Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 2
We applied a modified cDNA-AFLP technique by using cassava leaf samples under drought treatment, and obtained over 100 transcript-derived fragments (TDFs) which showed significant differentially expressed during the process of drought treatment and 63 TDFs of them were cloned, sequenced and annotated. The results suggested that the recognized site of MmeI enzyme was integrated into double-strand cDNAs, and the 5’ ends of TDFs that got by the modified technique were nearer to the transcript start site comparing with the previous method. Furthermore, most differentially expressed TDFs (DE-TDFs) really up-regulated or down-regulated through qRT-PCR validation and the homologies of part TDFs involved in or responded to drought tolerance. In general, the modified cDNA-AFLP technique has been established preliminary, and the genetic information of these TDFs needs further mining.
Keywords: Cassava; Drought; cDNA amplified fragment length polymorphism; Transcript-derived fragment
cDNA-AFLP: cDNA Amplified Fragment Length Polymorphism; TDF: Transcript-Derived Fragment; Qrt-PCR: Quantitative Reverse Transcript Polymerase Chain Reaction; TSS: Transcription Start Sites; M: Mmei; E: Ecori; B: Bstyi; ME, MM, MB, BE, EE: The Primers Pairs Basing On The Adapters Of Different Enzyme Combination
Cassava (Manihot esculenta Crantz) was a woody shrub of the Euphorbiaceae, originated from tropical America, called one of the three biggest tubers along with potato and sweet potato. It was an important staple crop worldwide and consumed by 600 million people. It could grow on very poor soils under prolonged drought for more than six months, reduced its leaf canopy and transpiration water loss, but its attached leaves remained photosynthetically active, though at greatly reduced rates and this drought tolerance mechanism led to high crop water use efficiency values [1]. The cDNA amplified fragment length polymorphism (cDNA-AFLP) technique was a large-scale detected technology of differential gene expression, and didn’t need the information of genome sequence [2,3]. Since its appearance, it was widely applied in disease resistance, nutrition stress, specific developmental stage and organ, etc, obtained many valued transcript-derived fragments (TDFs), and they had highly homologous to functional genes or unknown genes [4-7]. In order to get more information by cDNA-AFLP, some researcher optimized its enzyme combinations by AFLP inSilico, and found that BstYI/MseI was optimal restriction enzyme combination, could obtain more than 60% of all transcripts in tobacco [8]. In this study, a modified cDNA-AFLP technique would be established and used to screen the drought-stress induced genes in cassava. We applied the SMART™ technology to synthesize single- and double-strand cDNA, and integrated the recognized sequence (TCCRAC) of restriction enzyme MmeI in the 5’end of the synthesized double-strand cDNA, anchored a cutting site at the transcription start sites (TSS). Then, most TDFs that drought stress induced would start at the TSS and easy to obtain their full-length cDNAs in cassava.
Plant materials
The cassava varieties SC124 and KU50 were preserved in cassava germ plasma of Chinese Academy of Tropical Agricultural Sciences in Wenchang City of Hainan Province, China. The potted seedling of SC124 and KU50 were placed in greenhouse with soil maximum moisture capacity of 90% and fertilized with Hoagland’s solution [9]. After 2 months of planting, the uniform potted seedlings were subjected to drought stress treatment. The about 40 uniform potted seedlings were subjected to two types of drought treatment: 1) Drought treatment (DT): no water until an obvious symptom of drought appearing, total RNAs were collected at 6d, 8d, 10d and 12d after beginning drought treatment; 2) normal condition (NC): watering to soil maximum moisture capacity of 90% every day, total RNA was collected at 0d, 6d and 12d. All RNA samples were extracted from the leaves two individuals and used for cDNA-AFLP analysis, and further quantitative RT-PCR assays.
Modified cDNA-AFLP technique
Double-strand cDNA synthesis: The restriction enzyme MmeI was a key player in this modified cDNA-AFLP technique, and its cutting site was back 20bp to C of the recognized site on forward strand, and back 18bp on reverse strand (Figure 1A). Refer to SMART™ cDNA Library Construction Kit (Clontech, BD, USA), its 5’ SMART IV™ Oligonucleotide primer was changed and introduced a Mme ? Recognized site TCCGAC in it, now its sequence was AAGCAGTTCCGACTGGTATCAACGCAGAGTggg-3’ (Figure 1B), and the SMART CDS III/3’ PCR primer was simplified to dT30N1N2 (N1=A, G, C; N2=A, T, C, G). Then the double-strand cDNA was synthesized with these two primers (Figure 1C and D).
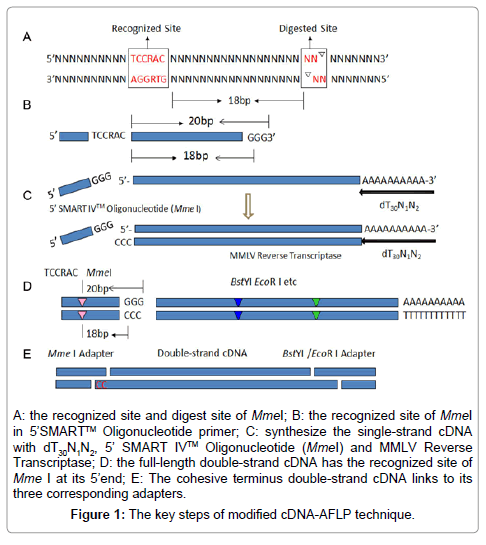
Figure 1: The key steps of modified cDNA-AFLP technique.
Double-strand cDNA digest
Three restriction enzymes, MmeI, EcoRI and BstYI, were used to digest double-strand cDNA, and the reaction system was follow: double-strand cDNA 10μl, NEB buffer4 2.0 μl, MmeI 1.0 μl, EcoRI 1.0 μl, BstYI 0.5 μl, ddH2O 5.5 μl, total 20.0 μl. The reaction system was incubated at 37°C for 2 h, 60°C for 1h, and 80°C for 20min to inactivate enzyme activity in water-bath.
Adapter ligation, pre- and selective amplify
Digested double-strand cDNA 10.0 μl, 10 × T4 DNA ligase buffer 2.0 μl, T4 DNA ligase 1.0 μl, MmeI adapter (50 μM) 1.0 μl, EcoRI adapter (25 μM) 0.5 μl, BstYI adapter (25 μM) 0.5 μl, and ddH2O 3.0 μl. The ligation system was incubated at 16°C overnight in water-bath (Figure 1E).
Mme I adapter: 5’-GTCCTCACAACGATTCCACAGG-3’
3’-CAGGAGTGTTGCTAAGGTGT-5’
EcoR I adapter: 5’-CTCGTAGACTGCGTACC-3’
3’-CATCTGACGCATGGTTAA-5’
BstY I adapter: 5’-CCGCGTTAACCGAGAT Pu-3’
3’-GGCGCAATTGGCTCTAPyCTAG-5’
The ligation products were diluted 50 times as template for preamplification. The pre-amplification with non-selective nucleotide primer pairs was performed for 26 cycles with the cycle profile: a 30 s DNA denature step at 94°C, a 30 s annealing step at 56°C, and a 60 s extension step at 72°C. The selective amplification with three selective nucleotides prime pairs was performed for 36 cycles with the cycle profile: a 30 s DNA denature step at 94°C, a 30 s annealing step, and a 60 s extension step at 72°C. The anneal temperature at the first cycle was 57°C, was subsequently reduced each cycle by 0.3°C for the next 10 cycles, and was continued at 54°C for the remaining 26 cycles. All amplification reactions were performed in a Biometra Thermocycler (Biometra Corp, Göttingen, GER). The primers of the three enzymes for pre- and selective PCR amplification were list in Table 1.
| Primers Code | Sequence (5’-3’) | Primers Code | Sequence (5’-3’) |
|---|---|---|---|
| M Mtgc Mtga Mtac Mtag Mtat Mttg Mtta Mtcg Mtca Mcta Mcat Matc Mact B Baagt Baagc Bagac Bagca Bgagc Bgagt Bggac Bggca E Egtc Egtg Eacg Etgc Egag Egca Ecgt Ectc |
ACAACGATTCCACAGG ACAACGATTCCACAGGtgc ACAACGATTCCACAGGtga ACAACGATTCCACAGGtac ACAACGATTCCACAGGtag ACAACGATTCCACAGGtat ACAACGATTCCACAGGttg ACAACGATTCCACAGGtta ACAACGATTCCACAGGtcg ACAACGATTCCACAGGtca ACAACGATTCCACAGGcta ACAACGATTCCACAGGcat ACAACGATTCCACAGGatc ACAACGATTCCACAGGact CGTTAACCGAGATPuGATCPy CGTTAACCGAGAT(a/t)GATC(t/a)AGT CGTTAACCGAGAT(a/t)GATC(t/a)AGC CGTTAACCGAGAT(a/t)GATC(t/a)GAC CGTTAACCGAGAT(a/t)GATC(t/a)GCA CGTTAACCGAGAT(a/t)GATC(t/a)AGC CGTTAACCGAGAT(a/t)GATC(t/a)AGT CGTTAACCGAGAT(a/t)GATC(t/a)GAC CGTTAACCGAGAT(a/t)GATC(t/a)GCA GACTGCGTACC AATTC GACTGCGTACCAATTCgtc GACTGCGTACCAATTCgtg GACTGCGTACCAATTCacg GACTGCGTACCAATTCtgc GACTGCGTACCAATTCgag GACTGCGTACCAATTCgca GACTGCGTACCAATTCcgt GACTGCGTACCAATTCctc |
Egcg Egac Ecat Ecag Ecaa Eccg Eccc Ecct Ecca Ecgg Ecgc Ectt Ecta Ectg Ectc Ecga Eata Etat Eact Eaca Eagt Eatc Eatg Eaga Egat Egta Etac Etag Etga Etgt Eagc Egct |
GACTGCGTACCAATTCgcg GACTGCGTACCAATTCgac GACTGCGTACCAATTCcat GACTGCGTACCAATTCcag GACTGCGTACCAATTCcaa GACTGCGTACCAATTCccg GACTGCGTACCAATTCccc GACTGCGTACCAATTCcct GACTGCGTACCAATTCcca GACTGCGTACCAATTCcgg GACTGCGTACCAATTCcgc GACTGCGTACCAATTCctt GACTGCGTACCAATTCcta GACTGCGTACCAATTCctg GACTGCGTACCAATTCctc GACTGCGTACCAATTCcga GACTGCGTACCAATTCata GACTGCGTACCAATTCtat GACTGCGTACCAATTCact GACTGCGTACCAATTCaca GACTGCGTACCAATTCagt GACTGCGTACCAATTCatc GACTGCGTACCAATTCatg GACTGCGTACCAATTCaga GACTGCGTACCAATTCgat GACTGCGTACCAATTCgta GACTGCGTACCAATTCtac GACTGCGTACCAATTCtag GACTGCGTACCAATTCtga GACTGCGTACCAATTCtgt GACTGCGTACCAATTCagc GACTGCGTACCAATTCgct |
Table 1: The pre- and selective primers used in this study.
Cloning and sequencing of drought stress induced TDFs: The selective amplification PCR products were analyzed on a 6% denaturing polyacrylamide gel, and were silver stained following the manufacturer’s instructions for sequencing kit Q4310 (Promega Corporation, USA). The interested TDFs were excised from the gel and eluted in 50 μl ddH2O overnight. The eluted DNA was amplified by using its corresponding selective-amplification primer pairs. The PCR products were cloned into the pGEM-T easy vector (Promega) and sequenced by Augct Company (Beijing, China) using an ABI377 automated DNA Sequencer (Perkin-Elmer corporation, MA, USA).
qRT-PCR validation of drought stress induced TDFS: The primer pairs for qRT-PCR were design basing on the sequences of TDFs. If the TDF’s length was less than 300bp, designed primer pair with its highest homologous gene, and one primer must locate in the TDF sequence. The RNA samples were converted to single-strand cDNA by using a polyT with reverse transcriptase, then PCR amplification with qRT-PCR primer pair and SYBR Premix Ex Taq™ kit (Takara, Dalian, China) using Rotor-Gene6000 machine (Corbett Robotics, Australia), the beta-actin gene as control, and all samples replicated three times. The relative expression quantification of mRNAs was calculated using by the 2ΔΔCT method, and the primer pairs for qRT-PCR were list in Table 2.
| TDF Code | Primers Code | Primer(5’-3’) | TDF Size | Amplification Size |
|---|---|---|---|---|
| Actin | ActinS | CCTTCGTCTGGACCTTGCTG | 180bp | |
| ActinF | CAAGGGCAACATATGCAAGC | |||
| MtatBgagt376 | SEC14S | ATGGTGTGGATAAAGAAGGGAGAC | 376bp | 161bp |
| SEC14F | AGCAATGGTACAAGCAGGAAATT | |||
| EatgEatg367 | Psb-S | GAAATAGGCACAAGGAAAGAGCA | 367bp | 159bp |
| Psb-F | TTGAAGTAGTTGAATAGGAGGATCG | |||
| MtatBgagt141 | Fas-S | ATCAGCAAGAGTTCTGGCAAG | 141bp | 150bp |
| Fas-F | GAGATTCCTCCTCCGGTTAAA | |||
| EctcMatc183 | EIF3-S | AGGAAAGGAGACAACAAAGAAACT | 183bp | 162bp |
| EIF3-F | CGGACAGGATTAACTGTAAGCATT | |||
| MtgaEcga189 | FBPase-S | GAATTGCAGCTCTAGTAGCGTCTC | 189bp | 186bp |
| FBPase-F | CATCATCTTCTTCTGAAGCCATGA | |||
| EcttMtat281 | WD40-S | GCTCTGATGGAAGTTGCGTTAT | 281bp | 194bp |
| WD40-F | AATAGTGCGTGAAATCTTCTCTCC | |||
| MtgaEgag310 | UN1-S | AAGTTCTGAGCGGGACAGTAAAG | 310bp | 198bp |
| UN1-F | CGTGCCATCATAGCTAGGATAGG | |||
| MtcgEccc215 | UN2-S | ACAATCGTTCGGACTTGGTAAA | 215bp | 169bp |
| UN2-F | GGCTTCGGGATCGAGGTATC |
Table 2: Primer pairs for quantitative RT-PCR.
Establishment of the modified cDNA-AFLP technique
According to the modified cDNA-AFLP technical route, the double-strand cDNAs that contained the recognized site of MmeI were obtained and digested with three restricted enzymes (MmeI, EcoRI and BstYI), then the digested double-strand cDNAs linked with their corresponding adapters for pre- and selective amplification. Totally, 483 TDFs were got by 63 selective primer pairs with their lengths varying from 50 bp to 1500 bp, mainly at 100 bp to 400 bp, and about seven TDFs per primer pair on average (Figure 2). Sixty-three TDFs were reused and sequenced, 45 TDFs of them were ME, and ME represented that TDF had M and E primer at its both ends, five of them were MM, nine of them were MB, and the rest four ones were EE and BE (Table S1). Most TDFs had M primer, suggested that the recognized site of MmeI was integrated to double-strand cDNA successfully.
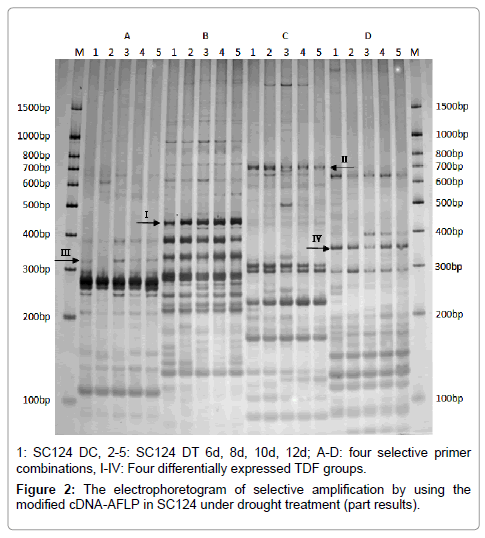
Figure 2: The electrophoretogram of selective amplification by using the modified cDNA-AFLP in SC124 under drought treatment (part results).
Many TDFs were induced by drought stress
The cDNA-AFLP gene differential expression display technique was considered as a semi-quantitative detection method, many TDFs showed differential expression under drought condition. Totally, 106 out of 483 TDFs showed differential expression, and could be divided into four groups according to their expression change trend during the drought treatment: 1) up-regulate, 32 TDFs; 2) down-regulate, 23 TDFs; 3) down-regulate after up-regulating, 44 TDFs; 4) up-regulate after down-regulating, 7 TDFs (Figure 2 and Table 3). In general, the group of down-regulate after up-regulating was the largest one, followed by up-regulate group and down-regulate group, and the group of upregulate after down-regulating was the smallest one. Furthermore, the number of down-regulated DE-TDFs (67) was more than that of upregulated ones (39) at the later period of drought stress.
| DE-TDF Groups | DE-TDF number | Percentage |
|---|---|---|
| Up-regulate | 32 | 30.2% |
| Down-regulate | 23 | 21.7% |
| Down-regulate after up-regulating | 44 | 41.5% |
| Up-regulate after down-regulating | 7 | 6.6% |
| Total | 106 | 100% |
Table 3: Four groups of differentially expressed TDFs (DE-TDFs).
The 5’end of DE-TDFs nearer to transcription start site
Sixty-three out of these 106 differentially expressed TDFs (DETDF) were reused, cloned and sequenced. After BLAST in http://www.phytozome.net/cassava, the positions of thirty-one DE-TDFs in their corresponding transcripts were determined in AM560 genome draft. The 5’end of four DE-TDFs began at transcription start site (TSS), two of them in 5’untranslated region (5’UTR), and the rest 16 ones located at coding sequence (CDS) regions, including two ones were less than 200bp, and 14 ones were more than 200bp far away from ATG. In addition, 35 DE-TDFs were got by the previous method with the same materials (Table S2), and among of them, the 5’end of only one DETDF began at TSS, other 34 ones all positioned in CDS region and over 90% of them were more than 200bp far away from ATG (Table 4). It suggested that there were much more 5’ends of DE-TDFs which began at TSS or nearer to TSS in the modified cDNA-AFLP technique.
| Position | Modified cDNA-AFLP | Previous cDNA-AFLPa | |||
|---|---|---|---|---|---|
| No. of DE-TDF | Percentage | No. of DE-TDF | Percentage | ||
| TSS/ATGb | 4 | 18.2% | 1 | 2.9% | |
| 5’UTR | 2 | 9.1% | 0 | 0 | |
| CDS | ≤200bpc | 2 | 9.1% | 1 | 2.9% |
| >200bp | 14 | 63.6% | 33 | 94.3% | |
| Total | 22 | 100% | 35 | 100% | |
Table 4: The 5’ends of DE-TDFs in their corresponding transcripts.
Homologies of many DE-TDFs involved in or responded to drought stress tolerance
The sequences of 67 DE-TDFs were annotated in http://www.phytozome.net/cassava and NCBI database, 32 of them were homologous with known functional genes, 20 of them were homologous with unknown genes, and the rest 15 ones which had no homologous gene, maybe novel genes that response to drought stress in cassava. These 32 annotated DE-TDFs involved in 11 biological processes, and majority of them were ascribed into five functional categories, including cell growth and apoptosis, signal transduction, transcription regulation, genetic information processing and peptide synthesis and metabolism (Table 5). Among of them, their homologies of several DE-TDFs involved in drought tolerance or responded to drought in previous reports. For example, Eukaryotic translation initiation factor 3 subunit (MactEctc183) imparted stress tolerance and could be a potential candidate gene for developing crop plants tolerant to abiotic stress in Arabidopsis [10]; Cellulose synthesis (MtgaEcga125) was important for drought and osmotic stress responses including drought induction of gene expression [11]; The enzyme activity of Ribulose-bisphosphate carboxylase (MtgaEcga58) were 10 to 30% lower in drought stress as compared to normal control in soybean [12]; Over-expression of a maize E3 ubiquitin ligase gene (MtagMtag277) enhanced drought tolerance through regulating stomatal aperture and antioxidant system in transgenic tobacco [13]; five maize Cystain family members (MtagMtag292) were down-regulated in response to water starvation [14]. Moreover, part of them responded to other abiotic stresses, such as LRR receptor-like Serine/threonine protein kinase (EcagMtat337), Phosphatidylinositol transfer protein (MtatBgagt376), Fructose-1,6-bisphosphatase (MtgaEcga189), etc [15-17].
| DE-TDFs | Size(bp) | Functional Description | Homologous Gene Number | Homology |
|---|---|---|---|---|
| Cell growth and apoptosis (6) | ||||
| MtagMtag292a | 292 | Ceramidases | cassava4.1_002110m XP_002520446.1 |
E=2e-48 ID=78/95(82%) |
| MtgaEgca176 | 176 | Prolyl 4-hydroxylase alpha subunit | cassava4.1_014646m DB948318.1 |
E=4.2E-52 ID=(117/119)98.3% |
| MtgaEcga93 | 93 | Leukemia Virus Rnase H Domain | 2HB5_A | E=5e-27 ID=69/69 (100%) |
| MtatEctc281 | 281 | WD domain, G-beta repeat | cassava4.1_000071m XP_002515073.1 |
E=5e-40 ID=70/73(96%) |
| MtagMtag277 | 277 | E3 ubiquitin ligase | cassava4.1_000529m XP_004307862.1 |
E=2e-56 ID=86/91(95%) |
| MtgaEcga92 | 92 | Ribosomal protein S10 | XP_003588337.1 | E=2.60E-06 ID=(37/42)88.1% |
| Signal transduction (5) | ||||
| EcagMtat337 | 337 | LRR receptor-like serine/threonine-protein kinase | cassava4.1_000537m XP_003618726.1 |
E=1e-34 ID=64/105(61%) |
| BgagcEctc346 | 163 | Nucleotide binding protein |
XP_002522890.1 |
E= 4e-28 ID= 52/54(96%) |
| MactEctc183 | 180 | Eukaryotic translation initiation factor 3 subunit | cassava4.1_000145m XP_002528386.1 |
E= 8e-19 ID= 42/52(81%) |
| EgtgMtga131 | 131 | Cystatin family member | cassava4.1_019939m AAF72202.1 |
E=1e-21 ID=42/42(100%) |
| MtgaEgcg439 | 439 | SH3 domain and tetratricopeptide repeats 1 | EDL37493.1 | E= 2.7 ID= 20/62(32%) |
| Transcription regulation (5) | ||||
| MtcgEccc215 | 215 | Mitochondrial ribosomal protein L9 | cassava4.1_016718m YP_005090474.1 |
E=7e-27 ID=68/71 (96%) |
| EcatMtag293 | 293 | Thiopurine S-methyltransferase | DB939009.1 | E=4.3 ID=20/49(41%) |
| MtcgBaagc141 | 141 | Fasciclin and related adhesion glycoproteins | cassava4.1_007317m XP_002309262.1 |
E=1e-15 ID=33/45(73%) |
| MtgaEgag281 | 281 | Protein coding | XP_002518871.1 | E=0.059 ID=21/56 (38%) |
| MtgaEgca213 | 213 | Protein coding | cassava4.1_000297m EMJ00881.1 |
E=1.7E-102 ID=(210/212)99.1% |
| Genetic information processing (3) | ||||
| BaagcMtcg132 | 132 | Retrotransposon protein | ABA94145.1 | E=3E-56 ID=128/131 (98%) |
| MtatBgagt376 | 376 | Phosphatidylinositol transfer protein SEC14 | cassava4.1_006270m XP_003603969.1 |
E=3E-69 ID=103/113(91%) |
| MtgaEgca115 | 115 | Splicing factor 3b, subunit 4 | cassava4.1_024763m DB937609.1 |
E=3.10E-36 ID=(84/84)100.0% |
| Peptide synthesis and metabolism (3) | ||||
| MtatEctt234 | 234 | Cell wall-associated hydrolase | XP_003638717.1 | E=2e-18 ID=41/41 (100%) |
| EccaMtga93 | 93 | Putative aminodeoxychorismatelyase | ZP_14386624.1 | E=2.00E-09 ID=27/28 (96%) |
| BaagcMtcg133 | 132 | Polyprotein | DB937177.1 | E=1E-54 ID=127/131 (97%) |
| Transcription factor (2) | ||||
| MactEctc330 | 331 | Transcription regulator AraC N-terminal arabinose-binding Domain | ZP_18864282.1 | E=2e-54 ID=99/109 (91%) |
| MtgaEagt266 | 266 | RNA-binding translational regulator IRP | cassava4.1_001348m XP_002530635.1 |
E=3e-94 ID=141/154(92%) |
| Energy metabolism (2) | ||||
| MtagEatc248 | 248 | ATP-dependent RNA helicase | cassava4.1_001726m XP_004306326.1 |
E=2e-45 ID=75/84(89%) |
| MtgaEcga58 | 58 | Ribulose-bisphosphate carboxylase | cassava4.1_017243m Q42915.1 |
E=9.7E-21 ID=(55/55)100.0% |
| Transport (2) | ||||
| EatcEatc527 | 527 | Sugar transporter | cassava4.1_034097m XP_002517103.1 |
E=7e-128 ID=199/208(96%) |
| EcgaMtga87 | 87 | Membrane-associated apoptosis protein | cassava4.1_012967m | E=1.9E-26 ID=(66/66)100.0% |
| Sugar metabolism (2) | ||||
| MtgaEcga189 | 189 | Fructose-1,6-bisphosphatase | cassava4.1_008978m XP_002532766.1 |
E=1e-21 ID=51/62(82%) |
| MtgaEcga125 | 125 | Cellulose synthase | cassava4.1_008148m | E=7.9E-53 ID=(117/118)99.2% |
| Photosynthesis (1) | ||||
| EatgEatg367 | 367 | Photosystem II P680 reaction center D2 protein |
cassava4.1_031110m YP_003330957.1 |
E=2e-28 ID= 66/68(97%) |
| Lipid metabolism (1) | ||||
| MtgaEgca99 | 99 | Triacylglycerol lipase | cassava4.1_004098m | E=3.4E-43 ID=(97/97)100.0% |
Table 5: Homology of 32 differentially expressed TDFs identified in modified cDNA-AFLP analysis.
Part of DE-TDFs were validated by qRT-PCR
Eight DE-TDFs were randomly selected to confirm their differential expression under drought treatment by using qRT-PCR, included WD domain (WD40), Eukaryotic translation initiation factor 3 (EIF3), Fasciclin (Fas), Phosphatidylinositol transfer protein (SEC14), Fructose-1,6-bisphosphatase (FBPase), Photosystem II P680 reaction center D2 protein (PsbD2), and two unknown DE-TDFs (UN1 and UN2). Five of them showed significantly differential expression (NCDT ≥ 1 or ≤ -1) in both KU50 and SC124 leaves under drought stress except for EIF3, FBPase and SEC14 (Figure 3). For example, Fas downregulated in both KU50 and SC124 consistently under drought stress; PsbD2 highly expressed in KU50 and SC124 leaves, but it’s up- or downregulation was opposite in these two varieties; WD40, UN1 and UN2 down-regulated in SC124, but down-regulated after up-regulating at DT6 in KU50. It suggested that majority of DE-TDFs really responded to drought stress and cDNA-AFLP technique was an effective tool to screen differentially expressed genes.
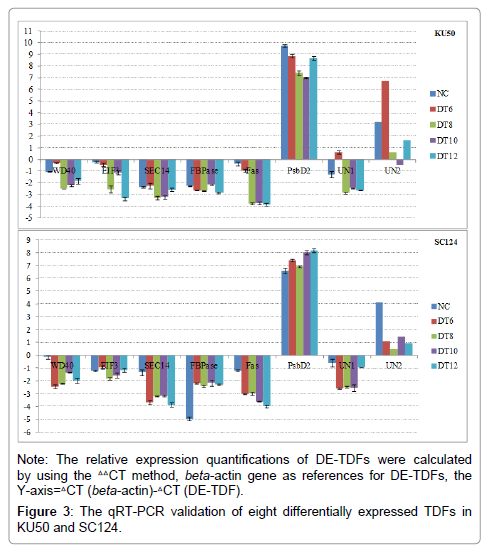
Figure 3: The qRT-PCR validation of eight differentially expressed TDFs in KU50 and SC124.
In this study, the modified cDNA-AFLP technique was established successfully, and over four hundred TDFs were got. The synthesis of single-strand and double-strand cDNA followed with the SMART technique in our modified technique. The MMLV reverse transcriptase that we used could add three C at the 3’end of single-strand cDNA when it arrived on the 5’ cap of mRNA, and this was the key point for our double-strand cDNA synthesis. The double-strand cDNA was synthesized with the modified 5’SMART IV™ Oligonucleotide primer and the simplified CDSIII/3’PCR primer, so a recognized site of MmeI was integrated to 5’end of the double-strand cDNA, and its cutting site was anchored at the transcription start site in theory.
In previous cDNA-AFLP technique, the low frequency of restriction enzyme EcoRI and the high frequency of restriction enzyme MseI were often used to digest the double-strand cDNA [18,19], and two high frequency of restriction enzymes-TaqI and MseI, were used for small genome-size microorganism occasionally [7]. In this study, because the cutting site of MmeI was anchored in the 5’end of double-strand cDNA, so another two enzymes were used to digest our double-strand cDNAs together. One was the frequently-used EcoRI, the other one was a novel enzyme BstYI, because the BstYI/MseI restriction enzyme combination could obtain more than 60% of all transcripts in tobacco by AFLP in Silico [8]. The tri-enzyme system resulted in 7.2 TDFs for each selective primer pairs on average, and the number of TDFs was less than that in the previous method. One cutting site was anchored at 5’end, the EcoRI and BstYI was low frequency enzyme, maybe three selective nucleotides should reduce to two ones for increasing the number of TDFs. Possibly, a high frequency enzyme with high GC content, e.g. CfoI (GCG?C), TaqI (T?CGA), etc, should be used to replace one of these two low frequency enzymes, because the GC content of CDS region was higher than that of other regions [20].
Although the 5’ends of DE-TDFs were nearer to the TSS than that in previous method, but only three 5’end out of 31 DE-TDFs started at TSS, and MmeI had recognized site in more than 10% of all transcripts in cassava. The MMLV transcriptase could add three C at the 3’end of single-strand cDNA when it arrived on the 5’ cap of mRNA, but if transcription interrupting by incomplete or complex second-structural mRNA, it also could add three C at the 3’end of single-strand cDNA with lower activity. Therefore, it might be the reason that many DETDFs didn’t start at the TSS.
Many homologies of DE-TDFs involved in or responded to drought stress, and five of eight randomly selective DE-TDFs up- or down-regulated in two varieties SC124 and KU50 significantly and consistency, the other three ones showed differentially expressed in at least one variety under drought stress. Especially, two unknown DETDFs responded to drought stress, suggested that possibly many novel drought resistance related genes existed in cassava genome.
In conclusion, the modified cDNA-AFLP method was not very perfect at this stage, had many details need to be optimized. Highquality RNA, the usage of high frequency enzyme and the reduction of selective nucleotide, maybe can make the modified technique to have more practicability. At present, although more and more gene differential expression analysis relied on the next generation sequencing technique, while our modified method gave a new light to cDNA-AFLP technique, possibly could prolong its usage life in some low-cost research activities or minor crops.
This work was supported financially by the National Basic Research and Development Program (2010CB126600) and Natural Science Foundation of China (31000537).