstocks [8,9]. In contrast, these effects of ‘M9’ were not observed for the same rootstock/scion combinations grown in the warmer and drier climate of Hawke’s Bay, where the final length and node number of the primary axis were similar irrespective of rootstock vigour [13]. Thus, it appears that the initial changes in scion architecture on a given rootstock genotype can differ between growing environments, indicating there is a degree of plasticity concerning how rootstockinduced dwarfing is first expressed. Identification of the architectural traits that are common across environments is required to further elucidate the physiological processes that dwarfing rootstocks modify.
To date, most studies investigating how dwarfing rootstocks first modify scion architecture after grafting of the composite tree have focused on ‘M9’ [8,9,11,13-16], thus it is presently unknown if other dwarfing rootstock genotypes modify scion architecture in the same way. The ‘Geneva 16’ dwarfing apple rootstock was reported to enhance axillary shoot development of ‘Brookfield Gala’ scions [17], which is in contrast to findings for ‘Royal Gala’ scions on ‘M9’ which developed few axillary shoots [15,18]. These disparities in scion phenotype suggest that other dwarfing rootstock genotypes might impose dwarfing by some slightly different mechanisms than that of ‘M9’. Alternatively, different rootstock effects on young tree architectural development in these previous studies may be due to disparities of tree culture, growing environments and their putative interactions.
We included the “very dwarfing” ‘M27’ and dwarfing ‘M9’ in this present study to further understand how these dwarfing rootstock genotypes modified scion architecture in the first year following tree grafting compared with the vigorous ‘M793’ rootstock. Compound trees were grown in three unique growing environments of New Zealand, with the experiment repeated in two different growing seasons, to identify the common architectural trait(s) that first appear causal in rootstock-induced scion dwarfing. Within each site and year, newly grafted trees were grown using the same growing medium and watering regimes, thereby standardising tree culture to lessen architectural variability due to non-genetic causes. Differences in tree culture may explain different timings and the degree dwarfing expression reported previously [8,9,11,13-16]. A further important cultural practice not considered in the methodology of previous work is that of the grafted bud type, which if not standardised, has potential to confound young tree vigour comparisons among different rootstock genotypes.
In summary, the aims of the study were to: 1) Quantify how two different dwarfing rootstock genotypes modified scion architecture in the first year after grafting compared with the vigorous ‘M793’ rootstock; 2) Determine scion architectural trait(s) consistently causal in initial scion dwarfing by ‘M27’ and ‘M9’ across different environments and years; and 3) Determine whether grafted bud type (vegetative vs. floral) modifies scion architecture and whole tree dry mass gain.
Plant material and phenotyping
Malus × domestica ‘Royal Gala’ scions were cleft grafted at a height of 35 cm onto 1-year-old clonal rootstock stools of ‘M793’, ‘M9’ (clone ‘NZ9’) and ‘M27’ rootstocks (Waimea Nursery, Nelson, New Zealand) in August (winter) of 2011 and 2012. Grafted trees were planted into 50-L bags containing growing medium; 0.8 bark: 0.2 pumice (by volume) containing: 1 gL-1 of 6-month slow-release fertilizer, 3 gL-1 of 9-month slow-release fertilizer (each 13N-5.7P-10.8K-1.2 Mg) (Nutricote®, Chisso-Asahi Fertilizer Co., Japan), and 0.6 gL-1 Magri- Trace (15Mg-3.2Ca-18Fe-3Mn-4Zn-3Cu-0.6B,0.08Mo) (HortFert plus Ltd, NZ). The distance between the soil level and graft junction was kept constant at 15 cm. Potted trees were placed 0.8 m apart along nursery rows, in a randomized block layout. The irrigation system consisted of a 19 mm polytube line to which pressure-compensating drippers were attached (one 4 Lh-1 dripper per tree). Irrigation was scheduled daily for 30 min at dawn and dusk using an automated time controller. At budbreak in mid-September, scions were de-budded to a single axis and thereafter received no pruning. Grafted trees were grown at three different Plant & Food Research sites in New Zealand: Havelock North (39.67°S, 176.88°E), Palmerston North (40.36°S, 175.61°E), and Motueka (41.12S, 173.01E).
In both years, 20 composite trees grafted to each rootstock genotype were grown at each of the three sites with similar irrigation and cultural management. Trees damaged by insects, animals or disease were excluded from analysis. The number of trees phenotyped for specific traits is shown in Supplemental Table 1. In year one (2011–12), no consideration was given to the type of scion bud (vegetative vs . floral) grafted onto the rootstocks, and the ‘M27’ rootstocks were slightly larger (10–12 mm) than the ‘M9’ and ‘M793’ rootstocks (8–10 mm). Scion budwood containing vegetative buds tends to be larger in diameter because vegetative buds develop at the base, with floral buds being more distal. In year two (2012–13), we ensured that there were equal numbers of trees with sympodial and monopodial primary axes for each rootstock genotype and that the diameter of both scion budwood and all rootstocks was between 8 mm-10 mm at the time of grafting.
The length and node number of the primary axis and the position, length and node number of sylleptic shoots were recorded approximately every two weeks from spring until the end of the first growing season. After one year of growth, the diameter (D) of the primary axis was measured 20 cm above the graft union. Trunk crosssectional area (TCA) was calculated as π(D/2)2. In June (beginning of winter) of 2013, a subset of 10 trees per genotype, which included five sympodial and five monopodial primary axes, were moved into a cool store at 4°C for 8 weeks to ensure sufficient chilling for flowering and to avoid Pseudomonas infection that can kill overwintering potted apple trees in New Zealand conditions. For all three sites, axillary floral buds along the primary axis were counted in October of year two (2013). For each tree, the number of floral buds was calculated as a percentage of the number of available nodes, i.e., the number of nodes on the primary axis minus the number of sylleptic shoots.
At 300 days after budbreak (DABB), trees from Havelock North and Palmerston North were destructively harvested and oven dried at 60°C to a constant mass before the dry weight of the roots, rootstock shank (stem), scion budwood, primary axis and sylleptic shoots were measured separately. Comparisons of architectural measurements are relative to the vigorous ‘M793’.
Automated weather stations at each site recorded hourly measurements. Growing degree days (GDD) were computed from the average daily temperature (calculated from 24 hourly temperature measurements), with a base temperature of 10°C. Thus, a daily average of 8°C contributes nothing to GDD, rather than -2°C. Daily wind speed (km/h) was calculated from the average of 24 h measurements and the diurnal temperature variation was the daily maximum minus the daily minimum. Accumulated daily wind speed and diurnal temperature variation were used for the principal component analysis. The graphs shown in Supplemental Figure S3 were averaged over 5 days. We excluded rainfall and humidity from our analysis because the trees were under non-limiting watering regimes.
Dry weights were fitted to linear mixed models and comparisons between monopodial and sympodial axes were made using Fisher’s least significant difference test between means (GenStat 17th edition software (VSN international, Hemel Hempstead, UK)). Architectural data from 20 replicates per rootstock genotype, three sites and two years were fitted to linear mixed models using rootstock, site, and year as factors (GenStat). Pairwise comparisons between dwarfing and vigorous rootstocks (Figure 3), monopodial and sympodial axes (Supplemental Table 2), and between sites for each rootstock and year (Supplemental Table 3) were made using Fisher’s least significant difference test between means. OriginLab 8.5 (Northhampton, MA) and Excel 2013 (Microsoft Corporation, Redman, WA) were used for graphing. Primary axis node appearance rate was calculated for each tree by dividing the difference in node number by the days between measurements. To calculate node appearance rate as a function of thermal time, the primary axis node number was plotted as a function of GDD for each tree. Exponential curves (in the form of y=A+B*Rx) were fitted to data using parameters estimated by maximum likelihood using GenStat. Node appearance rate values were derived from the fitted curves and are expressed as node appearance per 100 GDD. Principal component analysis was performed on architectural and weather data from all sites, both years and plotted by rootstock genotype using R 3.0.1 [19] and a script created by Dr Marcus Davy.
Expression of rootstock-induced dwarfing in different growing environments
In two consecutive years, final dry weights were measured at 300 DABB from trees grown in Havelock North and Palmerston North. In both sites and both years, trees grafted onto ‘M793’ had the highest scion and rootstock dry weight, while those grafted onto ‘M27’ generally had the lowest (Figures 2a and 2b). Trees with a monopodial primary axis (originating from a vegetative scion bud) accumulated more scion and rootstock dry weight than those with a sympodial primary axis on the same rootstock. The differences in dry weight between axis types were more pronounced in year two, when the total tree dry weights of monopodial axes were 15–45% greater than those of sympodial axes on the same rootstock and location. The site had a strong effect on dry weight, especially in year two when trees in Havelock North had nearly twice the total dry weight of comparable trees grown in Palmerston North (Figure 2c).
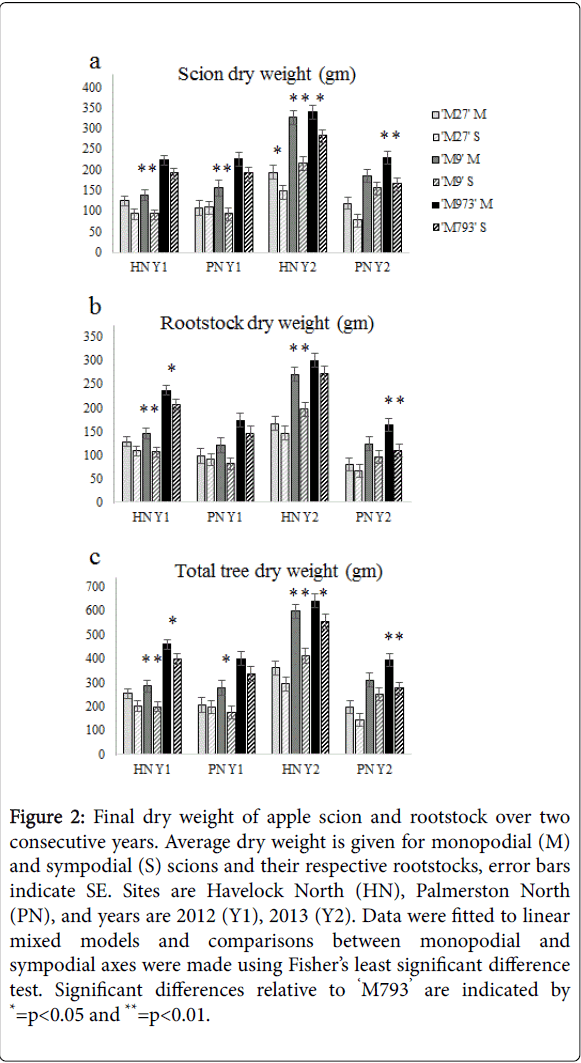
Figure 2:Final dry weight of apple scion and rootstock over two consecutive years. Average dry weight is given for monopodial (M) and sympodial (S) scions and their respective rootstocks, error bars indicate SE. Sites are Havelock North (HN), Palmerston North (PN), and years are 2012 (Y1), 2013 (Y2). Data were fitted to linear mixed models and comparisons between monopodial and sympodial axes were made using Fisher’s least significant difference test. Significant differences relative to ‘M793’ are indicated by *=p<0.05 and **=p<0.01.
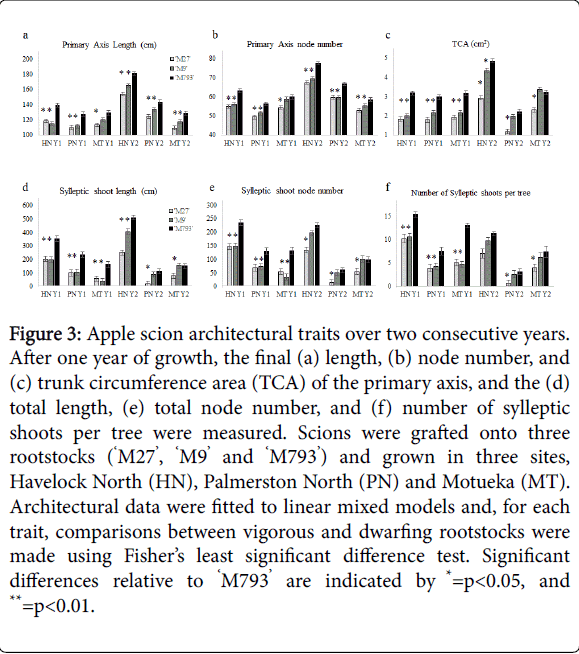
Figure 3: Apple scion architectural traits over two consecutive years. After one year of growth, the final (a) length, (b) node number, and (c) trunk circumference area (TCA) of the primary axis, and the (d) total length, (e) total node number, and (f) number of sylleptic shoots per tree were measured. Scions were grafted onto three rootstocks (‘M27’, ‘M9’ and ‘M793’) and grown in three sites, Havelock North (HN), Palmerston North (PN) and Motueka (MT). Architectural data were fitted to linear mixed models and, for each trait, comparisons between vigorous and dwarfing rootstocks were made using Fisher’s least significant difference test. Significant differences relative to ‘M793’ are indicated by *=p < 0.05, and **=p < 0.01.
Final primary axis length and node number, trunk circumference area (TCA), the number, length and node number of sylleptic shoots per tree varied greatly between years, among sites and rootstock genotype treatments (Figure 3). Overall, trees in Havelock North grew the most, especially in year two. At each site, the trend between rootstock effects was the same: ‘M793’ had the highest average primary axis length and node number, TCA, and sylleptic shoot number; and ‘M27’ had the lowest. For each rootstock, differences in architectural traits among sites were greater in year 2 (Supplementary Table S2). Monopodial primary axes tended to be longer with more nodes those than sympodial primary axes (Supplementary Table S3).
Zones of sylleptic branching and flowering on the primary axis
In spring of year two, we compared the relative frequency of branching and flowering at each node along the primary axis (Figure 4). In all sites, sylleptic shoots developed towards the base of the primary axis, and flowers more distally. Trees in Havelock North had the most sylleptic shoots and those in Palmerston North the least. The Palmerston North site trees had the largest number of nodes that did not develop into either sylleptic branches or floral buds during the first year of growth (Supplemental Figure S1). At each site, the zone of branching tended to extend further up the primary axis and be more frequent in monopodial axes and with increasing rootstock vigour. Conversely, the zone of flowering tended to extend more basally in sympodial primary axes and be more frequent with decreasing rootstock vigour. All trees flowered, however the percentage of floral buds per available node ranged from 40% to 51% for scions on the dwarfing rootstocks, compared with 20–35% for scions on ‘M793’ (Supplemental Figure S2).
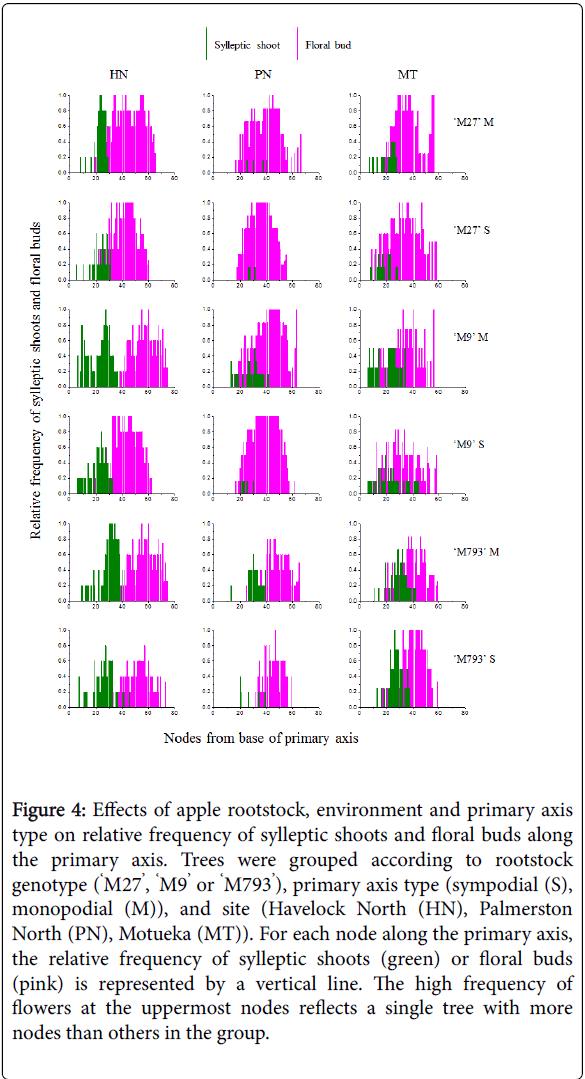
Figure 4: Effects of apple rootstock, environment and primary axis type on relative frequency of sylleptic shoots and floral buds along the primary axis. Trees were grouped according to rootstock genotype (‘M27’, ‘M9’ or ‘M793’), primary axis type (sympodial (S), monopodial (M)), and site (Havelock North (HN), Palmerston North (PN), Motueka (MT)). For each node along the primary axis, the relative frequency of sylleptic shoots (green) or floral buds (pink) is represented by a vertical line. The high frequency of flowers at the uppermost nodes reflects a single tree with more nodes than others in the group.
Principal Component Analysis of factors influencing variation in phenotype
During the two years of this study, Havelock North had the highest accumulated temperature (GDD), Palmerston North had the highest wind speed and lowest diurnal temperature variation, and Motueka had the lowest GDD and wind speed and the highest diurnal temperature variation (Supplemental Figure S3).
Principal component analysis (PCA) was used to examine relationships between the architectural measurements and weather data for all sites and both years (Figure 5). About half the variation in data was explained by PC1 (51%), which indicates that GDD was positively associated with all aspects of tree growth. PC2 explained about a quarter of the variation in data (24.7%) and was largely comprised of wind, diurnal temperature variation and GDD
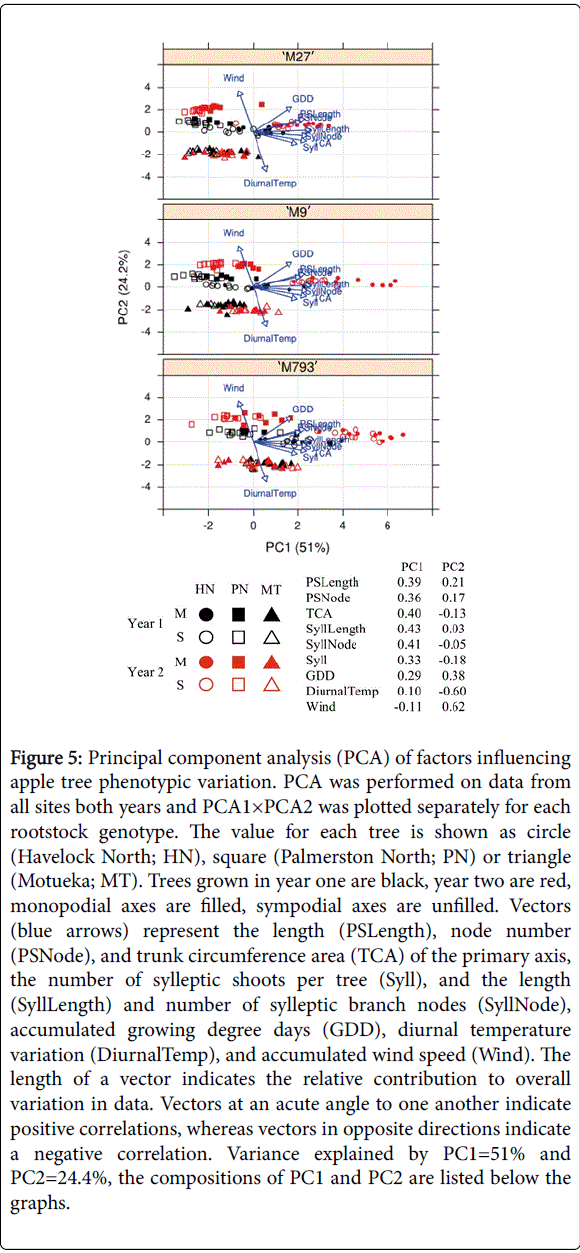
Figure 5: Principal component analysis (PCA) of factors influencing apple tree phenotypic variation. PCA was performed on data from all sites both years and PCA1×PCA2 was plotted separately for each rootstock genotype. The value for each tree is shown as circle (Havelock North; HN), square (Palmerston North; PN) or triangle (Motueka; MT). Trees grown in year one are black, year two are red, monopodial axes are filled, sympodial axes are unfilled. Vectors (blue arrows) represent the length (PSLength), node number (PSNode), and trunk circumference area (TCA) of the primary axis, the number of sylleptic shoots per tree (Syll), and the length (SyllLength) and number of sylleptic branch nodes (SyllNode), accumulated growing degree days (GDD), diurnal temperature variation (DiurnalTemp), and accumulated wind speed (Wind). The length of a vector indicates the relative contribution to overall variation in data. Vectors at an acute angle to one another indicate positive correlations, whereas vectors in opposite directions indicate a negative correlation. Variance explained by PC1=51% and PC2=24.4%, the compositions of PC1 and PC2 are listed below the graphs.
The combination of PC1 and PC2 reflected the environmental factors that represent the growing conditions for each site and year. The three sites were clearly separated from one another, especially for trees on ‘M793’. The variation between years was greatest for Havelock North, because of a higher GDD in year 2. In Palmerston North, wind contributed to a significant difference between years. There was very little variation between years in Motueka, in terms of both architectural and weather data. Trees on ‘M793’ showed a wider variation in phenotype than those on the dwarfing rootstocks.
In general, vectors representing aspects of primary axis and sylleptic shoot growth were closely aligned. The vectors representing primary axis length and node number were nearly superimposed, indicating that these traits are closely associated. The acute angle between vectors suggests that sylleptic shoot number is more influenced by diurnal temperature variation than other aspects of scion growth such as primary axis length or node number.
Primary axis growth rate was dependent on temperature and rootstock genotype
In this study, primary axis growth rate is defined as the rate of node appearance. The rate of node appearance on the primary axis was calculated for each tree over the first year of growth (Figure 6). For both years and all rootstock genotypes, the highest growth rates occurred in the first 150 DABB. The pattern of growth rates was similar between sites in year one, but in year two, trees in Havelock North exhibited a spike in growth rate around 100 DABB. In all sites and both years, trees with ‘M793’ rootstocks had a higher growth rate later in the season than those on ‘M9’ and ‘M27’.
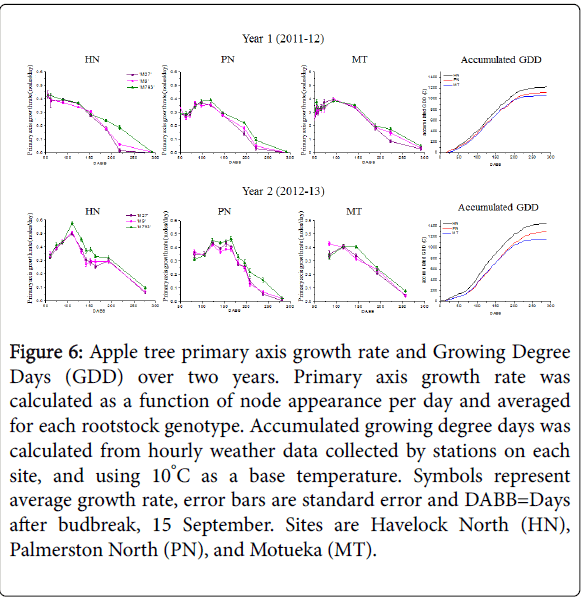
Figure 6: Apple tree primary axis growth rate and Growing Degree Days (GDD) over two years. Primary axis growth rate was calculated as a function of node appearance per day and averaged for each rootstock genotype. Accumulated growing degree days was calculated from hourly weather data collected by stations on each site, and using 10°C as a base temperature. Symbols represent average growth rate, error bars are standard error and DABB=Days after budbreak, 15 September. Sites are Havelock North (HN), Palmerston North (PN), and Motueka (MT).
Year one was cooler and there was little difference in final GDD between sites (Figure 6 and Supplemental Figure S3). Year two was much warmer, especially for Havelock North, which experienced a rapid increase in GDD between 75DABB-125 DABB. The period of increased temperature corresponded with the spike in growth rate observed in Havelock North in year two, consistent with our observation that primary axis growth rate is highly influenced by temperature.
To examine the relationship between primary axis growth rate and temperature, node number was plotted as a function of GDD for each tree. Exponential curves (in the form of y=A+B*Rx) were fitted to the data using parameters estimated by maximum likelihood. Growth rate values were derived from the fitted curves and are expressed as node appearance per 100 GDD.
When expressed as a function of GDD, the growth rate of each genotype was similar across all sites (Figure 7 and Supplemental Figure S4). Data for year one and two were plotted separately because of the variation in initial budwood diameter observed in year one. Both years showed the same trend: Primary axis growth rate was highest early in the season, with no clear difference between rootstock genotype. As GDD increased, scions on ‘M793’ maintained a higher growth rate than those on dwarfing rootstocks.
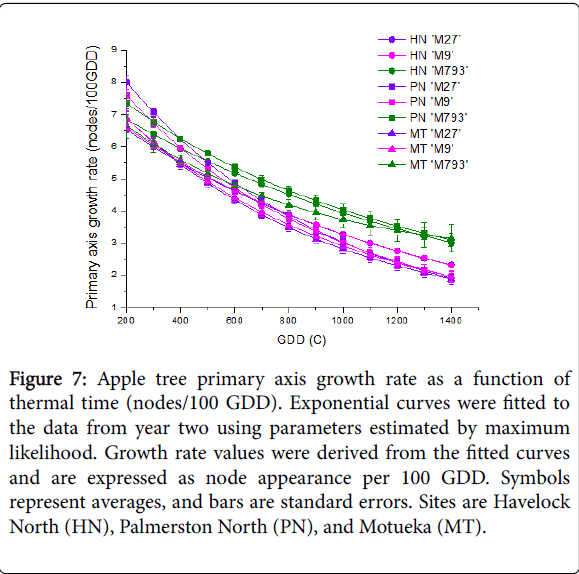
Figure 7: Apple tree primary axis growth rate as a function of thermal time (nodes/100 GDD). Exponential curves were fitted to the data from year two using parameters estimated by maximum likelihood. Growth rate values were derived from the fitted curves and are expressed as node appearance per 100 GDD. Symbols represent averages, and bars are standard errors. Sites are Havelock North (HN), Palmerston North (PN), and Motueka (MT).
Within a given site/year, there was a consistent trend in which trees on ‘M793’ were the largest in terms of primary axis length and node number, stem diameter, number and length of sylleptic shoots and tree dry weight, while trees on ‘M27’ were the smallest. However, the magnitude of rootstock-induced effects varied widely between sites and years, demonstrating a strong influence of growing environment and scion bud type on rootstock-induced dwarfing. For example, in year two of our study, the average whole tree dry weight of monopodial axes on the very dwarfing ‘M27’ in Havelock North was 31.3% greater than that of sympodial axes on the invigorating ‘M793’ in Palmerston North. The absolute effects of rootstocks are relative and cannot be directly compared between sites or years. This has implications for studies in which extend from year to year or have multiple sites or growth conditions. When assessing or comparing rootstock effects, it is essential to compare trees grown under the same growth conditions, with the same scion genotype and bud type.
There have been studies of rootstock effects across different environments [20,21], but these have measured field-grown trees after five to ten years in variable soil conditions, which complicates the analysis of environmental effects on early scion growth. In these studies, trees flowered and bore fruit which has been demonstrated to effect subsequent growth [13]. The present study is the first to demonstrate the effect of environment and scion bud type on the first expression of rootstock-induced dwarfing.
Effect of scion bud type on shoot and root growth
The extent to which the scion bud type affected the growth of both scion and rootstock was greater than anticipated. The differences in final primary axis length and node number between monopodial and sympodial primary axes correspond closely with the difference in preformed node number between vegetative and floral buds [4,6]. It is less clear how the scion bud type would affect scion and rootstock dry weight or the number and length of sylleptic shoots.
One explanation might be differences in the initial growth capacity of vegetative vs. floral apple buds. At budbreak, a vegetative bud has twice as many preformed nodes and a much larger shoot apical meristem than an axillary meristem that develops into a sympodial axis. A larger meristem is capable of increased rates of leaf initiation [22], which would lead to higher leaf area and increased dry matter accumulation in newly grafted trees. As a result, the developments of sympodial axes are initially delayed by about 7–10 days relative to the monopodial axes. For a given rootstock/environment treatment, both primary axis types terminate at the same time, which effectively shortens the duration of growth for sympodial axes and reduces overall growth.
Axillary sylleptic shoots vs. floral buds
All rootstock/axis types showed the same pattern of sylleptic branching at the base of the primary axis, and flowering being more distal, but the relative size of these two zones varied considerably between rootstock genotypes and sites. In general, monopodial axes on ‘M793’ had the highest frequency of sylleptic shoots and the lowest flowering frequency. Dwarfing rootstocks have been reported to increase precocity [23]. In our study, all trees flowered in spring of year two, indicating no rootstock effect on precocity.
Regardless of rootstock genotype, trees in Havelock North exhibited the most sylleptic growth and Palmerston North trees the least, which reflects the impact of environmental factors on this trait. Our observations are consistent with previous reports that sylleptic branching is highly variable and influenced by genotype and environment [24-26].
The balance between sylleptic shoots and axillary flowers greatly influences subsequent growth patterns. Sylleptic shoots provide additional leaf area and photosynthetic capacity during the growing season and translocate a larger proportion of carbon to the stem than proleptic shoots that grow out after a dormancy period [27]. Increased carbon to the root system promotes strong growth in year two. In contrast, the increase in axillary flowers initiated in year one results in fewer extension units in year two [13]. These early differences in growth are compounded over subsequent growing cycles.
Impact of environment on growth
Nearly a quarter of the variation in data was explained by PC2, which was comprised of a combination of wind, diurnal temperature variation and GDD that distinguishes each site and year. With the current data, it is not possible to quantify the impact of each environmental factor on a given architectural trait. However, trends between sites and years allow some conclusions about environmental effects on primary axis and sylleptic shoot growth.
Accumulated temperature (GDD) was positively associated with growth of both the primary axis and sylleptic shoots. Trees in Havelock North were exposed to the highest GDD and had the largest length and node number of both primary axes and sylleptic shoots and the greatest overall dry weight. Because of the importance to world food production, the impact of increased temperature on arable crops has been a focus of numerous studies [28-30]. Plant growth and productivity increases with higher temperature, up to a point, after which there is a sharp decline in productivity. A wide survey of mostly forestry trees found that deciduous trees are more sensitive to changes in temperature than evergreen trees, and tropical trees have a negative growth response to increased temperatures (Way and Oren 2010). The relationship between temperature and primary axis length has also been demonstrated in some fruit trees and vines [31-33], but never for compound trees grafted onto different rootstocks.
Palmerston North was characterized by high wind and low diurnal temperature variation, whereas Motueka was the opposite. The combination of diurnal temperature variation and wind had a stronger effect on sylleptic shoot growth than GDD. For example, trees in Palmerston North had fewer and shorter sylleptic shoots than did trees in Motueka, despite the former having a higher GDD in both years. Our observations support the theory that sylleptic branching is a very plastic trait that enables trees to respond to current conditions and grow more or less accordingly [34]. Final TCA was also increased by high diurnal temperature variation and low wind. The influence of night temperature on TCA has been documented in a number of trees [31].
Although we cannot separate wind from diurnal temperature in this study, we speculate that high wind in Palmerston North negatively affected sylleptic shoot development. Previous studies have reported that increased wind results in reduced primary axis and secondary shoot growth in both annual and perennial species [35-39]. Wind causes mechanical stress to leaves and stems and has been shown to increase ethylene synthesis [40,41] and the activity of protein kinases involved with wounding responses [42,43]. Physical disturbance also reduces phloem transport, which could alter the movement of carbohydrate, hormones and other signalling molecules [44,45]. To avoid dehydration, plants respond to wind by closing stomata, which would reduce CO2 uptake and photosynthesis.
Primary axis growth rate is dependent on accumulated temperature and rootstock genotype
To account for differences in accumulated temperature between sites, primary axis growth rate was modelled as a function of GDD. For all rootstocks, primary axis growth rate was dependent on GDD. Scions on both dwarfing rootstocks showed similar growth rates, but those on ‘M793’ had a higher growth rate, especially later in the season. Another way of interpreting this is that the primary axes on dwarfing rootstocks terminate growth earlier in the season and therefore have a shorter duration of growth in warmer growing seasons than those on ‘M793’. Low vigour or dwarfing rootstocks in kiwifruit and cherry are also associated with earlier termination of the primary axis [46,47]. Earlier termination on dwarfing rootstocks was consistent across all sites, suggesting that this trait is largely under genetic control. Other traits, such as sylleptic shoot growth, flowering and final dry weight, are controlled by a combination of rootstock genotype and growing conditions.
The first expression of rootstock-induced dwarfing exhibits a high degree of plasticity in response to growing environment, growing season and the type of scion bud grafted. Understanding that these factors can profoundly modify young tree vigour and architecture is highly relevant to both growers and rootstock breeders. For example, grafting vegetative scion buds onto any rootstock will produce a significantly larger tree in the first growing season, which improves nursery tree quality and profitability. For breeders, when assessing or comparing rootstock effects on scion vigour, precocity and flowering intensity, it is essential to compare trees grown under the same growth conditions, with the same grafted scion bud type, and across more than one growing season, to ensure accurate phenotyping. Our work demonstrates that earlier primary axis and/or sylleptic shoot termination is the most consistently expressed manifestation imposed by ‘M9’ and ‘M27’ dwarfing rootstocks across different growing environments and years. This provides new phenotypic information for future studies elucidating the physiological and genetic bases for apple rootstock-induced dwarfing.
We thank Amy Watson, Robert Diack, Daya Dayatilake and Chethi DeZoysa for data collection, Duncan Hedderley for statistics assistance, Marcus Davey and Peter McAtee for tutoring with R, and Tony Corbett for the illustrations in Figure 1.