Journal of Depression and Anxiety
Open Access
ISSN: 2167-1044
ISSN: 2167-1044
Research Article - (2018) Volume 7, Issue 4
Background: Argan Oil (AO) has been used as a natural remedy in traditional medicine, mainly in Morocco, for several centuries. In this study, we evaluated the beneficial effects of AO dietary on vulnerability of rats to the chronic unpredictable mild stress (UCMS) using behavioral tests, biochemical and histological markers of depression or anxiety.
Method: Rats were handled daily (home cage control) or subjected to the UCMS procedure during 6 weeks (i.e., from 43th to 85 Post-natal Day (PND)) (Stress group, n=12). Animals were previously administered orally by NaCl 0.9% (Control group, n=11) or AO (10 ml/kg/day) (AO+Stress group, n=12) for 10 weeks (i.e., from weaning 21th to 93 PND). The efficacy of UCMS or AO dietary on behavioral performances of the animals in the open field, the forced swimming, the light/dark, the novelty suppression of feeding and sucrose preference tests, was measured. Following behavioral assays, oxidative stress in amygdala, histologic semiquantitative analysis of neurodegeneration in the hippocampus, frontal cortex and basolateral amygdala (BLA) subregions, and corticosterone level in plasma was also performed.
Results: Our data supports pharmacological and biochemical evidences for the antidepressant and anxiolytic-like effects of AO. Prolonged supplementation with AO reverses all the behavioral changes that occurred due to UCMS and restored corticosterone level in the plasma, oxidative status of amygdala and the neurons level in the CA3 subregion of rats’ hippocampus.
Conclusion: This study suggests that antidepressant and anxiolytic like effects of AO in adult rats can be the result of modulation of brain antioxidant enzyme activities, the activation of hippocampal neurogenesis and the modulation of HPA axis activity. However, more experiment and detailed analysis is required for definitive conclusion.
Keywords: Argan oil; Chronic mild stress; Anxiety; Depression; Neuroprotection; Amygdala; Wistar rats
AO: Argan Oil; BLA: Basolateral Amygdala Nucleus; CAT: Catalase; EPM: Elevated Plus Maze; Fr: Frontal Cortex; FST: Forced Swimming Test; GSH: Glutathione; GSH-Px: Glutathione Peroxidase; LDT: Light/Dark Test; LPO: Lipid Peroxidation; MDA: Malondialdehyde; MDD: Major Depressive Disorder; NBT: Nitroblue Tetrazolium; NO: Oxide Nitrite; NSF: Novelty Suppressed Feeding; OFT: Open Field Test; PUFAs: Polyunsaturated Fatty Acids; ROS: Reactive Oxygen Species; SOD: Superoxide Dismutase; SPT: Sucrose Preference Test; UCMS: Unpredictable Chronic Mild Stress
Argan oil (AO) is obtained from the argan fruit of Argania spinosa L., an endemic tree located mainly in southwestern Morocco [1]. Up to now, AO has been used as a natural remedy in traditional medicine, mainly in Morocco, for several centuries [2], mostly for the treatment of skin infections and in cosmetics [3,4]. Virgin AO is characterized by high levels of linoleic and oleic acids, tocopherols [5,6] and melatonin [7,8], which exhibit significant antioxidant activities. This compound makes AO a very interesting with respect to their actions on the risk factors of numerous diseases, mainly cardiovascular diseases, associated with hyperlipidemia, hypercholesterolemia, and hypertension [9-11]. AO composition has also been correlated to cancer chemoprotective effects especially due to the abundance of tocopherol [12].
For mental health, recent evidence in animal models developed in our laboratory (LGNB) demonstrates that AO may exhibit neuroprotection.
• In the adolescent bing-like ethanol model, 20 weeks of AO pretreatment improves the cognitive abilities in rats and attenuated oxidative stress in both prefrontal cortex and hippocampus [13].
• In the pilocarpine model used to induce epilepticus in wistar rats, AO administered by oral gavage increased catalase activity and attenuated oxidative stress in rat hippocampus [14].
• In the separation maternal model used to induce depression in rats, 10 ml/Kg/day of AO dietary for 13 weeks, reduced sensitivity to frightening environments with sex dependent variation [15].
• In the isolate social model used to develop schizophrenia in rats, AO supplementation reduced negative symptoms and working memory deficits that characterize schizotypal personality [Unpublished data].
In human emotional disorders, etiologic factors are numerous and genetic factors can interact with environmental factors to influence the vulnerability to major depression [16] or to anxiety disorders [15]. The accumulation of stressful life events has been reported to be a particularly efficient factor to precipitate the pathology. In view to a better understanding of the phenomena existing in stress related behaviors (anxiety and depression), different rodent models have been developed. Among them, the unpredictable chronic mild stress (UCMS) model has been extensively investigated in rats [17]. Rats submitted to chronic exposure to mild stressors show a variety of symptoms that characterize anxiety and depression disorders as revealed by general decrease in responsiveness to reward [18], anhedonia, depressed mood, and low energy [19,20].
Studies have shown that these behavioral deficits are correlated with structural changes in hippocampus, prefrontal cortex and amygdala, subregions highly sensitive to stress [21-23]. More, pathogenic associations among chronic stress, neuroplastic change, depression and anxiety are now established [24,25]. In the amygdala a key structure in managing emotions, stress-induced changes in morphological and functional plasticity including increased dendritic length and complexity [26-28]. These changes appear to be associated with altered protein expression [29] and oxidative stress [30]. Furthermore, exogenous stress promotes the release of glucocorticoid in vivo. In previous studies, chronic mild stress indirectly induces an exorbitant level of corticosterone, which can damage specific brain regions and lead to depression [31-33]. Moreover, all these behavioral and biochemical alterations induced by chronic mild stress can be reversed by chronic antidepressant administration [34-37], but always with side effects that alters the quality of life of the subjects.
To the best of our knowledge, no study has investigated simultaneously the effects of AO dietary on the behavioral response, biochemical and histological profiles of amygdala and hippocampus involved in emotional responses to stress. Accumulating evidence shows that consumption of AO with its unique profile of tocopherols, sterols, phenolic compounds and especially fatty acids should confer valuable mental health benefit. In particular, some compounds of AO may have preventive effects against stress since 1) their transport across the blood-brain barrier is easily done [38]; 2) they have shown a relevant anti-inflammatory effect [39] : increasing in inflammation markers is one of stress manifestations; also 3) lack of polyunsaturated fatty acids is of a etiological importance in stress-related affective disorder and there are evidences showing the antidepressant effects of omega-3 polyunsaturated fatty acids [40].
In respect of the above data, the main proposal of this work was to investigate in Wistar rats submitted to UCMS, the effect of virgin AO supplementation on:
• Anxiety – and depressive-like behaviors.
• Neurodegeneration in amygdala, hippocampus and frontal cortex sub-regions.
• Oxidative status of amygdala.
• Corticosterone level in plasma.
Animals
All experimental procedures were performed according to the NIH Guide for the Care and Use of Laboratory Animals. The animals used in this study are rats from the Wistar strain. They were born, bred, and housed on a 12 h light/12 h dark cycle, 50–60% humidity and at a standard temperature 21°C ± 1°C. Water and food were provided at labium. The standard pellet diet (ALF SAHEL Society, El-Jadida, Morocco) was a balanced diet containing protein 20.1%, fat 4.1%, carbohydrates 60.0%, fiber 5.8%, minerals 8%, and vitamins 2.0%.
The cages were cleaned, and food and water supplied three times per week during the course of the experiment. This work has been fully realized in Genetic, Neuroendocrinology and Biotechnology Laboratory (LGNB) located at Ibn Tofail University (Kénitra, Morocco). The study was carried out on 35 male and female rats (34 ± 2 g, 21 days old) that were divided into three groups:
• Control group: untreated (n=11)
• Stress group: submitted to UCMS (n=12)
• AO+Stress group: supplemented by AO & submitted to UCMS (n=12)
The experimental procedures are summarized in Figure 1.

Figure 1: Experimental design. Rats were handled daily (home cage control) or subjected to the chronic unpredictable mild stress (UCMS) procedure during 6 weeks (i.e., from 43th to 85 Postnatal Day (PND)) (Stress group, n=12). Animals were previously administered orally by NaCl 0,9% (Control group, n=11) or Argan Oil (AO) (10 ml/kg) (AO+Stress group, n=12) for 10 weeks (i.e., from weaning 21th to 93 PND). The efficacy of UCMS or AO dietary on behavioral performances of the animals in the open field (OFT), the forced swimming (FST), the light/ dark (LDT), the Novelty Suppression of Feeding (NSF) and Sucrose Preference (SPT) tests, was measured. Following behavioral assays, oxidative stress (OS) in amygdala (Amy), Histologic Semiquantitative analysis (His) of neurodegeneration in the Hippocampus, frontal cortex and BLA subregions, and corticosterone level in plasma (CORT) was also performed.
AO: Oral gavage procedure
The AO used in this study had a composition similar to that used in human food (Table 1, Annex) [41], originating in southwestern Morocco (Agadir-Inzggan) and was extracted from fresh seeds by artisanal methods without any preliminary treatment. It was preserved at room temperature in a bottle made of brown glass.
| Fatty acid | % |
|---|---|
| C16 : 0 | 13.4 |
| C18 : 0 | 5.4 |
| C18 : 1 n-9 | 44.8 |
| C18 : 2 n-6 | 35.7 |
| C18 : 3 n-3 | 0.1 |
| Sterols | mg/100 g oil |
| Schottenol | 142 |
| Spinasterol | 115 |
| Stigmasta-8,22-dien-3 β-ol | 9 |
| Others | 29 |
| Tocopherols | mg/Kg oil |
| α | 35 |
| β | 122 |
| γ | 480 |
| Phenolic compounds | μg/Kg oil |
| Vanilic acid | 67 |
| Syrigic acid | 37 |
| Feurulic acid | 3147 |
| Tyrosol | 12 |
Table 1: (Annex) Composition of virgin Argan Oil [41]. Virgin argan oil is harvested from the fruit of the argan tree (Argania spinosa L. Skeels, 1911), which naturally grows in Morocco (Agadir-Inzggan). Virgin argan oil is obtained by artisanal methods and, consequently, is not altered during the extraction step. Virgin argan oil is characterized by high levels of linoleic and oleic acids, tocopherols and melatonin.
Rats were aged 21 days (at weaning) when AO supplementation began. They were supplemented daily by intra-gastric gavage (1 ml of AO/100 g of body weight, between 8:00 am and 10:00 am) [13-15,42,43] during 10 weeks (i.e., from 21th to 93th PND). Control animals were treated orally for the same duration with NaCl 0.9%. At the same time, for 2 months, body weights were measured weekly.
Stress regimen
A UMCS protocol adapted from works of Willner or Moreau was used for this study [44,45]. Rats (Stress and AO+Stress groups) were submitted to unpredictable stressing events one to three times per day, during 6 consecutive Weeks (i.e., from 43th to 85th PND). The stress regimen consisted in 1 hour of period of confinement in restriction tube (20 cm long, 8 cm diameter) with unexpected noises, 30 min of forced bath in 30°C water, 3 h paired housing in damps sawdust, 2 h cages tilts 45°, 1 h social stress by changing partners in the cage, 3 h sawdust removal, 1 h darkness in the morning, 1 h hard light, 30 min shouts of raptor, or 2 consecutive days of light/dark cycle inversions. The sequence and moment of stressing events were randomly organized to avoid habituation or foresight and maximize their stressing effects. For ethical reasons, all nociceptive stressors have been excluded as well as water and food deprivation. The period of behavioral tests was considered as a continuation of the UMCS.
Behavioral testing
Immediately at the end of stress period, rats were submitted to different tests allowing evaluation of anxiety- or depressive-like behaviors in the morning of 8 consecutive days (i.e., from 86th to 93th PND). All rats were tested each day using one test, which was recorded on videotape. Animals were allowed to adapt to the experimental room for at least 1 h before testing. AO supplementation was continued during the testing period.
Open Field Test (OFT): The OFT was performed as described previously [46]. In brief, apparatus consist of a square wooden arena (100 × 100 × 40 cm) with a white surface covering the inside walls. The floor of the wooden arena was divided equally into 25 squares marked by black lines and defined as 9 central and 16 peripheral squares. The room was kept in a dim light with a video camera placed centrally above the box for recording the events. Each animal was placed in the center of the box and allowed to explore freely for 10 min. The quantified parameters were the time spent in the center of the area and the number of returns to the nine square central sections. Central perimeter residence time is used as measure of anxiety [47]. The number of returns to the central area is also an indicator of the emotional reactivity [48]. The central area of a novel environment is anxiogenic and aversive and the behavioral inhibition appears therefore as an avoidance behavior towards the central zone of the open field [49]. This wooden apparatus was cleaned after every trial using 5% ethanol.
Elevated Plus Maze Test (EPM): The EPM is an ethological model of anxiety in rodents provoked by the novelty and repulsion as a result of elevation and illumination of the maze [50]. The EPM consists of a wooden plus-shaped platform elevated 70 cm above the floor. Two of the opposing arms (50 cm × 10 cm) are closed by 40 cm high side and end walls, having an open roof. In order to avoid fall, the other two arms (open arms) were surrounded by 0.5 cm high edge, the four arms had at their intersection a central platform (10 × 10 cm). A 100-W lamp was placed exactly over the central platform. At the beginning of the test, the rats were placed on the central area of the maze facing an open arm. The following parameters of anxiety-related behavior were measured during the 5 min testing period: 1) entries into open arms, 2) time spent in open arms, 3) and number of full entries into the arms. Decreased anxiety- like behavior is illustrated by a significant statistical increase of parameters in open arms (time and/or entries).
Forced Swim Test (FST): The method followed was described by Porsolt et al. [51]. To assess depressive-like responses, rats were individually placed in a cylinder (height 50 cm; diameter 30 cm) containing 27 cm water (22 ± 1°C) from which they could not escape. The rats were placed in the water for 6 min and the duration of immobility was measured. The latency to the first bout of immobility was also recorded starting immediately after placing the rats in the cylinders. A rat was judged immobile when it ceased all active behaviors (i.e., struggling, swimming and jumping) and remained passively floating or making minimal movements necessary to maintain the nostrils above water. High percent time floating is interpreted as an increased depressive-like response.
Light/Dark Test (LDT): This test allows evaluation of rat’s aversive reaction in highly illumination situation [52-54]. The apparatus consists of two PVC boxes (25 × 25 × 25 cm), one dark and the other highly illuminated (1.700 Lux), connected by a tunnel (7 × 7 × 10 cm). Animals are placed individually in the dark side. The time spent in the light side, the latency to enter into the light side and the numbers of dark-light transitions are measured for 6 min as an index of anxiety.
Novelty Suppression of Feeding test (NSF): Animals were tested one at a time using a modified version of the Britton and Thatcher-Britton paradigm as a measure of novelty-induced suppression of appetitive behavior [55-57]. Animals were food deprived for 24 hours before testing. The animals were then provided with food either in a novel environment. The novel environment was a 100 × 100 × 40 cm wooden arena (standard lighting, 350 lx) with food provided in a cylindrical wire mesh hopper located in the center of the novel arena. To begin the test, all animals were placed one at a time into the periphery of the arena at the same starting point. The test session lasted for 6 min, and during this period, the experimenter scored the latency (s) to begin feeding, the total amount of time spent feeding, and the latency to first “visit” the food (i.e., to move within a 5 cm radius of the food container). If an animal did not eat within the test period, a score of 360 seconds was assigned for latency measures.
Sucrose Preference Test (SPT): The present model was a modified version of the test previously described [58]. At the beginning of the test (89th PND) all the groups (n=11 – 12) were singly housed during 3 days in individual cages. Two bottles were available in each cage, one with 300 ml of 30% sucrose (w/v) and the other also with 300 ml of tap water. (1) Sucrose consumption (ml) 300 ml; (2) water consumption (ml) 300 ml; and (3) total liquid, sucrose consumption + water consumption was daily measured. Bottles were counterbalanced across the left and the right sides of the feeding compartment and alternated in each test day. At the end of 72th hour, the bottles were removed, the consumption was noted, and the animals were returned to their previous housing conditions. Sucrose preference (percent) was calculated as follows: Preference (%) = [sucrose solution intake (ml)/ total fluid intake (ml)] × 100.
Histological evaluation
Following behavioral assays, a set of animals (half strength for each group, n = 5 animals) were deeply anesthetized with Hydrate Choral 7% (0.5 ml/100 g I.P) and transcardially perfused with heparinized 0.9% saline solution followed by 4% paraformaldehyde in 0.2M phosphate buffer. Surgical manipulation was performed only after both the corneal and the paw withdraw reflexes were abolished. Brains were removed from the skull and post-fixed for 12 h in the same fixative and cryoprotected in sucrose-glycerol solutions over 24 h. The following day, 30 μm serial slices are made with vibratome (LEIKA VT1000 S, China) and maintained at 4°C in a glycerol solution. Every section was mounted on gelatin-coated slides and dried overnight. Staining was performed with Cresyl violet for histological examination as previously described [59,60].
The amygdala (in particular basolateral amygdala nucleus (BLA)), the hippocampus and the frontal cortex neurons were visualized under an optical microscope (OPTIKA, B-500TiFL, Italy) with a professional HDMI camera. To highlight any differences in number and size of morphological elements, slices were observed at several magnifications (4x, 10x, 20x and 40x).
Assessment of neurodegeneration
Assessment of neurodegeneration was performed in Nissl-stained coronal brain sections in the hippocampus, amygdala and frontal cortex. First, severity of neuronal damage was semi quantitatively assessed by a grading system as previously described [61,62]: score 0, no obvious damage; score 1, slightly abnormal appearance of the structure without clear evidence of visible neuronal loss; score 2, lesions involving 20%–50% of neurons; and score 3, lesions involving >50% of neurons. Scoring was performed in hippocampal subregions (CA1, CA3 and dentate gyrus (DG)), amygdala, and frontal cortex (Fr1=area 1, Fr2=area 2) in four sections per rat (−2.30, −3.30, –3. 80, and −4.30 mm for the Hippocampus and BLA; 3.20, 2.70, 1.70, 1.00 for the Fr1 and Fr2 relative to bregma according to Paxinos and Watson [63]). Averaged scores from these sections of both hemispheres in each rat were used for calculation of group data. Second, the amount of polymorph neurons in CA3, BLA and Fr1 and Fr2 were quantified as described earlier [64] using Image 1.45 software (USA). Only cells of neuronal morphology and a diameter larger than 8 μm were counted. Per rat, three sections (at −2.30, −3.30, and −4.30 mm for the CA3 and BLA; at 3.20, 2.70, 1.70 for the Fr1 and Fr2 relative to bregma) were analyzed, and numbers of neurons were averaged from these sections.
Biochemical analysis
For the other half of the rats for each group (n=5 or 6 animals), venous blood samples were collected (between 18:00 h & 19:00 h) in order to determine corticosterone levels in plasma. The remaining blood was put into a dry tube for serum extraction and analysis of corticosteronemia (ng/ml) was measured by chemiluminescence (CMIA) on automaton architect system Abbott.
Subsequently, the brains were immediately removed and cooled on dry ice. Then, the amygdala (Bregma -2.12 to -4.30) was separated from the rest of the encephalon and homogenized in ice-cold 20 mM Tris-HCl buffer (pH 7.4) for determinations of lipid peroxidation level (LPO), nitrite content (NO), catalase (CAT) and superoxide dismutase (SOD) activities.
NO assay: An aliquot of crude homogenate (10%) for the amygdala was centrifuged at 4°C (800xg for 10min), and supernatant was used to analyze nitrite levels as described elsewhere [65]. Briefly, samples were incubated at room temperature for 20 min with Griess reagent (0.1% N-(1-naphthyl) ethylene diamine dihydrochloride; 1% sulfanilamide in 5% phosphoric acid; 1:1). The absorbance was measured at 550 nm and compared to that of standard solutions of sodium nitrite.
LPO assay: Lipid peroxidation was evaluated by measuring the thiobarbituric-acid-reacting substances (TBARS) in homogenates, as previously described [66]. Briefly, an aliquot of crude homogenate of the amygdala was centrifuged at 4°C (1000xg for 10min), and supernatant was mixed with 1 ml 10% trichloroacetic acid and 1 ml 0.67% thiobarbituric acid. It was then heated in a boiling water bath for 15 min and butanol (2:1, v/v) was added to the solution. After centrifugation (800xg for 5 min), TBARS were determined from the absorbance at 535 nm. The results above were expressed as nmol of malondialdehyde (MDA)/g wet tissue.
CAT activity: Catalase activity was measured by the method that uses H2O2 to generate H2O and O2 [67]. In a 3 ml quartz tank, 1.95 ml of 50 mM phosphate buffer, 1 ml of H2O2 (0.019 M) and 0.05 ml of the sample were added [68]. The absorbance was measured at 240 nm at time 0 (T0) and after every 30 seconds for 2 min. Results are expressed as mmol/min/mg of protein.
SOD activity: The SOD activity was evaluated by measuring its ability to inhibit the photoreduction of NBT (nitro blue tetrazolium) [69]. In an aerobic medium, the riboflavin/methionine/NBT mixture gave a blue coloration, the optical density of which was measured at 580 nm. One unit of SOD corresponds to the amount of protein necessary to inhibit photoreduction by 50%.
Statistical analysis
Statistical analyses were performed using Prism 6.01 software (GraphPad Software). All data were expressed as mean ± SEM and have been statistically analyzed with one-way ANOVA followed by Tukey’s test or Student’s t-test. Nonparametric data resulting from semiquantitative analysis of histological staining were analyzed by Kruskal–Wallis ANOVA, followed by Dunn’s post hoc test comparing the control group with the Stress group (Stress effect) and Stress with AO+Stress groups (AO effect). p<0.05 were considered as statistically significant.
AO Supplementation reduces a body weight loss and relative Adrenal weight gain in rats exposed to UMCS over a period of 6 weeks. As illustrated in Figure 2A, the body weight of the animals was evaluated during the entire period (10 weeks) of supplementation with AO (i.e., from the 21th day until the 93th day of life) at intervals of 7 days. One-way ANOVA with repeated measures revealed significant effects for the treatment factor (F2, 96 = 8.76; p=0.0003) and for the repetition factor (F7, 96 = 39.82; p<0.0001). However, no difference was founded for the interaction factor between treatment and repetition (F14,96 = 0.96; p=0.4995).
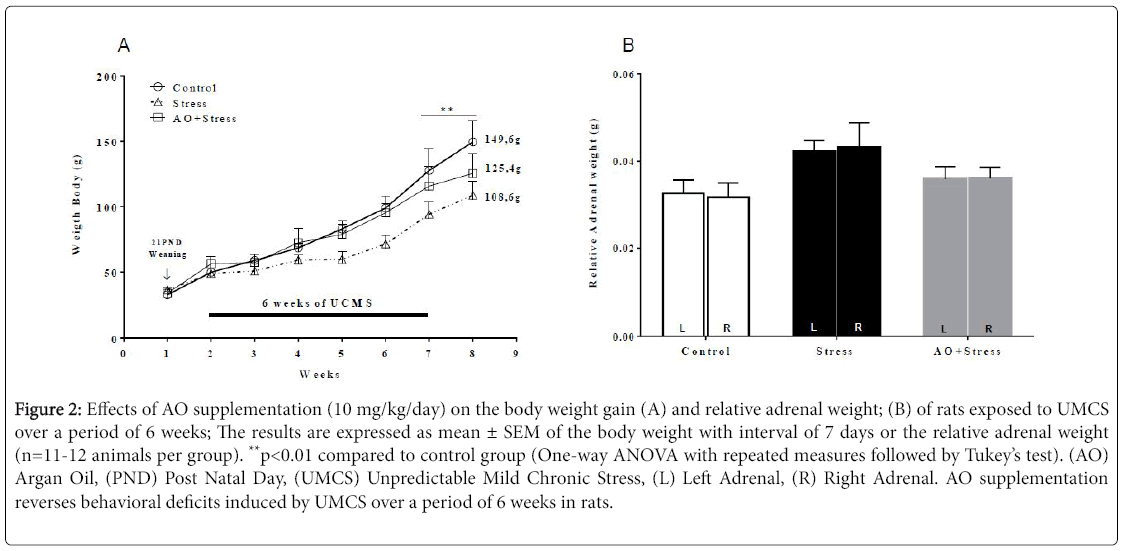
Figure 2: Effects of AO supplementation (10 mg/kg/day) on the body weight gain (A) and relative adrenal weight; (B) of rats exposed to UMCS over a period of 6 weeks; The results are expressed as mean ± SEM of the body weight with interval of 7 days or the relative adrenal weight (n=11-12 animals per group). **p < 0.01 compared to control group (One-way ANOVA with repeated measures followed by Tukey’s test). (AO) Argan Oil, (PND) Post Natal Day, (UMCS) Unpredictable Mild Chronic Stress, (L) Left Adrenal, (R) Right Adrenal. AO supplementation reverses behavioral deficits induced by UMCS over a period of 6 weeks in rats.
Subsequent post-hoc comparisons using the Tukey’s test revealed that UMCS induced a significant reduction in the body weight gain of rats specifically from the 1st week after the stress period (p<0.01). On contrary, no difference was observed between Stress and AO+Stress groups for this parameter, and this during all the period of the experiment. This result suggests an improvement in the physical state of animals under stress conditions by AO supplementation. In the other hand, the estimate of relative weight of the adrenal glands (Figure 2B) shows no significant difference between the three groups (ANOVA, p=0.0918).
All parameters evaluating the behavioral deficits stress related (anxiety and depression) of the animals were affected by the UMCS. Student’s t-test indicated a significant reduction (p<0.05) in the time spent in central area, in the open arms and in the light side respectively in the OFT, the EPM and the LDT. Also, the Sucrose preference (%) and the latency to feed after 24 h of food-deprivation in SPT and NSF respectively, was very significantly decreased in stressed rats, while the immobility time in the FST has been significantly increased by the UMCS. The locomotor activity of animals was also affected. Indeed, total number of entries in all arms and the total number of squares crossed in the EPM and the OFT respectively, were very significantly decreased in stressed rats compared to Control group.
On the contrary, in rats supplemented with AO (AO+Stress group), all these parameters were improved, especially those related to the depressive-like behaviors. Indeed, AO supplementation increases significantly the % of Sucrose preference and the latency to feed after 24 h of food-deprivation. On the other hand, AO supplementation increases the time of immobility of stressed rats in the FST. The only parameter in relation to anxiety that has been significantly improved by the AO supplementation is the time spent in open arms in the EPM test. This result suggests that AO pretreatment over period of 10 weeks induces an anti-depressive and anxiolytic like effect in animals exposed to 6 weeks of UCMS. Concerning the locomotor activity of animals, no significant changes were observed after the AO supplementation. Table 2 summarizes all results.
| Tests | Parameters | Control | Stress | AO+Stress | p(a) | p(b) |
|---|---|---|---|---|---|---|
| Control vs. Stress | Stress vs. AO+Stress | |||||
| OFT | Total number of area | 147 ± 13.54 | 90 ± 6.01 | 110 ± 6.86 | p=0.0048** | p=0.0535NS |
| Time spent in central area (s) | 34 ± 4.79 | 15 ± 2.20 | 17 ± 2.81 | p=0.0070** | p=0.5609NS | |
| EPM | Total number of entries in all arms | 26 ± 2.04 | 15 ± 1.28 | 17 ± 2.07 | p=0.0011** | p=0.4987NS |
| Time spent in open arms (s) | 123 ± 5.03 | 98 ± 3.09 | 110 ± 4.17 | p=0.0020** | p=0.0373* | |
| SPT | Sucrose preference (%) | 78 ± 5.57 | 45 ± 4.95 | 71 ± 5.07 | p=0.0008*** | p=0.0030** |
| FST | Time of immobility (s) | 20 ± 1.60 | 39 ± 2.88 | 28 ± 2.59 | p=0.0003*** | p=0.0162* |
| LDT | Time spent in Light box (s) | 220 ± 6.42 | 158 ± 12.7 | 191 ± 7.06 | p=0.0052** | p=0.0631NS |
| NSF | Latency to feed (s) | 151 ± 6.97 | 590 ± 12.17 | 233 ± 10.96 | p<0.0001*** | p<0.0001*** |
Table 2: Effects of AO supplementation (10 mg/kg/day) on performances of rats exposed to UMCS during adolescence over a period of 6 weeks in behavioral tests. The results are expressed as mean ± SEM of Times measured in seconds. p: Probability; (a) Stress effect, (b) AO effect, NS Nonsignificant; *Significant; **Very significant; ***Highly significant. Six rats per group were tested. Unpaired t test with Welch's correction. (OFT) Open Field Test; (EPM) Elevated Plus Maze test; (SPT) Sucrose Preference Test; (FST) Forced Swimming Tests; (LDT) Light/Dark Test; (NSF) Novelty Suppressed Feeding test. AO Supplementation reduces oxidative stress of amygdala and Corticosteronemia in rats exposed to UMCS over a period of 6 weeks.
In order to assess modification of biochemical parameter in rats supplemented for 10 weeks by AO, biochemical analyses were performed on amygdala homogenate. Table 3 shows that oxidative status in the amygdala of rats was significantly modified by the UMCS, as evidenced by a significant increase of the pro-oxidant markers and a decrease of the anti-oxidant markers. Indeed, Student’s t-test indicated a significant increase (p<0.05) in both NO and MDA levels in stressed rats. However, this same statistical test reveals a significant reduction (p<0.05) in both CAT and SOD enzymatic activities in rats exposed to UMCS.
| Parameters | Control | Stress | AO+Stress | p(a) | p(b) | |
|---|---|---|---|---|---|---|
| Control vs. Stress | Stress vs. AO+Stress | |||||
| Oxydative stress in Amygdala |
MDA (nmol/g of homogenate) |
5.46 ± 0.39 | 8.55 ± 0.59 | 6.39 ± 0.26 | p=0.0024** | p=0.0101* |
| NO (µmol/g of homogenate) |
87.90 ± 6.08 | 157.85 ± 4.58 | 130.52 ± 10.89 | p<0.0001*** | p=0.0650NS | |
| CAT activity (µmol/min/mg of protein) |
7.87 ± 0.54 | 5.60 ± 0.48 | 6.02 ± 0.14 | p=0.0144* | p=0.4552NS | |
| SOD activity (µmol/min/mg of protein) |
57.69 ± 3.05 | 34.99 ± 4.58 | 43.46 ± 2.83 | p=0.0045** | p=0.1623NS | |
| Corticosteronemia (ng/ml) (between 18:00 h & 19:00 h) |
117.94 ± 2.29 | 138.63 ± 5.47 | 125.52 ± 5.47 | p=0.0156* | p=0.1288NS | |
Table 3: Effects of AO supplementation (10 mg/kg/day) on oxidative status of amygdala and Corticosteronemia in rats exposed to UMCS over a period of 6 weeks. The results are expressed as mean ± SEM. p: Probability; (a) Stress effect, (b) AO effect, NS Non-significant; * Significant; **Very significant; ***Highly significant. Six rats per group were tested. Unpaired t test with Welch's correction. (MDA) Malondialdehyde; (NO) Nitrite content; (CAT) Catalase; (SOD) Superoxyde Dismutase.
Nevertheless, in supplemented rats the pro-oxidant/antioxidant balance in the amygdala tends to be regulated. Indeed, AO supplementation reduces the NO levels and increases the enzymatic activities of CAT and SOD, but the difference between Stress and AO +Stress groups is not significant. In contrast, the decrease in lipid peroxidation is statistically significant in UCMS rats supplemented with AO. Which might as interpret by a protective effect of AO against oxidative damage of the amygdala.
Additionally, corticosterone level in plasma was measured with the aim to evaluate emotions related to stress in supplemented rats. Our results show that 6 weeks of UMCS increases significantly a corticosteronemia in rats. However, this parameter has a tendency to increase in supplemented animals (Table 3).
AO Supplementation reduces a neurodegenerative effect of stress in brain of rats exposed to UMCS over a period of 6 weeks. Effects of AO supplementation (10 mg/kg/day) on Histologic Semiquantitative analysis of neurodegeneration in hippocampal subregions, amygdala, and cortical subregions in rats exposed to UMCS over a period of 6 weeks. (Figures 3A-3F).
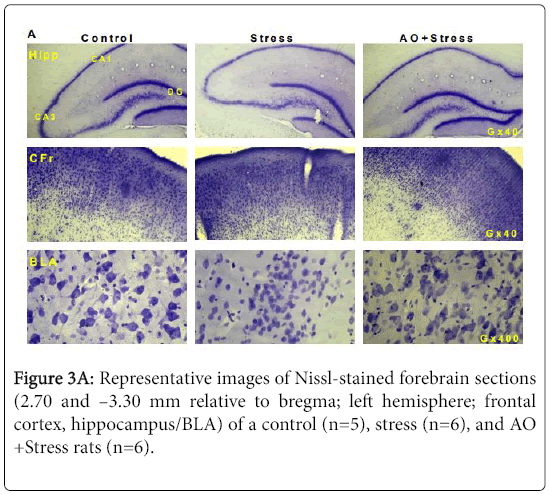
Figure 3a: Representative images of Nissl-stained forebrain sections (2.70 and –3.30 mm relative to bregma; left hemisphere; frontal cortex, hippocampus/BLA) of a control (n=5), stress (n=6), and AO +Stress rats (n=6).
Figure 3b: Regions of interest and their anatomical positions: Fr1 and Fr2: Frontal Cortex area 1 and 2; Hipp: Hippocampus; BLA: Basolateral Amygdala Nucleus.
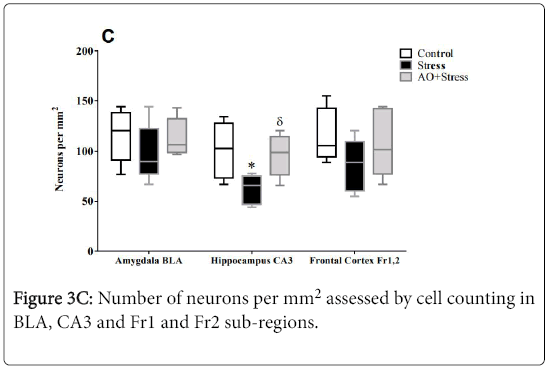
Figure 3c: Number of neurons per mm2 assessed by cell counting in BLA, CA3 and Fr1 and Fr2 sub-regions.
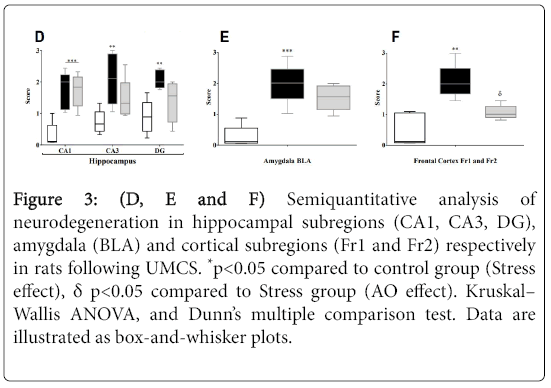
Figure 3: (D, E and F) Semiquantitative analysis of neurodegeneration in hippocampal subregions (CA1, CA3, DG), amygdala (BLA) and cortical subregions (Fr1 and Fr2) respectively in rats following UMCS. *p<0.05 compared to control group (Stress effect), δ p<0.05 compared to Stress group (AO effect). Kruskal– Wallis ANOVA, and Dunn’s multiple comparison test. Data are illustrated as box-and-whisker plots.
Semiquantitative analysis of neurodegeneration in hippocampal and cortical subregions as well as amygdala revealed neuronal loss (example shown in Figure 3A) already after 6 weeks of UCMS in the BLA (Control vs. Stress; p=0.0008; Figure 3E) and the frontal cortex (Control vs. Stress; p=0.0017; Figure 3F). Also, hippocampal pyramidal cells of CA1, CA3 and DG subregions reached statistical significance (p=0.0020; p=0.009; p=0.0023 respectively; Figure 3D). Moreover, the number of neurons per mm2 as assessed by cell counting was significantly reduced in CA3 subregion of hippocampus in stressed rats (p=0.0026; Figure 3C). Thus, in both of BLA and frontal cortex, no difference was found between groups in the number of neurons (Control vs. Stress; BLA, p=0.3260; Fr1 and Fr2, p=0.1086; Figure 3C).
Nevertheless, in supplemented rat’s neurodegeneration has a tendency to increase in both hippocampus and BLA (Stress vs. AO +Stress; CA1 p=0.9781; CA3 p=0.1983; DG p=0.0598; BLA p=0.2340; Figures 3D & 3E). Indeed, in the frontal cortex AO supplementation decreases significantly a neuronal loss in rats submitted to stress condition (Stress vs. AO+Stress; p=0.0067; Figure 3F). Additionally, the number of neurons tended to increase in both of BLA and frontal cortex in AO+Stress rats. Thus, in the CA3 subregion of hippocampus AO supplementation increases significantly this parameter (Stress vs. AO+Stress; p=0.0246). These data suggest a neuroprotective effect of AO against neurodegenerative processes induced by chronic stress in rats.
The present study demonstrates, for the first time, that AO supplementation over a period of 10 weeks induces a significant improvement in emotional behaviors severely impaired by chronic stress in rats. These behavioral changes were accompanied by a significant
• Reduction of oxidative stress in the amygdala.
• A regulation of cortico-steronemia.
• Neuroprotection against cellular alterations in the frontal cortex, hippocampus and BLA subregions of UCMS rats.
The diathesis-stress hypothesis of emotional disorders states that chronic stress could induce physiological alterations that promote an exaggerated stress response, which in turn would cause depressive or anxious events [70,71]. Thus, in this study, we evaluated the beneficial effects of AO dietary (10 ml/Kg/Day) on vulnerability of rats to UCMS using behavioral tests, biochemical and histological markers of depression or anxiety. In the following paragraphs we will discuss separately the antidepressant and the anxiolytic-like effects of AO in rats submitted to 6 weeks of UCMS (i.e., from the 43th day until the 85th day of life).
Antidepressant-like effects of AO supplementation
UCMS, one of the most reliable and valuable rat models of depression, are extensively used to screen for antidepressants and to investigate the pathophysiology of depression [72,73]. In the UCMS model, animals were subjected to different kinds of stress daily, which imitated unpredictable stressful life events and induced depressive-like behavior (e.g. anhedonia, despair state) that resemble those seen in Major depressive disorder (MDD) patients and could be reversed by chronic treatment of antidepressants [74]. Confirming previous findings [75,76], our UCMS rats showed decreased sucrose preference (%) in SPT, decreased locomotor activity in both of OFT and EPM test, increased immobility time in FST, and increased latency to feed in NFS test. Besides anhédonie and despair state, UCMS paradigm generates other outcomes related to depression such as body weight loss. This sign may be interpreted as a manifestation of a depressive-like state in rats since reduction of body weight is considered a symptom of the human disorder, mainly in comorbidity with anxiety disorders [77].
Notably, we demonstrated that UCMS-induced behavior changes could be reversed by prolonged administration of AO (10 mg/kg/day), which confirmed further the antidepressant effect of AO. These data are in line with a previous study conducted in our laboratory, evaluating the impact of AO dietary on the behavioral response to stress, using depressive and anxiety measurement paradigms [15]. Without using a stress induction procedure, the results of the study referred to, showed that AO supplementation (10 ml/Kg/day) during 13 weeks has resulted in an increase in locomotor activity and reduced sensitivity to frightening environments (FST, EPM and OFT). Additionally, no changes were found for the body weight gain in AO +Stress rats during the entire period of the experiment.
Restriction of body weight in rats exposed to chronic stressors is a physiologic correlate evoked by changes in a central level. Stress response is mediated by a main secretagogue, the 41-aminoacid peptide corticotrophin releasing factor (CRF), which is synthesized in neurons of hypothalamic or extrahypothalamic nuclei (i.e., the paraventricular nucleus and central amygdala, respectively) [78]. A huge number of studies demonstrate that the neuroendocrine response to acute and chronic stresses is mainly mediated by the hypothalamus pituitary adrenal (HPA) axis [79]. This axis plays an important role in the pathophysiology of stress-related diseases such as depression and anxiety [80]. Several lines of animal and human studies demonstrated that chronic stress is associated with hyperactivity of HPA axis, high plasma glucocorticoid levels and adrenal hyperplasia [81-83]. In agreement with this data, we found that 6 weeks of UCMS induces elevated levels of corticosteronemia in rats. AO supplementation did not change this setting. The relative weight of the adrenal was not equally affected either by stress or by AO supplementation.
Increasing evidence indicates that oxidative stress has been implicated in the etiology of depression [30]. Studies have demonstrated that reactive oxygen species (ROS) including superoxide anion, hydrogen peroxide and singlet oxygen induced neuronal damage has an important role in the pathophysiology of depression, probably via membrane omega 3 polyunsaturated fatty acids (PUFAs) pathology [84,85], decreasing activity of glutathione peroxidase (GSHPx), CAT and SOD [86], and antioxidant vitamin [87], suggesting oxidative damage. In agreement with a previous report [30], our results showed that UCMS induces an imbalance between oxidative and antioxidative stress makers in rats’ amygdala. Compared to control group, UMCS decreased significantly the CAT and SOD enzymatic activities and increased the NO and the MDA contents in amygdala.
Nevertheless, in supplemented/stressed rats the pro oxidant/ antioxidant balance in the amygdala tends to be regulated. We found in UCMS animals, that AO supplementation slightly reduced the NO level and increased the enzymatic activities of CAT and SOD. Interestingly, AO supplementation reduced significantly the LPO in the amygdala of UCMS rats. Results from our laboratory using the same dose of AO (1 ml/100 g weight), and the same duration of supplementation (10 weeks), demonstrated that AO pretreatment produces significant decrease of the LPO and NO levels and increases of the CAT activity in Wistar rat hippocampus after a acute phase of pilocarpine-induced epilepticus seizures [14].
The biological activities of AO are mainly attributed to its content in major antioxidant molecules, tocopherols (α- and γ-tocopherol) and polyphenols [41,88,89]. Recent evidence suggests that Coenzyme Q10 (CoQ10) and melatonin, also identified in AO, have antioxidant properties [88,89]. As tocopherols, polyphenols, CoQ10 and melatonin are able to prevent oxidative stress and mitochondrial and/or peroxisomal dysfunctions, which are considered major events in several neurodegenerative diseases [90,91], these biological properties could at least in part explain some of the neuroprotective effects of AO found in our study.
Thus, as AO, which contains numerous nutrients able to cross the blood-brain barrier (fatty acids, phytosterols, polyphenols, tocopherols, etc.), can prevent neurotoxicity in several animal models and stimulate the activity of several anti-oxidant enzymes in the brain, it was important to determine its impact at the cellular levels on nerve cells. By histologic examinations we found that 6 weeks of UCMS did not lead to changes number of neurons in frontal cortex (Fr1; Fr2) and BLA subregions. Contrariwise, this parameter was significantly affected in the CA3 subregion of hippocampus, suggesting a neurodegenerative effect of UCMS in rats. Moreover, semiquantitative analysis of neurodegeneration in hippocampal subregions (CA1, CA3, DG), amygdala (BLA) and frontal cortex (Fr1,2), revealed neuronal loss in UMCS rats compared to control group. Early studies on effects of stress on the brain demonstrated that animal models of repeated stress cause dendritic atrophy in CA3 pyramidal neurons of the hippocampus [92,93], and dendritic atrophy in pyramidal neurons of the medial frontal cortex [94]. Interestingly, our results show that Argan supplementation induces an improvement in all these structural changes, suggesting a neuroprotective effect of this oil against chronic stress-related neuronal damage.
Studies have demonstrated that AO administered by oral gavage was shown to have cytoprotective effects on the brain of Sprague Dawley rats treated with acrylamide to induce inflammatory and oxidative stress-related neurotoxicity. These protective effects were reported on mitochondrial function, the anti-oxidant system and the activities of NADPH-generating enzymes [95]. AO has also been reported to attenuate genetic damage and emperipolesis in rats treated with acrylamide [96]. In addition, in the model of neurodegeneration induced by aluminum chloride in male wistar rats (2.5 years old), AO given by oral gavage (6% w/w of AO in the food) for 42 days was also able to attenuate the decrease in catalase activity and to stimulate glutathione peroxidase activity in the hippocampus and cortex [97]. Therapeutic potentials of AO have been well demonstrated experimentally in neurological problems such as Alzheimer’s disease [98] and cognitive deficits [13]. Therefore, it seems that AO attenuated depression-like behaviors in stressed animals partially through its antioxidant, anti-inflammatory, and antiapoptotic properties.
Anxiolytic-like effect of AO dietary
Since anxiety is another psychiatric disease where the stressful events play an important etiologic role, we decided to evaluate the impact of AO supplementation on rodent’s anxious like behaviors. In this study, we evaluated anxious-like behaviors using three tests sensitive to previous stress exposure: OFT, EPM and LDT [99]. We found that UCMS decreased significantly the time spent in central area in the OFT, decreased the time spent in open arms in the EPM and decreased the time spent in light box in the LDT. These data together suggest that stressed animals have an increased preference for the closed and dark spaces which is considered an anxious-like behavior [100,101]. A huge number of previous studies [102,103] have similarly reported the anxiogenic effect of UCMS using the same behavioral tests in different strains of rodents (Sprague Dawley, Long Evans and Wistar rats).
In contrast with results of depressive-like behaviors, here we found that AO supplementation only increased the time spent in open arms in the EPM test. Thus, suggesting an antidepressant-like effect instead of anxiolytic-like effect of AO supplementation in UCMS rats. Without stress exposure procedure, Rim and al. showed that 13 weeks of AO supplementation affected significantly the number of recoveries, the number of returns to central area in OFT and the time spent in open arm in EPM test [15].
Relying on recent evidence indicating that stress-induced structural plasticity in amygdala neurons may provide a cellular substrate for enhanced anxiety triggered by chronic stress [104,105], we analyzed in current study the neurodegeneration in BLA. Unlike previous studies, which demonstrate that anxious-like behaviors induced by chronic stress, is related to BLA hypertrophy [106], our UCMS rats showed a neuronal loss in the BLA compared to control. Nevertheless, in supplemented rat’s neurodegeneration has a tendency to increase in this structure.
It is known that the stressor and the ability to effectively deal with it contribute to the development of anxious and depressive symptoms [70,71]. This relationship conduces to the coexistence of anxiety and depression symptomatology in the same patients; this fact has led to suggest that the comorbidity of anxiety and depression symptomatology is due to linked neurobiological abnormalities. In this respect, the current study suggests that the beneficial effects of AO found for depressive-like behavior, may also explain the anxiolytic effect observed in UCMS rats in the EPM test.
According to our knowledge, our work is the first one analyzing the effect of AO supplementation on emotional behaviors in Wistar rats submitted to UCMS. In summary, our data supports pharmacological and biochemical evidences for the antidepressant and anxiolytic like effects of AO. Prolonged administration of AO (10 mg/kg b.w.) reverses all the behavioral changes that occurred due to UCMS and restored corticosterone level in the plasma, oxidative status of amygdala and the neurons level in the CA3 subregion of rats’ hippocampus. Knowing the interesting chemical composition of AO and the complexity of emotional behaviors pathophysiology, which involves oxidative stress, neural plasticity, inflammatory pathways and the hyperactivity of HHS axis, we can consider the possible synergistic effects of these compounds that would be more beneficial than the use of each one. The current study suggests that antidepressant and anxiolytic like effects of AO in adult rats can be the result of modulation of brain antioxidant enzyme activities, the activation of hippocampal neurogenesis and the modulation of HPA axis activity.
Altogether, these data support the notion that a diet that includes AO or functional foods enriched with AO or with some of its specific compounds, which can be synthetized [107], may have beneficial effects in preventing or slowing the development of some emotional disorders related to stress such as depression and anxiety. Finally, our results indicate that AO could be an attractive neuro-pharmacological agent for the prevention (or the treatment) of neuropsychiatric disorders. The antidepressant effect of AO may be beneficial as a therapy for anxiety and depression. Thus, our results fortify the ethnopharmacological importance of AO in psychiatric disorders like anxiety and depression. However, more researches are needed to elucidate the exact mechanisms underlying its anxiolytic-andantidepressant- like effects in chronic stress model of rats.
Authors declare no actual or potential conflict of interest in this study.