Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2016) Volume 6, Issue 1
The use of experimental patho-systems is a widely used strategy for understanding the pathogenic processes developed by many infectious agents. Ustilago maydis is a Basidiomycota fungus that naturally infects only maize and teozintle. We have established that under axenic conditions, either haploid or diploid strains of U. maydis can infect Arabidopsis thaliana, and in this work we analyzed the different behaviour of U. maydis haploid, diploid, and merodiploid strains in the mating genes during Arabidopsis infection. We determined the severity of infection, cell death, and the production of phytohormones and reactive oxygen species (ROS), occurring in the invaded host. Our results indicate that whereas U. maydis diploid and merodiploid strains behave as biotrophic parasites, the behaviour of the haploid strain is necrotrophic. We suggest that this difference resides in the fact that only the former two strains, but not the haploid one, are able to synthesize the master regulator of the biotrophic pathogenic behaviour, the bE/bW heterodimer encoded by the b genes of the two mating partners.
<Keywords: Ustilago maydis, Arabidopsis thaliana, Experimental pathosystem, Biotrophic, Necrotrophic
The use of alternative hosts for pathogenic microorganisms, and experimental pathosystems, has been a common practice to understand the pathogenic mechanisms in animal and plant infectious diseases. Under this idea, rabbits, mice, and even plants and Drosophila melanogaster have been used as experimental hosts for understanding the pathogenic processesof fungi on humans [1-3]. In the field of phytopathology, model pathosystems have also been established using model plants as host [4,5].
Ustilago maydis is a Basidiomycota fungus with a complex life cycle. The natural hosts of the fungus are maize (Zea mays ) and teozintle (Zea mays ssp parviglumis or Z. mays ssp mexicana ), but our working group has described that under axenic experimental conditions, U. maydis can infect different alternative hosts [6], among them Arabidopsis thaliana [7]. The master regulator of the biotrophic pathogenic process in the natural hosts is a heterodimeric protein complex denominated bE/bW, constituted by proteins encoded by the corresponding b genes of the mating strains that make the pathogenic heterokaryon [8]. Based on previous studies and results obtained in the infection of A. thaliana we hypothesized that the U. maydis infectious process may be necrotrophic or biotrophic depending as to whether A. thaliana is infected with haploid or diploid strains respectively [9].
In the present study we proceeded to determine whether the hypothesis suggested above was or not correct. Accordingly we have analyzed the severity of the symptoms caused in A. thaliana by U. maydis strains differing in their ploidy status, among them, the production of reactive oxygen species (ROS), cell death in the host tissues, and the production of different plant hormones. Our results suggest that the difference in behaviour among the strains, biotrophic or necrotrophic, is probably due to their capacity or incapacity to synthesize the bE/bW heterodimer.
Fungal strains and plant species
The U. maydis strains used in this study were the haploid FB2 (a2b2) [10], the merodiploid SG200 (a1mfa2bW2bE1) [11] and the diploid Uid1 (a1b1pan/a2b2Δodc::HygR) [12]. The plant host used was Arabidopsis thaliana L. Landsberg erecta .
Conditions for plant growth
A. thaliana plants were grown on solid MS synthetic medium [13]. The seeds were sterilized and germinated according to Martínez-Soto et al. [9].
Fungal growth and plant inoculation
U. maydis strains were maintained in 50% glycerol at 70°C, and recovered in liquid complete medium (MC; [14]) with carbenicillin (50 μg/ml) at 28°C, and shaken (200 rpm) for 24 h. Inocula preparations and plant inoculation were made according to Martinez-Soto et al. [9].
Damage and symptoms in infected plants
Symptoms of infection were determined according to Martínez-Soto et al. [9]. Tissue damage was observed with a stereoscope (Leica MZ-8), and photographed with a Spot digital camera (Diagnostic instruments). Plant biomass was determined by measurement of their dry weight.
Determination of reactive oxygen species (ROS)
Samples were stained according to Sato et al. [15] with some modifications. Tissue sections obtained with a scalpel were placed in an Eppendorf tube, and 500 μl of 1,2,3-dihydrorhodamine [1,2,3-DHR; Sigma-Aldrich (2.5 mg/ml in ethanol)] were added. Samples were incubated in darkness for 5 min, and observed with an epi-fluorescent Leica DMRE microscope, and photomicrographed as above. The presence of bright yellow fluorescence in the plant tissues indicated ROS production.
Determination of cell death
Cell death was determined according to Martinez-Pacheco and Ruiz-Herrera [16]. Tissue sections obtained as above were placed on a slide, and 15 μl of berberine sulfate (Sigma-Aldrich; 10 μg/ml), were added. The samples were incubated in darkness for 5 min. For microscopic observation by epi-fluorescence and microphotography we proceeded as above. Dead plant cells showed bright yellow fluorescence.
Determination of phytohormones produced by A. thaliana
Salicylic acid (SA): Salicylic acid was determined according to Malamy et al. [17]. Samples of 250 mg of wet plant tissue were frozen and macerated in a mortar with liquid N2. Next, 0.75 ml of 90% methanol and 5 μl of a solution containing 0.1 μg/μl of ortho-anisic acid (orthomethoxybenzoic acid) as internal standard were added. Samples were incubated for 24 h at 4 °C, centrifuged at 13,000 g for 15 min, the supernatant was saved, and the pellet was suspended in 0.75 ml of methanol, and centrifuged again. The supernatant was recovered and mixed with the previously obtained one. Methanol was evaporated with a N2 stream, and the pellet was suspended in 0.5 ml of 5% trichloroacetic acid (TCA). The sample was centrifuged at 6000 g for 10 min, the supernatant was recovered and 2 volumes of a mixture of ethyl acetate and cyclopentane or ethyl acetate-hexane (1:1) were added. The samples were incubated at room temperature for 10 min, the organic phase was recovered and dried again with N2 gas. Derivatization was performed by addition of 20 μl of pyridine and 80 μl of MSTFA [N-Methyl-N-(trimethylsilyl) trifluoroacetamide (Sigma- Aldrich, 394866)], and incubated at 80°C for 1 h. Finally, the problem samples, and samples of a standard curve were injected into a gas chromatograph (Agilent Technologies 7890A GC System, Palo Alto, CA) with a column DB-1 MS IU (60 m X 60.26 X 60.5 μm) coupled to a MSD 5973 detector.
Jasmonic acid (JA)
Jasmonic acid was determined according to Pluskota et al. [18]. Tissue wet samples of 500 mg were frozen and macerated with liquid N2 as above, and 1 ml of ethyl acetate, and 10 μl of dihydroxy jasmonic acid (DHJA; 0.1 mg/ml) as internal standard, were added. The sample was shaken, placed at 4°C for 24 h, and centrifuged at 13,000 g for 15 min at 4°C. The supernatant was recovered, and 500 μl of ethyl acetate were added to the pellet, shaken, and centrifuged again at 13,000 g for 15 min at 4°C. The supernatant was recovered and mixed with the previously obtained one. The solvent was evaporated with N2 gas and the derivatization process began. For derivatization process, 100 μl of N´N´diisopropylethylamine, 100 μl of chloroform, and 10 μl of pentafluorobenzyl bromide (PFB-Br) were added; the samples were incubated at 60°C for 30 min, the solvent was evaporated witha stream of N2 gas, and the sample was suspended in 100 μl of methanol HPLC grade. Finally, the samples and standard solutions were injected into a DB-1MS UI (60 m X 60.26 X 60.5 μm) column in the gas chromatograph (Agilent Technologies 7890A GC System; Palo Alto, AC) coupled to MSD 5973 detector.
Ethylene (ET)
A closed and sterile system was designed for measurement the ethylene production by plants of A. thaliana grown on MS synthetic medium. Petri dishes were sealed with egapack and parafilm, and a rubber stopper adapted to the upper lid to remove gaseous samples (10 ml) with the help of a sterile syringe and needle.
The samples were stored in sterile Vacutainer tubes, and subsequently were injected into a gas chromatograph (Hewlett 5890 Packard Series II; Ramsey, MN), with an Agilent Technologies HPPLOT Q (30 m × 0.530 mm × 40 μm) column. A standard curve with ethylene gas was prepared and used for data analysis.
Symptoms in infected Arabidopsis plants
As it was reported [9], under axenic conditions Arabidopsis plants infected with the haploid strain suffered more severe damage of infection than those infected with the diploid strain. Interestingly, plants infected with the merodiploid strain SG200 showed symptoms of infection similar to those infected with the diploid strain. Among the main symptoms we may cite: decreased size and biomass, growth of fungal mycelium mainly at the inoculation site, and necrosis (Figure 1).
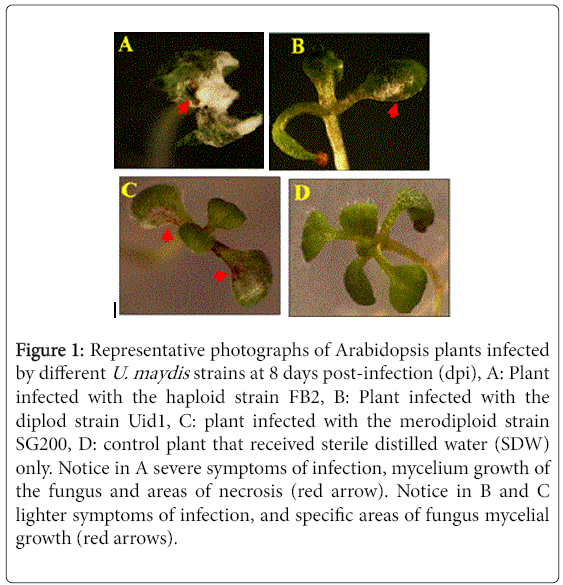
Figure 1: Representative photographs of Arabidopsis plants infected by different U. maydis strains at 8 days post-infection (dpi), A: Plant infected with the haploid strain FB2, B: Plant infected with the diplod strain Uid1, C: plant infected with the merodiploid strain SG200, D: control plant that received sterile distilled water (SDW) only. Notice in A severe symptoms of infection, mycelium growth of the fungus and areas of necrosis (red arrow). Notice in B and C lighter symptoms of infection, and specific areas of fungus mycelial growth (red arrows).
Growth of infected A. thaliana plants
Infected plants showed a reduced growth as compared to noninoculated plants. Nevertheless, it was observed that the biomass of Arabidopsis plants infected with the U. maydis haploid strain was much lower than plants infected with the SG200 merodiploid or Uid1 diploid strains respectively (Figure 2).
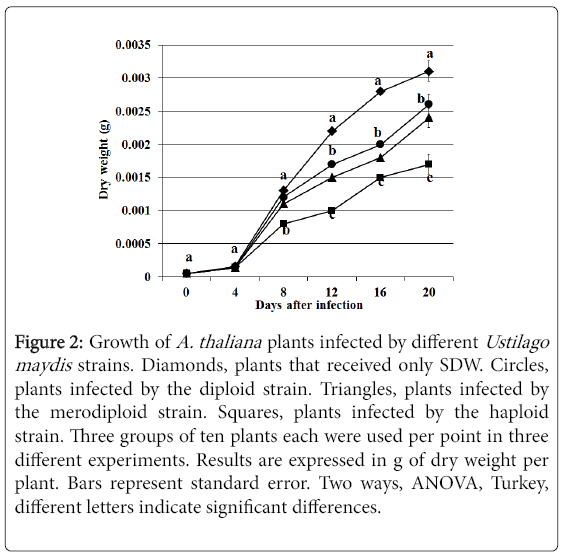
Figure 2: Growth of A. thaliana plants infected by different Ustilago maydis strains. Diamonds, plants that received only SDW. Circles, plants infected by the diploid strain. Triangles, plants infected by the merodiploid strain. Squares, plants infected by the haploid strain. Three groups of ten plants each were used per point in three different experiments. Results are expressed in g of dry weight per plant. Bars represent standard error. Two ways, ANOVA, Turkey, different letters indicate significant differences.
ROS production in infected Arabidopsis plants
In Arabidopsis plants infected by the haploid strain, the production of ROS was observed in practically in all the infected tissue.
On the other hand, in those plants infected with the diploid strain, ROS production was observed only in some areas; and in plants infected with the merodiploid strain, only some small points of ROS production were observed (Figure 3). As expected, no ROS was observed in control plants that received DSW only.
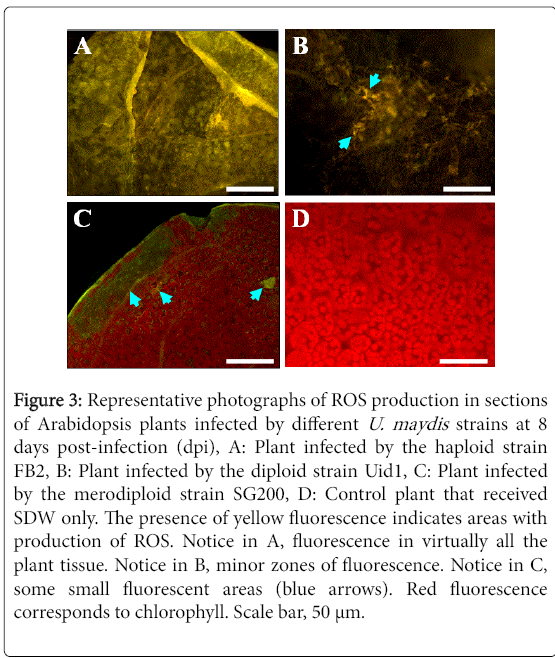
Figure 3: Representative photographs of ROS production in sections of Arabidopsis plants infected by different U. maydis strains at 8 days post-infection (dpi), A: Plant infected by the haploid strain FB2, B: Plant infected by the diploid strain Uid1, C: Plant infected by the merodiploid strain SG200, D: Control plant that received SDW only. The presence of yellow fluorescence indicates areas with production of ROS. Notice in A, fluorescence in virtually all the plant tissue. Notice in B, minor zones of fluorescence. Notice in C, some small fluorescent areas (blue arrows). Red fluorescence corresponds to chlorophyll. Scale bar, 50 μm.
Occurrence of cell death in Arabidopsis plants
Large areas of cell death were observed in Arabidopsis plants infected with the haploid strain, whereas in contrast those infected by the diploid or merodiploid strains, only some small points of cell death were observed (Figure 4). In control plants that received DSW only, no cell death occurred during the whole incubation period.
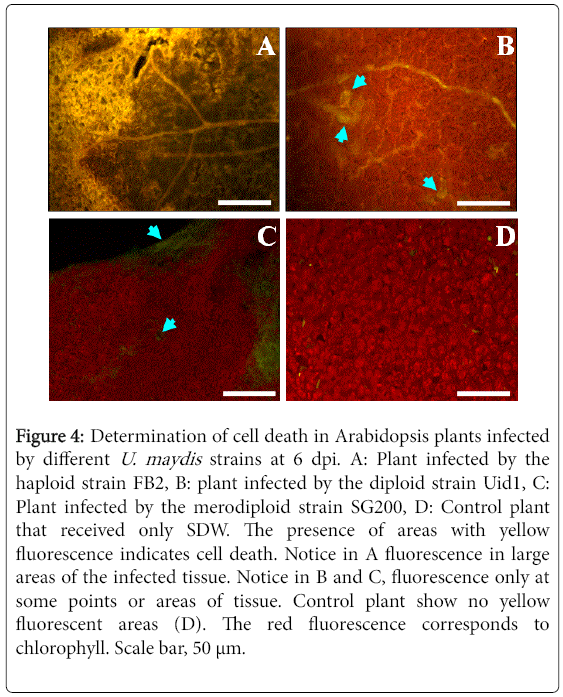
Figure 4: Determination of cell death in Arabidopsis plants infected by different U. maydis strains at 6 dpi. A: Plant infected by the haploid strain FB2, B: plant infected by the diploid strain Uid1, C: Plant infected by the merodiploid strain SG200, D: Control plant that received only SDW. The presence of areas with yellow fluorescence indicates cell death. Notice in A fluorescence in large areas of the infected tissue. Notice in B and C, fluorescence only at some points or areas of tissue. Control plant show no yellow fluorescent areas (D). The red fluorescence corresponds to chlorophyll. Scale bar, 50 μm.
Determination of phytohormones production in Arabidopsis plants
Infected Arabidopsis plants with the different strains of U. maydis produced increased levels of phytohormones, mainly jasmonic acid (JA), and salicylic acid (SA), compared to control plants that received SDW only.
Nevertheless the production of JA and ethylene was higher in plants infected with the FB2 haploid strains, whereas SA levels were higher in plants infected with the diploid or merodiploid strains (Figure 5).
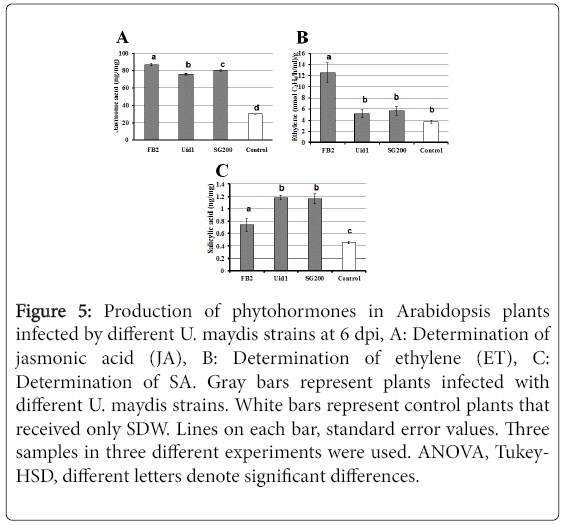
Figure 5: Production of phytohormones in Arabidopsis plants infected by different U. maydis strains at 6 dpi, A: Determination of jasmonic acid (JA), B: Determination of ethylene (ET), C: Determination of SA. Gray bars represent plants infected with different U. maydis strains. White bars represent control plants that received only SDW. Lines on each bar, standard error values. Three samples in three different experiments were used. ANOVA, Tukey- HSD, different letters denote significant differences.
The results obtained by infection of A. thaliana plants with the several strains of U. maydis showed interesting differences. For example, it was observed that growth of plants infected by the haploid strain showed a reduced growth compared with that of plants infected by the diploid or merodiploid strains.
These data agreed with the results obtained by the determination of ROS production, and cell death in the infected plants. Accordingly, Arabidopsis plants infected with the haploid strain FB2 developed large areas of ROS production in almost all their tissues, compared to those plants infected either by the diploid or merodiploid strains, which showed only small spots, or in some cases no production of ROS. Also noticeable was the difference in the levels of cell death observed in the tissues of the plants infected with the haploid strain when compared to those infected by the diploid or merodiploid, where cell death was almost absent. In this regard it is important to recall that programmed cell death (hypersensitive response, HR), together with the oxidative burst are induced in plants as part of their defense mechanism against pathogens [19].
It is known that during the biotrophic infection of maize by U. maydis there occurs the production of the effector protein Pep1, which inhibits the oxidative burst in the plant, and therefore the early immune response [20]. This phenomenon, as well as others involved in the biotrophic behaviour of the heterokaryon or diploid forms of the fungus, are under the control of a master regulator constituted by two products of the b mating loci of the fungus: a protein heterodimer made by a bW protein coming from one of the mating partners, and a bE protein coming from the other one (the bE/bW heterodimer) [8]. Accordingly, the merodiploid and the diploid strains used in this study are able to form the corresponding heterodimer, and it would be expected their observed biotrophic behaviour during the infection of A. thaliana . On the other hand, this does not occur in the Arabidopsis plants infected by the haploid strain that obviously is not able to synthesize the heterodimer, suggesting that this is the reason behind the necrotrophic parasite behaviour of this strain that causes severe damage in the plant, and the induction of an uncontrolled oxidative burst. In agreement with this hypothesis, previously we have demonstrated the over-expression of the PEP1 gene in the diploid strain, and its repression in the haploid strain of U. maydis during the Arabidopsis infective process [9].
An additional result that supports the present hypothesis is the observation that A. thaliana plants infected by the haploid strain produce a greater amount of jasmonic acid (JA) and ethylene (ET), and a lower production of salicylic acid (SA) in comparison with the plants infected with the diploid or merodiploid strains, a characteristic of the plants defense mechanism against necrotrophic fungi [19]. On the contrary, A. thaliana plants infected by the diploid or merodiploid strains produced higher amounts of salicylic acids (SA), than the haploid strain. These results are similar to the defense mechanisms developed by plants against necrotrophic or biotrophic fungi [19].
One interesting aspects of the pathogenic behaviour of U. maydis is its ability to reprogram transcriptionally its natural host, the maize, inducing changes in the primary and secondary metabolism of the plant during the infection process; including changes in the defense responses and production of phytohormones [21,22]. It has been demonstrated, that this interesting ability of the fungus depends on several effector proteins and the virulence factor Cmu1 (revised by Okmen and Doehlemann [23]), a chorismate mutase secreted by U. maydis , which is taken up by the plant cells causing metabolic changes [24]. Accordingly, this host manipulation is part of its biotrophic strategy [21]. We have demonstrated that the gene CMU1, several effector genes, and genes encoding degrading proteins of plant compounds are epigenetically regulated by the histone acetyltransferase Hat Gcn5 [25], and that GCN5 disruption eliminates the virulence of U. maydis in maize [26].
Recently, it was reported that in maize cobs infected with U. maydis , the levels of phytohormones, cytokinin (CKs) and abscisic acid (ABA) are induced (Morrison et al.). Both hormones can induce the synthesis of ET [27]. In agreement with that study, we observed that ET production is induced in A. thaliana plants infected by U. maydis mainly by the haploid strain. Moreover, H2O2 may induce defense mechanisms and production of phytohormones [19,21]. However, the diploid and merodiploid U. maydis strains can respond to early defense mechanisms of the plant by inducing genes such as YAP1, which in turn regulates several genes encoding peroxidases and which together form a system of detoxification against H2O2 [28]. These results agree with our observations that only diploid and merodiploid strains have the ability to avoid ROS production in the infected plant, something that does not occur in plants infected by the haploid strain.
In summary, the data presented here, confirm that in contrast to diploid and merodiploid strains, the haploid U. maydis strain behaves as a necrotrophic parasite in A. thaliana plants, probably because it is unable to synthesize the bE/bW heterodimer. These data are further evidence that the biotrophic capacity of U. maydis when infecting maize is under the control of this heterodimer. In addition, our data demonstrate the extreme plasticity of U. maydis to change its virulent capacity depending on the host, and its ploidy state.
This work was partially supported by Consejo Nacional de Ciencia y Tecnología (CONACYT), Mexico. Thanks are given to Jorge Molina- Torres and Juan J. Peña-Cabriales; J. Antonio Vera-Nuñez, Enrrique Ramírez-Chávez, John M. Haro-Velez, Ma. del Rosario Razo-Belmán, Claudia G. León-Ramírez and Lucila Ortiz-Castellanos for assistance in some analyses.