Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2014) Volume 2, Issue 1
Prebiotics include nondigestible carbohydrate dietary additives and other biological components that stimulate the growth of one or more types of bacteria in the gastrointestinal tract that are beneficial to the host. These bacteria can inhibit colonization of pathogenic bacteria by producing antimicrobial substances and competing for niches within the gut. In this study, we evaluated the effects of Biolex® MB40 and Lieber® ExCel which are commercial prebiotics derived from brewer’s yeast cell walls. The two prebiotics were added to Genetically Modified Organism (GMO)- free chicken feeds with each group consisting of 1) control (no prebiotic), 2) Biolex® MB40 with 0.2% and 3) Leiber®
ExCel with 0.2%. Feeds were supplemented with the prebiotics during the 8 weeks of the entire experimental period. At 8 weeks, a total of 15 birds from each group were randomly selected for necropsy. The PCR-based Denaturing Gradient Gel Electrophoresis (PCR-based DGGE) technique was utilized to compare microbial populations in control and treatment groups. Feeds supplemented with either Biolex® MB40 or Leiber® ExCel prebiotics exhibited more consistent microbial populations (i.e. relatedness) compared to the control group. For Biolex® MB40 supplemented group, all samples were clustered with over 74% of relatedness. The Leiber® ExCel supplemented group exhibited 77% relatedness among 4 samples with the exception of one outlier. According to sequencing results, Bacteriodes salanitronis were consistently detected in all groups, and Barnesiella ciscericola and Firmicutes were identified in
both treatment groups. In addition, the class 1 integron gene prevalence was evaluated and frequencies of 93.3% for the control, 73.3% for the Biolex® MB40 and 73.3% for the Leiber® ExCel treatment group of chickens were observed. Last, Campylobacter concentrations were significantly lower in both treatment groups compared to the control group.
Keywords: Pasture chickens; Foodborne pathogens; Prebiotics; DGGE
Prebiotics have been used as alternative feed additives to improve gut health as well as for reducing pathogen colonization in various animal production systems including poultry [1-4]. Prebiotics include nondigestible carbohydrate dietary additives and other biological components that stimulate the growth of one or more bacteria in the gastrointestinal tract that are beneficial to the host [5]. In general, prebiotics are mixed as additives with feeds during the milling process so that all birds receive similar levels of prebiotics over the entire growth cycle. Prebiotics can be utilized preferentially by beneficial bacteria such as Lactobacillus and Bifidobacteria species [4,6-8], which leads to the production of lactic acid and Short Chain Fatty Acids (SCFA) both of which can be inhibitory to pathogens [3,9-12]. In addition, the presence of prebiotics can lead to the maintenance of a normal microbial population [10] which in turn can potentially inhibit the colonization of pathogenic bacteria through competitive inhibition [8,13]. The prebiotic compounds that are derived from yeast cell walls include beta-D-glucan and Mannan-Oligosaccharides (MOS) and these have been evaluated for their impact on poultry gut microflora [1,14,15].
Because of the potential impact of prebiotics on gut health as well as their Generally Recognized As Safe (GRAS) status, operators of alternative animal production systems including pasture raised chickens can safely use prebiotics during production. With respect to pasture raised chickens, the birds are generally grown outside on fresh grass, air and sunlight without antibiotic growth promoters, while prebiotics, probiotics and vaccines are allowed [16]. Several reports have evaluated the effects of pasture raised chicken growth performance and meat quality by not only chicken genotypes (fast-, mediumand slow-growing) but also for examining responses to prebiotics supplement [17-20]. Most studies have been focused on the bird growth performance and carcass yield [21], while the impacts of prebiotics on gut microflora in these chicken production systems with prebiotics have not been fully explored. Both Biolex® MB40 and Lieber® ExCel are commercial prebiotics derived from brewer’s yeast cell walls. The Biolex® MB40 contains high concentrations of beta-D-glucan and MOS that have been shown to bind substances detrimental to pathogenic bacteria as well as the Lieber® ExCel which is similar to Biolex® MB40. The effects of Biolex® MB40 have been evaluated using animal models such as lambs [22].
In conventional poultry production systems, antimicrobial agents have been utilized to increase bird performance and control pathogens but have come under increased regulatory scrutiny in more recent times [23]. Part of the controversy associated with the utilization of antibiotics is the need for additional antibiotic resistance baseline data from animals in production systems not fed antibiotics such as pasture flock poultry [24]. However, even if pasture raised chickens are not traditionally exposed to antibiotics, they still might acquire resistance genes from Gram-positive bacteria in the environment via horizontal gene transfer [24-26]. Integrons generally play an important role in the exchange of resistance genes among bacteria and can integrate the obtained resistance gene cassettes from the environment into microorganisms [27,28]. Rosser and Young (1999) identified 85 integron-positive Gram-nagative bacteria isolated from a river in the UK [29], and Roe and Pillai (2003) reported that 42 Escherichia coli (13%, n=322) isolated from irrigation water contained class 1 specific integron sequences [26].
Based on the above information, here we investigate the effects of two commercial prebiotics originated from yeast cell walls on both the normal microflora and pathogen carriage in pasture raised chickens in order to identify potential microbial population shifts and class 1 integron gene frequencies. First, the culture independent, PCR-based DGGE, was utilized to analyze and compare the microflora profiles of chickens receiving no prebiotics or either the Lieber® ExCel or Biolex® MB40. Second, integron gene frequencies were also investigated to generate initial baseline data on the presence of class 1 integrons in pasture raised chickens and to determine the impact of prebiotics on integron presence, if any. Last, the impact of prebiotics on the presence and concentration of Campylobacter and Salmonella carriage was assessed using culture-based techniques.
Chicken housing
A total of 147 day-of-hatch naked neck chicks were acquired from a local hatchery (Peterson Farms, Decatur, AR, USA). The birds were randomly distributed to 3 pens for a total of 49 birds per each pen. Birds had access to feed and water ad libitum for the duration of the experiment (8 weeks). In addition, the pens and birds were moved twice a week to fresh locations within the pasture that had not been previously used for poultry rearing purposes.
Two prebiotics were added to Genetically Modified Organism (GMO)-free normal chicken feeds (Hiland Naturals, Killbuck, OH, USA) in the starter, grower and finisher feeds in each group. Each group consisted of 1) control (no prebiotic), 2) Biolex® MB40 with 0.2% (Leiber GmbH, Hafenstraße, Germany), and 3) Leiber® ExCel with 0.2% (Leiber GmbH). Feeds were supplemented with the prebiotics for the duration of the experimental period. At week 8, a total of 15 birds from each group were randomly selected and euthanized humanely using CO2 gas followed by necropsies. The IACUC was exempted for this study since birds were grown off-campus farm and limited to microbiological evaluation. The ceca were extracted immediately for microbial analysis and stored at -20°C until DNA extraction. All remaining birds were processed and the corresponding weight for each bird was recorded.
Analysis of Campylobacter and Salmonella
The concentration of Campylobacter and the presence of Salmonella in each chicken were evaluated using individual bird cecal contents. Briefly, 9 ml of Buffered Peptone Water (BPW) was added to 1 g of cecal contents and 1 ml of the suspension was serially diluted for Campylobacter analysis using Campy-Cefex plates (BD Biosciences, San Jose, CA, USA). After a 48 h incubation under microaerophilic conditions at 42°C, suspect Campylobacter colonies were identified as small, mucoid, grayish and flat-shaped. Presumptive Campylobacter colonies were confirmed by phase contrast microscopy (Nikon instrument Inc, Melville, NY, USA) and latex agglutination immunoassay (Oxoid, Hampshire, UK). For Salmonella analysis, a 1 ml suspension was added to 9 ml of tetrathionate (TT) broth and incubated at 37°C for 24 h. After enrichment, 1 loop of TT broth was plating on xylose lysine tergitol 4 (XLT4; BD Biosciences) and brilliant green (BG; BD Biosciences) selective media to verify Salmonella prevalence. Salmonella suspected colonies appeared as either black colonies on XLT-4 or white opaque colonies with a red background on BG media and were confirmed by colony PCR.
DNA extraction
DNA was isolated from cecal contents of 15 birds from each group using the QIAamp Stool Mini Kit (Qiagen, Valencia, CA, USA) with some modifications to enhance DNA yields. In brief, 0.7 mm garnet beads (MO BIO Laboratories Inc., Carlsbad, CA, USA) were added to cecal contents to lyse cells by vortexing vigorously for 1 min. Samples were centrifuged to remove unhomogenized materials and the supernatant was transferred into a fresh 2 ml microcentrifuge tube containing 0.1 mm glass beads (MO BIO Laboratories Inc.). Horizontal vortexing was performed for 10 min and the samples were incubated in a heating block at 95°C for 6 min. The remainder of the DNA extraction protocol was performed according to the manufacturer’s instructions (Qiagen). The extracted DNA concentration and purity were measured using a NanoDrop ND-1000 (Thermo Scientific, Wilmington, DE, USA) and DNA was subsequently stored at -20°C for future analysis.
Class 1 integron gene analysis
Class 1 integron gene prevalence in DNA isolated from chicken ceca (n=45) was determined by a conventional PCR assay. The primer pair (Int 1F: GGC ATC CAA GCA GCA AG, Int 1R: AAG CAG ACT TGA CCT GA) for integron gene detection and PCR conditions used here have been previously described [29]. PCR products were sequenced using the ABI 3100 capillary analyzing system (Applied Biosystems, Foster City, CA, USA) and sequences were compared with database in GenBank using the Basic Local Alignment Search Tool (BLAST) algorithm.
PCR assay for DGGE
The conventional PCR assay was optimized using an MJ PTC 100 thermocycler (Bio-Rad, Hercules, CA, USA). A total reaction volume of 50 ul was comprised of 50 ng of template DNA, 800 nmol L-1 of each primer [P2 (ATT ACC GCG GCT GCT GG) and P3 (CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGG GCCTAC GGG AGG CAG CAG)] [30] (IDT, Coralville, IA, USA), 25 μl of Jump Start Ready Mix (Sigma, St. Louis, MO, USA), and DNase- RNase free water (Invitrogen, Carlsbad, CA, USA). The PCR conditions consisted of pre-denaturation at 95°C for 2 min, followed by 17 cycles of denaturation at 94°C for 1 min, annealing at 67°C for 45 s decreasing by -0.5°C per cycle to a touchdown temperature of 59°C, and annealing at 72°C for 2 min. The touchdown cycle was followed with 12 cycles of denaturation at 94°C for 1 min, annealing at 58°C for 45 s with a final extension step at 72°C for 7 min. The PCR products were confirmed on 1.5% agarose by gel electrophoresis and visualized on a transilluminator (Bio-Rad).
PCR-based DGGE
PCR-based DGGE was performed using 10 μl of the PCR products mixed with 5 μl of loading buffer. The samples were loaded into the wells of a polyacrylamide gradient gel composed of acrylamide:bisacrylamide (37:1) (Bio-Rad) with a 35% to 60% gradient of urea (Amersham Biosciences, Piscataway, NJ, USA) and formamide (Sigma). Electrophoresis was performed using a DCode Universal Mutation Detection System (Bio-Rad) in 1X TAE buffer at 59°C and 55 V for 17 h. The polyacrylamide gel was stained with SYBR Green (Cambrex Bioscience, Walkersville, MD, USA) in 1X TAE for 40 min with gentle shaking, destained in distilled water for 10 min, and viewed on a transilluminator. DGGE banding patterns among individual samples in each treatment as well as between treatments were analyzed using the Unweighted Pair Group Method with Arithmetic mean (UPGMA) algorithm to determine any correlation or relatedness.
DNA recovery from DGGE gel for sequence analysis
By comparing banding patterns among groups, common or specific bands were excised from the polyacrylamide gel for sequence analysis. Briefly, excised fragments were disrupted via a pinhole tube, transferred into 300 μl of TE buffer and incubated for 15 min in a heating block at 65°C to dissolve DNA. The suspension was transferred to a Spin-X® centrifuge tube (Corning, Tewksbury, MA, USA) and centrifuged at 16,000 g for 5 min to extract DNA from the polyacrylamide gel. In order to precipitate DNA, the filtrate was mixed with 900 μl of 100% ethanol, 133 μl of 7.5 mol L-1 ammonium acetate, 3 μl of glycogen (20 mg ml-1) and vortexed vigorously and incubated at -70°C for 1 h. The mixture was pelleted via centrifugation at 16,000 g for 15 min and the resulting pellet was washed with 70% of cold ethanol kept in -20°C. The isolated DNA was subsequently sequenced using an ABI 3100 capillary analyzing system (Applied Biosystems) and the sequences were compared with the database in GenBank using the BLAST algorithm.
Statistical analysis
Statistical analyses were performed using JMP® Pro 10.0 (SAS, Cary, NC, USA). One-way ANOVA was applied to determine statistical differences in body weight and Campylobacter concentrations in cecal samples among treatments. In all cases, statistical significance was set at a ≤ 0.05 and p<0.05. The phylogenetic tree was generated based on the DGGE banding patterns using the UPGMA algorithm.
Bird performance and pathogen concentration
Although there were no bird mortalities during studies, 3 birds escaped from control group and moved to Biolex® MB40 group (1 bird) and Leiber® ExCel (2 birds), respectively. In addition, there was no significant difference in Feed Conversion Ratio (FCR) between control and treatment groups (Table 1). The average bird weights by treatment before and after processing are shown in Table 1. Based on one-way ANOVA, there were no significant differences among treatments.
| Control | Biolex® MB40 | Leiber®ExCel | |
|---|---|---|---|
| Feed Conversion Ratio (FCR) | 2.47A | 2.37A | 2.46A |
| Body weight (8 weeks) | 6.00A | 6.02A | 6.19A |
| Body weight (after processing) | 4.43 ± 0.65A | 4.29 ± 0.59A | 4.34 ± 0.53A |
| Campylobacter* | 6.49 ± 0.36A | 6.07 ± 0.12B | 6.07 ± 0.1B |
| Salmonella | None | None | 1 chicken |
Table 1: Body weight (lbs), and Campylobacter levels and Salmonella (italic) presence of birds fed one of two prebiotics.
Campylobacter were detected in all cecal contents of the control and two treatment groups. Campylobacter concentrations in the control group exhibited 6.49 (log10CFU) per 100 mg and were significantly higher when compared to the two treatment groups (Biolex® MB40 (6.07) and Leiber® ExCel (6.07)) (Table 1). In contrast, Salmonella were only detected in cecal contents of one bird fed the Leiber® ExCel treatment out of a total of 45 samples (Table 1).
Class 1 integron gene analysis
The class 1 integron gene analysis revealed a positive level of 93.3% (n=15) for the control group, 73.3% (n=15) for the Biolex® MB40 supplemented group and 73.3% (n=15) for the Leiber® ExCel supplemented group, respectively. Overall, 80% (n=45) of the pasture raised chickens tested in this study contained the integron gene in their cecal contents. The sequenced PCR product was identified as a partial sequence of class 1 integron gene typically present in Escherichia coli (KF921523.1), Salmonella Heidelberg plasmid pSH1148 (JN983049.1), Aeromonas species (JQ837997.1), and Gram-positive bacteria including Staphylococcus aureus (KF687971.1).
Microbial population shifts in chicken ceca using PCR-based DGGE
All chicken cecal samples produced 233-bp amplicons via conventional PCR, and these products were subsequently used for DGGE analysis. DGGE analyses were performed to detect microbial population shifts in the ceca of birds supplemented with each one of the prebiotics (Biolex® MB40 and Leiber® ExCel) compared to control birds. Fifteen chicken cecal samples in the control group and 14 samples in both treatment groups (one sample for each treatment was discarded due to contamination) were utilized for DGGE analysis. Based on DGGE banding patterns, each group’s phylogenetic tree was generated by the UPGMA algorithm, which illustrates the correlation among individual chickens (Figure 1). Fifteen individual chicken cecal samples in the control were clustered with over 58% homology (Figure 1A). Each of the 14 cecal samples supplemented with Biolex® MB40 or Leiber® ExCel exhibited over 66% and 51% homology, respectively (Figures 1B and 1C). The Biolex® MB40 group exhibited more consistency with greater homology than the other two groups.
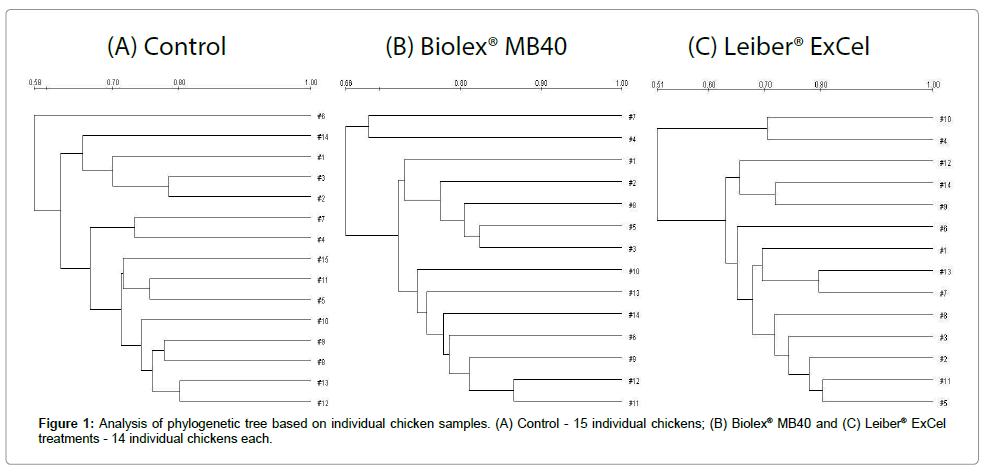
Figure 1: Analysis of phylogenetic tree based on individual chicken samples. (A) Control - 15 individual chickens; (B) Biolex® MB40 and (C) Leiber® ExCel treatments - 14 individual chickens each.
In order to compare the correlation between control and treatment groups, cecal samples in each group were pooled into 5 samples after comparing individual DGGE banding pattern similarities based on UPGMA analysis and DGGE was subsequently performed using the pooled samples (Figure 2). A phylogenetic tree was generated based on DGGE results and is shown in Figure 3. Interestingly, the phylogenetic tree of the control and treatment groups exhibited 3 distinct clusters in each group except one outlier occurring in the Leiber® ExCel treatment (Figure 3). In the control group, four sample banding patterns were over 68% related and one sample clustered in the Leiber® ExCel group with 70% relatedness. Feeds supplemented with either Biolex® MB40 or Leiber® ExCel prebiotics were more consistent compared to control group among pooled samples. For the Biolex® MB40 supplemented group, all samples were clustered within 74% of relatedness. The Leiber® ExCel supplemented group was 77% related among the 4 pooled samples except for the one sample which was an outlier (62%).
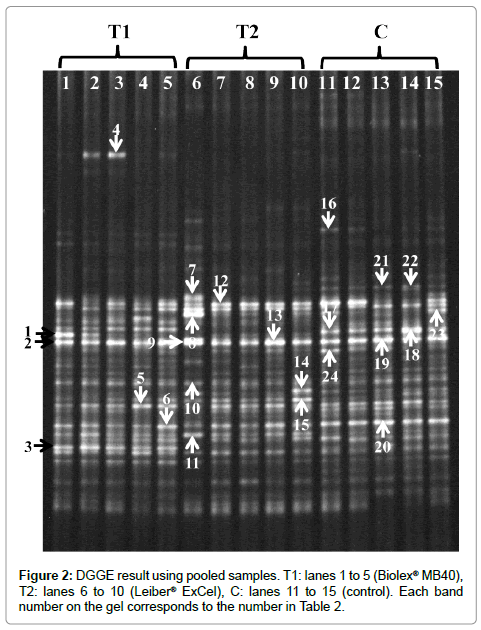
Figure 2: DGGE result using pooled samples. T1: lanes 1 to 5 (Biolex® MB40), T2: lanes 6 to 10 (Leiber® ExCel), C: lanes 11 to 15 (control). Each band number on the gel corresponds to the number in Table 2.

Figure 3: Analysis of phylogenetic tree based on DGGE results using pooled samples. T1: Leiber® MB40 (#1 to #5), T2: Leiber® ExCel (#6 to #10), C: Control (#11 to #15).
Sequencing
DGGE banding patterns showed high similarities within control and both treatment groups (Figure 2). However, some bands were specific within one group and the intensity of several common or specific bands were different among groups. For instance, band number 4 and 16 were specific for the Biolex® MB40 treatment and control groups, respectively (Figure 2). In addition, band number 2, 9, and 19 were common and yielded a fairly consistent intensity over all groups (Figure 2). These specific and common bands were excised from a polyacrylamide gel for sequencing analysis and identification results are shown in Table 2.
| Band No. | Identification |
|---|---|
| 1 | Helicobacter ganmani |
| 2, 9, 13, 19 | Bacteriodes salanitronis |
| 4 | Uncultured porphyromonadaceae |
| 6 | Barnesiella viscericola, Firmicutes |
| 10 | Barnesiella viscericola |
| 11 | Campylobacter jejuni, C. coli, C.lari |
| 14 | Uncultured rumen bacterium |
| 15 | Uncultured Rikenellaceae |
| 16 | Bacteriodes coprocola |
| 17 | Paraprevotella clara |
| 20 | Alistipes sp. |
| 12, 21, 22, 23, 24 | Uncultured Bacterium |
Table 2: Identification of DGGE bands via sequencing between control and two treatments
From the sequencing results, although several bands were identified as an uncultured bacterium (12, 21, 22, 23, and 24) and could not be sequenced (3, 5, 7, 8, and 18), the remainder of the bands were identified as a specific species with a high homology. Bacteriodes salanitronis (2, 9, 13, and 19) were consistently present in all groups, and Barnesiella viscericola and Firmicutes (6) were detected in both treatment groups. Interestingly, Helicobacter ganmani (1) and uncultured Porphyromonadaceae (4) were only present in Biolex® MB40 treatment group while Paraprevotella clara (17), Alistipes spp. (20) and Bacteriodes coprocola (16) occurred only in the control group.
Considering the prebiotic influences on the gastrointestinal microflora and the impact that microflora have on host health, it is important to determine whether supplemented prebiotics influence gastrointestinal bacteria composition and prevalence of foodborne pathogens. The purpose of this study was to evaluate the microbial population responses in slow growing (naked neck) chickens raised on pasture that were supplemented with one of two commercial prebiotics obtained from yeast cell walls including beta-D-glucan and MOS. Beta-D-glucan polysaccharides are composed of D-glucose monomers joined by glycosidic bonds and MOS have been used as nutritional additives to preserve gut health since they were initially introduced in the late 1980s [15,18,31]. It has been reported that the mannose sugar site in MOS can bind to Salmonella fimbriae and inhibit Salmonella colonization in the broiler intestinal cells and reduced levels of E. coli and Clostridium perfringens [1,15,31,32].
More recently, pasture raised chicken production has expanded due to increased consumer demands for local and sustainable foods [17-19]. Similar to conventional poultry production, operators of alternative poultry production systems look for ways to improve the health and performance of their birds, and supplementation of feed with prebiotics may be one way to accomplish this. Pelicia et al. evaluated the effect of prebiotics (MOS) extracted from Saccharomyces cerevisiae cell walls and probiotics (Enterococcus species) on pasture raised chickens and reported that the treatment group supplemented with both prebiotics and probiotics exhibited less mortality as well as a higher carcass yield than the control group [18]. Hanning et al. examined the effects of three dietary supplements (plum fibers, galactooligosaccharides (GOS) and Fructo Oligosaccharides (FOS)) on two breed broilers (Cornish White Plymouth Rock (CWPR) and Naked Neck (NN)) [21]. They concluded that CWPR and NN broiler fed with FOS and plum fibers exhibited higher body weight than the control group, respectively.
Antimicrobial agents have been used to control pathogenic bacteria in poultry and increase production; however, pathogens may share antibiotic resistance genes via horizontal transfer to overcome exposure to antibiotics [25]. Integron genes are considered important genetic components that can mediate the integration of antibiotic resistant mobile genes into target DNA of host cells leading to resistance against various antibiotics [33]. Among the 9 integron classes, the class 1 integron has been commonly identified in gastrointestinal tracts of animals, livestock and birds as well as Gram positive bacteria such as Staphylococcus species in poultry litter [25,27,33]. Furthermore, since most integron genes are present in a plasmid DNA or mobile elements such as transposons, they can be easily transferred to foodborne pathogens including Gram-negative bacteria via conjugation [25,33,34]. Although pasture raised chickens in this study were not treated with antibiotics, the identification of class 1 integron gene presence in cecal contents could potentially be associated with horizontal transfer of antibiotic resistant genes [3,25]. In this study, 80% (n=45) of cecal samples were positive by class 1 integron gene analysis. It would be of interest in future studies to collect environmental samples in proximity to the pasture raised birds to determine the presence of integrons and antibiotic resistant bacteria [34-36]. Their presence might explain the detection of integrons in the bird digestive tract via their relatively constant contact with these environments and organisms.
Microbial populations in birds can shift in response to alterations in feed additives and other factors [12,37,38]. In this study, the gastrointestinal bacteria Firmicutes and the Bacteriodes were identified in all groups (Figure 2 and Table 2). Firmicutes are 1) a phylum of bacteria presenting Gram-positive cell wall structure, 2) commonly present in gastrointestinal tracts, and 3) composed of over 250 genera including Bacilli and Clostridia [39]. The bacteria in the Bacteriodes genus are Gram-negative bacteria that utilize plant glycans as their main energy sources [40]. Moreover, members of the Bacteriodes genus are one of the predominant anaerobic bacterial groups found in chicken ceca [41]. In this study, Bacteriodes salanitronis was identified in all groups with considerable band intensity, and Bacteriodes coprocola was found only in the control group. Campylobacter species are commensal bacteria colonizing the gastrointestinal tract in poultry [42] and three species (C. jejuni, C. coli and C. lari) were detected in both treatment groups in this study based on a DGGE band sequencing.
Although the PCR-based DGGE technique has several limitations for analysis of whole bacterial populations in gastrointestinal tracts, this approach proved to be useful for comparing microbial population shifts influenced by prebiotic treatments. Furthermore, sequencing data provided additional information concerning specific species of bacteria associated with each treatment; however, the role these bacteria may play in the gut ecosystem remains unclear. In conclusion, microflora in both prebiotic supplemented groups is more consistent than the control group based on phylogenetic tree analysis. Also, further studies to analyze all microbial populations using whole genome sequencing for confirmation are warranted to precisely differentiate more subtle shifts in gut microflora both over time and as a function of diet.
Author SHP was supported on a partial graduate stipend by the Cell and Molecular Biology program at the University of Arkansas. Research funding was awarded to author SCR and KEG from Leiber GmbH, Hafenstraße, Germany.