Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2016) Volume 5, Issue 6
In this study 13 different accessions of medicinal and aromatic plant Cassia tora obtained from different agro climatic zones of Madhya Pradesh (Central India) were evaluated for its genetic variability using the random amplified polymorphic DNA (RAPD) markers. During RAPD analysis, amplification of genomic DNA of the 13 accessions by 10 selected primers yielded 88 bright and discernible bands, of which 86 were polymorphic with an average of 9.6 bands per primer and polymorphism obtained was 97.72%. The amplified products varied in size from 2,111 to 153 bp. The Jaccard similarity coefficient value ranged from 0.23 to 0.65 indicate that the distribution of variation is diverse. A dendrogram constructed based on the UPGMA clustering method revealed three major clusters, which are again divided into sub clusters. The result revealed rich genetic diversity among Cassia tora accessions obtained from different agro climatic zones of central India, which gives its suitability to grow and conserve anywhere at any environmental conditions.
Keywords: Polymorphism; Agro climatic Zone CTAB; RAPD – PCR; Cassia tora
Cassia tora Linn. belongs to family leguminous, is an important medicinal plant already reported in the ancient Ayurvedic literatures for curing many ailments. It is a well-known plant widely distributed in India and other tropical countries [1]. This is a shrub basically grows wild in the wasteland in India and called the “rainy herb”. It contains several medicinally important constituents like anthraquinones and Sennosides which are used in the formulation of many Ayurvedic cough syrups and expectorants and also used for various skin diseases such as ringworm, eczema, and scabies [2-4]. It also possesses a variety of biological/pharmacological activities such as Hypolipidemic, Antitumer, Antimutagenic, Antioxidant, Immunomodulatory, Antinociceptive, Antithelmintic, Antibacterial, Antifungal, Antihepatotoxic, Anti-inflammatory, Antigenotoxic, and anticoagulant activities [5-8].
Due to its high demand and utilization in the Ayurvedic preparations and Medications, its identification with respect to environmental influence has become increasing important for documentation of genetic information to overcome the disputes related to taxonomic identity, relations and authentication of species for future reference and protection of genetic diversity. Previous study of systematic and diversity analysis was performed using morphological characteristics and phenotypic traits, but they fail to serve as unambiguous markers due to the environmental influences [9,10]. To overcome the problem associated with phenotype based classification many molecular markers have been developed like Random Amplified polymorphic DNA (RAPD) [11], Simple sequence repeats (SSRs microsatellites) [12], Sequence tagged sites (STS) [13], Inter simple sequence repeat (ISSR) [14], Restriction fragment Length Polymorphism (RFLP) [15] and Amplified Fragment Length Polymorphism (AFLP) [16] to detect genetic diversity, genotype identification and genetic mapping amongst the species. Of these markers Random Amplified polymorphic DNA (RAPD), which involves PCR amplification using the arbitrary nucleotides, are a powerful tool for investigating genetic variation within species [17]. Random Amplified polymorphic DNA (RAPD) marker technique has many advantages over other molecular techniques such as relatively lower analysis, cost, it does not require previous genomic sequence information which makes it very simple and quick [11] and not influenced by any environmental conditions. Therefore, RAPD has been extensively used for population genetic analyses [18-20], DNA fingerprinting [21-23] and genetic diversity study [24] of various plant species, including many rare and endemic plants [25]. Sandeep Kumar et. al., investigated rich genetic diversity among Aloe vera (L.) accession from different agro-climatic zones of India by using SSR marker [26]. Shilpa et al studied the genetic diversity of an important medicinal plant Solanum trilobatum from south India using RAPD and ISSR markers [27]. Soni and Kaur analysed rapid in vitro propagation, conservation and genetic stability of Viola pilosa. Pruthvish and Chikkaswamy revealed the genetic diversity and relationships among mango varieties using RAPD molecular markers.
In the present study, we have been using RAPD markers to investigate the genetic relationship and distribution of genetic variation among population of medicinal plant Cassia tora, strictly distributed in central India. The molecular Markers can characterize plants with greater precision than biochemical parameters [28]. Among these, randomly amplified polymorphic DNA (RAPD) markers are efficient to assess genetic variation and extensively to evaluate natural genetic diversity in plant populations [23,29,30]. The purpose of the current study was to develop methods of DNA isolation without the use of liquid nitrogen to investigate genetic variation among different species of Cassia tora, medicinally important plants of central India.
Study area for population sampling
The plant material for the study comprises a total of 13 accessions, geographically distinct genotypes of Cassia tora, were collected from different agro climatic zones of Central India (Madhya Pradesh). For each sampled individual, fresh leaves (10 GM) were collected and placed in a sealed plastic bag with the appropriate label and stored in -20°C until subsequent DNA extraction and future use. The distance between the sampling points is more than 50 kms.
Genomic DNA extraction
Genomic DNA from young leaves was isolated from all the 15 accessions (Table 1) using the protocol of Doyle and Doyle (1987) with some modifications. Purification of DNA was done using the standard procedures. The extracted DNA was air dried at RT and finally, dissolved in 50 μl of TE buffer or deionised water (RNAse, DNAse free). Purity of total genomic DNA was checked using a UV Spectrophotometer (ND-1000) by calculating the A260/280 ratio [31] and running the samples on 0.8% agarose gel based on the intensities of the band when compared with Lambda DNA marker (100 bp ladder, used to determine concentration) by visualizing under UV light in Gel Documentation System.
RAPD-PCR analysis
RAPD profiles were generated using the PCR amplification of DNA extracted from fifteen different accessions of Cassia tora, with 10 single decamer random oligonucleotide primers (Bangalore Genei, India). Each amplification reaction mixture of 50 μl contained about 1 μl of genomic template DNA (50 ng), 25 μl of 2X red dye master mix, 2 μl of primer, (Bangalore Genei Pvt. Ltd., India) and 22 μl de-ionized water (RNAse, DNAse free). In negative control’s template DNA was replaced with nuclease free water. The reactions were performed in a Gradient Automatic Thermal Cycler (Eppendrof master cycle gradient, Hamburg, Germany) in the following cycles: holding at 94°C for 5 min, followed by 8 cycles of 94°C for 45 Sec, 55°C for 1 min and 72°C for 1.5 min and 35 cycles of 94°C for 45 Sec, 55°C for 1 min and 72°C for 1 min and a final additional extension at 72°C for 10 min and a holding temperature of 4°C. Amplified PCR products were electrophoretically separated in 1.2% (w/v) agarose gels, in 1 × TAE buffer at 60 V and stained with ethidium bromide (0.5 μg/ml). Gels with amplification fragments were visualized and photographed under UV light using Gel Documentation System (Alpha Innotech). The size of the polymorphic fragments was analyzed using lambda DNA (Eco R1-Hind III double digests) 100 bp DNA ladder was used as size standard molecular markers. Raw gel images were recorded through Alpha View Software. All the experiments were repeated thrice to ensure reproducibility. The best gels of the replicates were used for band scoring.
RAPD data analysis and scoring
In RAPD analysis, each fragment/band produced was treated as unit character. Only the primers that displayed reproducible, clear and intense bands were scored and their molecular weight was calculated using Alpha view software. For all accessions, amplified DNA fragments were converted into binary character matrices as (1) when present or (0) when absent. Polymorphic information content (PIC) values were calculated for RAPD primer employing the formula of Roldan Ruiz et al.:
PIC=2fi(1-fi)
Where fi=Frequency of amplified allele (present band) and (1-fi)=frequency of the null allele (absent band) of marker (I).
Marker index was calculated according to the formula MI=Product of PIC and the number of polymorphic bands per assay unit.
The data obtained were combined in a single matrix and evaluation of fragmentation patterns was carried out using a similarity index. The Similarity index (SI) values between the RAPD profile of any two individuals were calculated using the Nei genetic similarity index [32], on the basis of the equation, SI=2Nij/(Ni+NJ)
Where Nij is the number of common bonds shared between 2 samples I and j, Ni and NJ are the total number of DNA bands for genotypes I and j, respectively.
Using dice coefficients, a similarity matrix involving 15 accessions was generated by PAST (Paleontological statistics) software [33]. The similarity matrix data were subjected to an unweighted pair group method for arithmetic average (UPGMA) cluster analysis to generate a dendrogram using the average linkage procedure. The results were analyzed based on the principle that a band is considered to be ‘polymorphic’ if it is present in some individuals and absent in others, and ‘Monomorphic’ if present in all the individuals or accessions.
Amplifications were repeated twice to confirm the results.
In the present study a total of 15 accessions of medicinally important plant Cassia tora was collected from different geographical regions (Table 1). The genetic variability analyses amongst the Cassia tora genotype were investigated using Random Amplified Polymorphic DNA markers (Bangalore Genie). In this investigation, 10 random decamer oligonucleotide primers were used for fingerprinting of 15 accessions of Cassia tora. Of these 10 primers, the amplifications of only 6 primers (RPI-3, RPI-4, RPI-5, RPI-6, RPI-7 and RPI-8) were satisfactory and reproducible. The non-amplifications of the primers could probably be due to unavailability of the binding site for the primers. A similar non amplification of decamer primers in different plant species [34,35]. In RAPD profiling, a total of 84 clear and reproducible bands was produced, of which all 84 bands (100%) were polymorphic, which unambiguously discriminated 15 accessions, indicating the presence of a high degree of genetic variation in the studied accessions. The number of bands obtained per primer ranged from 5 (RPI-4) to 21 (RPI-3). The average number of polymorphic bands per primer was 14. The band obtained ranged in size from 153 to 2111 bp. The largest amplicon (2111 bp) was amplified by the primers RPI-3 and the shortest amplicons (153 bp) by RPI-6. The typical DNA fingerprints generated using RAPD primers are shown in Figures 1-4. The number of amplified products, number polymorphic fragments, percentage polymorphism, Polymorphic information, content, marker Index and size range of bands for the primer are given in Tables 2 and 3. Out of 6 primers all primers were found to be 100% polymorphic.
| S. | Place of | Accession ID | Geographical | S. | Place of | Accession | Geographical | ||
|---|---|---|---|---|---|---|---|---|---|
| No. | origin | location | No. | origin | ID | location | |||
| Latitude | Longitude | Latitude | Longitude | ||||||
| (N) | (E) | (N) | (E) | ||||||
| 1 | Khandwa | CT 1 | 22° 30' N | 77° 00' E | 9. | Bhopal | CT 9 | 23° 16' N | 77° 36' E |
| 2 | Raisen | CT 2 | 23° 15' N | 77° 5' E | 10. | Gunna | CT 10 | 24° 40' N | 77° 20' E |
| 3 | Sehore | CT 3 | 23° 12' N | 77° 00' E | 11. | Vidisha | CT 11 | 23° 32' N | 77° 51' E |
| 4 | Hosangabad | CT 4 | 22° 46' N | 77° 45' E | 12. | Indore | CT 12 | 22° 44' N | 75° 50' E |
| 5 | Harda | CT 5 | 21° 88' N | 77° 98' E | 13. | Dhar | CT 13 | 22° 35' N | 75° 20' E |
| 6 | Betul | CT 6 | 21° 88' N | 77° 98' E | 14. | Dewas | CT 14 | 22° 58' N | 78° 06' E |
| 7 | Jhabua | CT 7 | 22° 45' N | 74° 38' E | 15. | Ujjain | CT 15 | 23° 09' N | 75° 43' E |
| 8 | Allirajpur | CT8 | 22° 11' N | 74° 24' E | |||||
Table 1: Location of germplasm collection of 15 accessions of Cassia tora from different of Madhya Pradesh (Central India).
| S. No | Primer | Primer Accession no. Bangalore Genei | Total no. of brands | Size Range of Amplified Product (bp) | Total no. of polymorph hic bands | Polymor phism (%) | Monomor phism (%) | PIC=2 × fi × (1-fi) | MI=PIC × % polymorphism |
|---|---|---|---|---|---|---|---|---|---|
| 1 | RPI 3 | AM 773310 | 21 | 180-2111 | 21 | 100 % | 0 % | 0.41 | 8.61 |
| 2 | RPI 4 | AM 773769 | 5 | 400-1150 | 5 | 100 % | 0 % | 0.25 | 2.25 |
| 3 | RPI 5 | AM 773770 | 8 | 200-1555 | 8 | 100% | 0 % | 0.46 | 4.14 |
| 4 | RPI 6 | AM 773771 | 13 | 183-766 | 13 | 100 % | 0 % | 0.49 | 6.37 |
| 5 | RPI 7 | AM 773312 | 18 | 220-1400 | 18 | 100 % | 0 % | 0.49 | 8.82 |
| 6 | RPI 8 | AM 773773 | 18 | 153-1625 | 18 | 100 % | 0 % | 0.5 | 8.64 |
| Total | 6 | 84 | 86 | 600% | 0 % | 2.6 | 38.83 | ||
| Average | 14 | 14 | 100 % | 0 % | 0.43 | 6.47 | |||
PIC: Average polymorphic Information contents for polymorphic bands; MI: Marker Index
Table 2: Degree of polymorphism, PIC and Marker index for RAPD primers in 15 accessions of Cassia tora.
| Sample | CT-1 | CT-2 | CT-3 | CT-4 | CT-5 | CT-6 | CT-7 | CT-8 | CT-9 | CT-10 | CT-11 | CT-12 | CT-13 | CT-14 | CT-15 |
| ID | |||||||||||||||
| CT-1 | 1.00 | ||||||||||||||
| CT-2 | 0.53 | 1.00 | |||||||||||||
| CT-3 | 0.45 | 0.44 | 1.00 | ||||||||||||
| CT-4 | 0.44 | 0.43 | 0.48 | 1.00 | |||||||||||
| CT-5 | 0.26 | 0.27 | 0.29 | 0.36 | 1.00 | ||||||||||
| CT-6 | 0.33 | 0.33 | 0.33 | 0.38 | 0.41 | 1.00 | |||||||||
| CT-7 | 0.33 | 0.40 | 0.4 | 0.43 | 0.37 | 0.42 | 1.00 | ||||||||
| CT-8 | 0.32 | 0.43 | 0.4 | 0.40 | 0.39 | 0.45 | 0.47 | 1.00 | |||||||
| CT-9 | 0.39 | 0.38 | 0.42 | 0.49 | 0.35 | 0.59 | 0.42 | 0.49 | 1.00 | ||||||
| CT-10 | 0.38 | 0.41 | 0.31 | 0.45 | 0.44 | 0.46 | 0.38 | 0.53 | 0.47 | 1.00 | |||||
| CT-11 | 0.37 | 0.37 | 0.26 | 0.41 | 0.34 | 0.34 | 0.32 | 0.45 | 0.36 | 0.63 | 1.00 | ||||
| CT-12 | 0.32 | 0.26 | 0.26 | 0.47 | 0.43 | 0.36 | 0.4 | 0.44 | 0.39 | 0.49 | 0.38 | 1.00 | |||
| CT-13 | 0.35 | 0.28 | 0.39 | 0.35 | 0.30 | 0.41 | 0.40 | 0.43 | 0.38 | 0.41 | 0.41 | 0.43 | 1.00 | ||
| CT-14 | 0.4 | 0.28 | 0.36 | 0.39 | 0.23 | 0.36 | 0.4 | 0.49 | 0.54 | 0.34 | 0.41 | 0.4 | 0.53 | 1.00 | |
| CT-15 | 0.34 | 0.31 | 0.42 | 0.42 | 0.32 | 0.38 | 0.36 | 0.42 | 0.41 | 0.47 | 0.47 | 0.46 | 0.65 | 0.52 | 1.00 |
Table 3: Jacquard’s similarity coefficient among 15 different accessions of Cassia tora using RAPD data.
Figure 1: Collecting sites of 15 accessions of Cassia tora from different geographic regions of Madhya Pradesh (Central India).
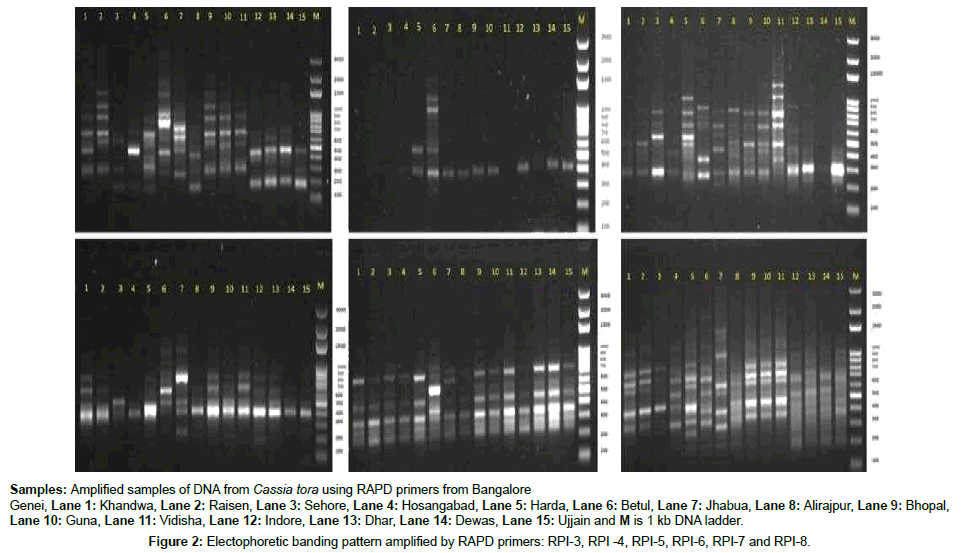
Figure 2: Electophoretic banding pattern amplified by RAPD primers: RPI-3, RPI -4, RPI-5, RPI-6, RPI-7 and RPI-8.
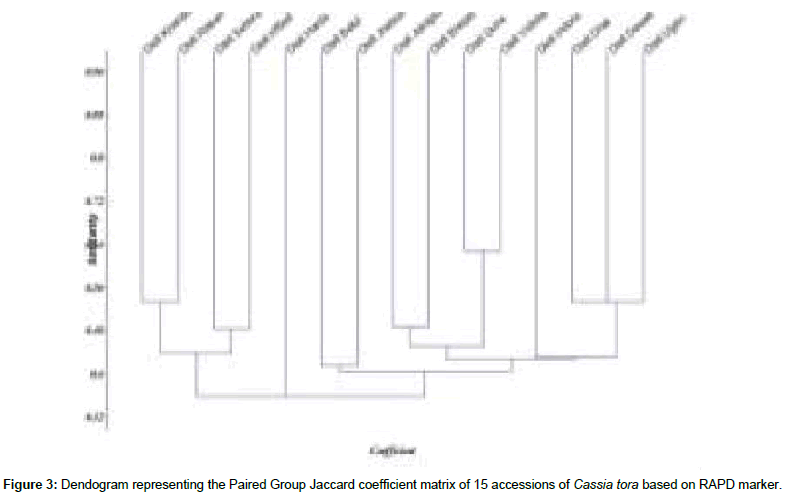
Figure 3: Dendogram representing the Paired Group Jaccard coefficient matrix of 15 accessions of Cassia tora based on RAPD marker.
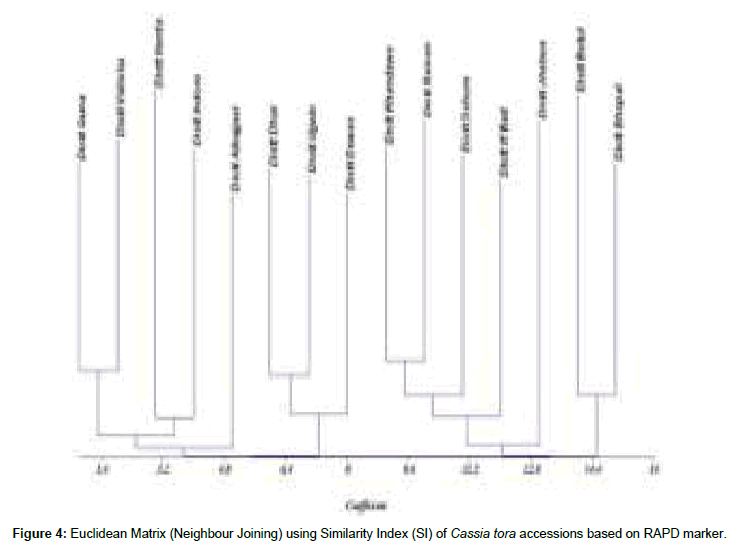
Figure 4: Euclidean Matrix (Neighbour Joining) using Similarity Index (SI) of Cassia tora accessions based on RAPD marker.
The genetic similarity index calculated using Nei’s similarity index value based on RAPD approach. The similarity index ranged from 0.23 to 0.65. The highest similarity value (0.65) was recorded between district Ujjain (CT-15) and district Dhar (CT-13), and the lowest similarity value (0.23) between district Dewas (CT-14) and District Harda (CT-5).
Cluster analysis was performed based on the Jaccard’s similarity coefficient matrices, calculated from RAPD data and dendogram was constructed using UPGMA (Unweighted Paired Group with Arithmetic Average). Out of 15 accessions of Cassia tora, accessions from district Harda (CT-5) and district Indore (CT-12) forms an out group and remaining 13 accessions were divided into four major groups and these clusters were further divided into subgroups. First cluster grouped together accession from collected from District Khandawa (CT-1), Raisen (CT-2), Sehore (CT-3) and Hoshangabad (CT-4). The second cluster contained accession from district Betul (CT-6) and Jabua (CT- 7). Accessions collected from district Alirajpur (CT-8), Bhopal (CT- 9), Guna (CT-10), Vidisha (CT-11) formed the third cluster and forth cluster contained accession collected from district Dhar (CT-13), Dewas (CT-14) and Ujjain (CT-15).
The plant Cassia tora is highly important for preparation of herbal formulations, it is very important to conserve the plant in situ and also maintain its genetic diversity. The level of genetic diversity and the pattern of genetic structure in plant population are mainly related to the species’ life history and breeding system and knowledge of this pattern is necessary for management, use and conservation of Biological Diversity [36]. In present study high level of genetic diversity was observed between the accessions, which could be a result of spontaneous mutations, which are quite common in the species, along with selection, geographic and environmental factors, which make the local population of this species an important genetic resource [37,38]. High level studies of genetic diversity amongst the sweet potato accessions [39,40]. The accessions CT-5 and CT-12 have been placed distantly from the rest of the accessions suggesting a high level of diversity present compared to the other accessions or they may be genetically highly heterogeneous in nature, which may be due to the structural changes in DNA such as deletion, inversion, translocation and mutation and changes in the genomic constitution of the species. Molecular analysis of Cassia tora reveals that there is no correlation between the geographical origin and the cluster obtained, which favors the hypothesis of the existence of a common origin for all Cassia tora accessions due to the selection pressure induced by the natures tale. Similar observations have been made earlier in the case of Cymbopogon [38], Tinospora cordifolia [41], Ocimum sanctum [39]. The high diversity of Cassia tora indicates the adoption to the environment, where it can be propagated easily and can be conserved and utilized. Comparing our result with other work on the study of molecular polymorphism in Cassia tora species by RAPD marker shown that these markers are even an efficient tool for differentiation of closely related of Cassia, Senna and Chamarcrista [22]. From the previous reports it was observed that there were many taxonomic mis classifications of plants observed, which by the advent of these molecular markers were reclassified resolving the previous confusions [42]. Use of RAPD primers has also been successful in resolving the phylogenetic relation between the Rosa species accessions by arranging them in the specific taxonomic groups. The germplasm collection of rice accessions have analyzed and classified then using RAPD marker [43]. The RAPD markers are superior to all other markers on the basis of their capacity to reveal more informative bands in single amplification, they are simple, require no prior information of sequences, cheap to analysis and very low amount of DNA is required for amplification. We know that the genetic diversity is very important for sustainability of plant population [44]. Also that the change in genetic diversity within the population may occur due to the changes in the complex genetic structuring, which is influenced by many factors such as the long-term evolutionary history of the species (e.g., shifts in distribution, habitat fragmentation, and/or population isolation), mutation, genetic drift, mating system, gene flow, and selection [45,46]. In the conservation management of a species, knowledge of interspecies genetic variations may help to assess extinction risks such as inbreeding and evolutionary potential in a changing world [47]. Effective conservation of a vulnerable species depends largely on the knowledge of patterns of genetic variation. For example, the spatial structure of genetic variation can provide information for sampling strategies for ex situ or in situ conservation [48].
This study was an attempt to establish the genetic diversity within Cassia tora from different geographical regions with RAPD markers. The present study shows the usefulness of RAPD analysis in distinguishing the Cassia tora species from-different localities, particularly the identification of the specific banding pattern may be considered important in the identification and for conservation of species and may lead to planning of a better management of conservation program in the country. High levels of polymorphism found in the present work could also pave the way for detailed research to understand all the aspects of this divergence.
The authors express heartfelt thanks to Prof Pramod K. Verma, DG MPCOST for the facilities and the financial assistance given at MP Council of Science and Technology, Vigyan Bhawan, Nehru Nagar, Bhopal.