Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Short Communication - (2015) Volume 3, Issue 2
β-amino butyric acid (BABA), applied as inducer of resistance to the seeds of susceptible cultivar of pearl millet [Pennisetum glaucum (L) R Br] against downy mildew [Sclerospora graminicola (Sacc) Schroet]. Infection offered protection to the host operative through vegetative and reproductive growth. Biochemical studies identified a close association between BABA-treatment and accumulation of defence-related proteins like phenylalanine ammonia lyase, peroxidise and β-1,3-glucanase. Studies on phenotypic host defence identified modifications in host cell wall leading to papillae formation with hydroxyproline-rich glycoproteins (HRGPs) as its major constituent. The HRGPs were identified as extensions by use of MAC 265 monoclonal antibodies. PgMPK4 (mitogen-activated protein kinase) was identified as the crucial signal transduction module signalling of pathogen cues. This increased accumulation of PgMPK4 mRNA and kinase activity in BABA-treated samples and further kinase inhibitor treatments providing critical evidence of PgMPK4 involvement in the JA- and SA mediated expression of defence genes, it may be assumed that BABA-induced resistance could be a JA/SA mediated phenomenon.
Keywords: BABA; Inducer of resistance; Pearl millet; Defencerelated enzymes; HRGPs; PgMPK4
A phenomenon of resistance details a host defence mechanism wherein plant, without changing its basic genetic constitution, utilizes its own defence mechanism to restrict pathogen ingress and spread with surface-localized, pattern-recognition receptors (PRPs) recognizing conserved pathogen-associated molecular patterns (PAMPs) [1] in constitutively resistant hosts. Further, synthetic and biological agents with a capacity to control a large variety of plant diseases, termed inducers of resistance are reported to be effective against a broad spectrum of pathogen attack viz., oomycetes, fungi, bacteria to viruses. Phenotypic similarities in inducer treated plants were comparable to the constitutively resistant cultivars by expression of hypersensitive response, trailing necrosis, wall strengthening by formation of papillae with lignifications [2]. Some chemical inducers thoroughly investigated include 2,6-dichloroisonicotinic acid (INA), benzo-(1,2,3)-thiadiazole- 7-carbothioic acid S-methyl ester (BTH; BION-commercialized tradename) credited to their capacity to interfere salicylic acid pathway [3]. In the very recent past, DL-β-aminobutyric acid (BABA), a rare non-protein amino acid identified in the root exudes of tomato plants [4]. BABA was identified as interesting due to its close relation to the neurotransmitter gama-amino butyric acid, a bioactive molecule of the central nervous system of animals. In contrast to BABA, GABA (γ-amino butyric acid) has been well-documented in plants as a naturally occurring molecule [5]. Some 50 years ago, Papavizas and Davey and Papavizas [6,7] recorded the protection offered by BABA in host pea plants to the oomycete pathogen Aphanomyces euteiches effectively reducing the root rot severity. A broad spectrum of activity due to disease-causing organisms such as virus, bacteria, oomycetes, fungi and nematodes supports a role for this molecule as an inducer of resistance and not a biocidal agent [8,9]. In India pearl millet [Pennisetum glaucum (L) R Br] finds a rare place as staple food for the poorer section of the society in dry regions like Rajasthan, however the yield is affected by the oomycetous biotropic fungus, downy mildew pathogen Sclerospora graminicola (Sacc) Schroet. A case study to elucidate the protective effect of use of BABA as inducer of resistance by applying it to the seeds, we took advantage of this pearl millet/ downy mildew pathogen as model system and have tried to evaluate the resistance offered by phenotypic observations, biochemical and molecular studies.
Effect of seed treatment and induction of resistance by β-amino butyric acid (BABA) in pearl millet
Studies identified seed treatment with BABA (50 mM, 6 h) [10] influenced seedling vigour and protected the seedlings from downy mildew disease up to 75%. Quantification of downy mildew pathogen in these plants by ELISA indicated statistically significant reduction of fungal biomass in susceptible plants protected by this inducer. A 2-day gap between the inducer treatment and challenge inoculation was identified for build-up of resistance, if further increased resulted in gradual decrease in protection. Further resistance induced was confirmed to be systemic in nature as BABA-treated plants recorded resistance to any further challenge inoculation at a site distant from the site of inducer treatment.
BABA protection was operative through reproductive phase of the host
Pearl millet plants are generally prone to infection by sexually produced seed, soil-borne oospores and asexually produced sporangia. The critical period for protecting these plants from sowing seeds to completion of panicle formation to tillers, lie in the range of 25-35 and 30-36 days, respectively. BABA-induced resistance durability was tested in plants raised from seeds treated with the inducer and identified as resistant, by second challenge inoculation with the downy mildew pathogen at tillers and inflorescence axes suggested induced resistance was operative through vegetative and reproductive growth of these plants.
Mechanism of BABA induced resistance
The mechanism by which BABA acted as an inducer was credited by its capacity to diffuse through the microscopic ruptures caused by imbibition of seeds or by initiation of interaction upon seed germination and diffusion into coleoptiles and radicals by nondifferentiated tissues of the germinating embryos. Further it could act by interfering with normal amino acid and/or protein synthesis affecting the metabolism of the host rather than the fungus, thus making the interaction incompatible. A concentration dependent phenomenon of resistance suggested the possible occurrence of specific receptors for BABA for its binding. Our method of induction of resistance by root treatment, including protection in plants from challenge inoculation with the downy mildew pathogen at the leaf-whorl suggested acropetal translocation of BABA possibly through xylem from the roots and a systemic nature of resistance involved.
BABA induced resistance is associated with accumulation of defence-related proteins
Further studies attempted to elucidate the biochemical basis for this protection since plants defence is attributed to early recognition of invading pathogen and initiation on an appropriate signalling process. Chemical barriers in form of lignins, phenylpropanoids and toxic phytoalexins by triggering phenylpropanoid pathways, activating phenyl alanine ammonia lyase (PAL) [11] are reported. During an incompatible interaction, a specific activation of PAL-dependent defence response was observed [12] along with its association in reducing the rate of pathogen multiplication and its further ingress [13,14]. In our studies, a primed increase of PAL in BABA-treated seedlings and its further significant increase after inoculation with S. graminicola was observed, an indication of incompatible environment created by this molecule in previously susceptible host [15]. Another enzyme, peroxidise with various physiological functions include oxidation of hydroxycinnamyl alcohols into free radical intermediates, phenol oxidation, polysaccharide cross-linking, cross-linking of extension monomers and lignifications. Elevated level of peroxidise recorded in susceptible pearl millet subjected to seed-treatment with BABA indicates creation of incompatible situation for downy mildew pathogen. Hydrolases like β-1,3-glucanase identified with a major role during seed germination resulting in removal of cell wall or catabolism of storage polymers may also provide protection to the host by directly degrading fungal cell wall components with a role in stimulating other host defence mechanisms viz., phytoalexin accumulation [16]. A close association was found between BABA-induced protection and increased accumulation of β-1,3-glucanase.Isoelectric focusing studies followed by direct detection of the β-1,3-glucanase isoforms (pI 9.6 and 8.8) on the gel suggested differential expression of this enzyme in pearl millet during pathogenesis by S. graminicola. Prevention of ingress by the pathogen was attributed to the possible modification of host cell wall. Cell wall structures are reported to dynamically change with physiological fluctuations as a response to external stimuli. Hydroxyproline-rich glycoproteins (HRGPs), as an important plant cell wall structural component are induced in response to pathogen attack. Increased accumulation of hydroxyproline rich glycoprotein (17 kDa molecule) as analyzed by western blot analysis with MAC 265 monoclonal antibodies [originally isolated by Bradley et al., [17] based on its recognition of interface proteins in pea-rhizobium symbiosis] quantitated to be comparable to the highly resistant cultivars [15,18].
Involvement of mitogen-activated protein kinase cascade in BABA induced resistance
Prior to phenotypic and biochemical mechanisms detailed above, transducing environmental (presence of pathogen) cues into intracellular responses include mitogen-activated protein kinase (MAPK) cascades. MAPKs cascades involve signal transduction via three types of reversible phosphorylated kinases leading to the phosphorylation of substrate proteins, whose altered activities mediate a wide array of responses regulating numerous processes in stress and hormonal responses, innate immunity and development. The kinase activity assay measure phosphorylation of protein substrate with canonical serine-proline or threonine-proline phosphorylation site with myelin basic protein (MBP) as preferred substrate. The cascades may share kinase components maintaining spatio-temporal constraints, protein-protein interaction resulting in cross inhibition, feedback control and scaffolding. Studies by genetic analysis identified dominant MAPK (MAPKsMPK3, 4 and 6; MAP2Ks, MKK1, 2, 4 and 5.The first reports of plant mitogen-activated protein kinase (MAPKs) indentified MsERK1 in alfalfa and D5 kinase in pea [19]. Plant MAPKs with TDY or TEY phosphorylation motifis in their activation loop are similar to ERK kinases reported in signal transduction pathways in animals [20]. Studies by immune-blot and immune-precipitation analysis confirmed the activated 48 kDa (PgMPK1) and 44 kDa (PgMPK2) proteins in pearl millet/downy mildew pathogen model system as orthologs of SIPK and WIPK, respectively [21]. A full length cDNA of Group B MAPK (PgMPK4) was isolated from pearl millet resistant cultivar located in the vacuole. Sequence alignment of PgMPK4 identified an n-terminal ATP-binding region with glycinerich loop and b-sheet along with characteristic activation loop of TEY for phosphorylation between subdomains VII and VIII. Among a few tested elicitors of resistance, BABA was also identified as one with a capacity to activate PgMPK1 [21]. Invading pathogen with a capacity to trigger biosynthesis of Jasmonic acid, salicylic acid and ethylene resulting in induction of an array of defence-related genes provides a model system for identification of signal transduction pathway [20] activated upon application of inducer [22] proposed PgMPK4 from pearl millet could be involved in the JA and SA mediated pathway by the way of PgMPK4 inhibitor treatment studies involving expression three defence genes, lipoxygenase, catalase 3 and polygalacturonaseinhibitor protein. As earlier studies involving resistance induced to a susceptible cultivar upon BABA-treatment also exhibited the presence of PgMPK4 prominently [21] may be it can be assumed that BABAinduced resistance could be a JA/SA mediated phenomenon (Figure 1).
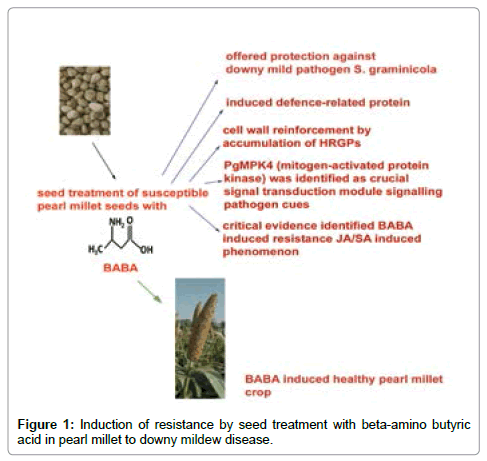
Figure 1: Induction of resistance by seed treatment with beta-amino butyric acid in pearl millet to downy mildew disease.
Induction of resistance by BABA in pearl millet resulted in increments in both vegetative and reproductive parameters. Further post-inflectional protection offered by seed treatment resulted in rapid biochemical alterations in the host viz., induction of defence-related enzymes, creating an incompatible environment for S. graminicola. Physical barriers in form of papillae at the sites of pathogen entry due to cross-linked HRGPs prevent its ingress. Presence of pathogen cues to the host by group B MPK gene (PgMPK4) from resistant pearl millet cultivar induced in response to S. graminicola infection was also induced in BABA-treated pearl millet as a prominent response. MPK cascade inhibitor treatments showed that PgMPK activity is involved in SA- and JA-induced activation of defence genes [lipoxygenase (LOX), catalase 3 (CAT3) and polygalacturonase-inhibitor protein (PGIP)]. As PgMPK was prominently expressed in BABA-treated pearl millet among the inducers tested, it may be assumed that BABA-induced resistance could be a JA/SA mediated phenomenon.
We acknowledge the recognition of University of Mysore as an Institution of Excellence by Government of India with financial support from the Ministry of Human Resource Development and University Grants Commission, India. This work was financially supported by the CSIR and UGC, New Delhi, India. The European Molecular Biology Organization is thanked for a short-term fellowship to MP. JM thanks the Novo Foundation for support from the University of Copenhagen Denmark-India program.