Enzyme Engineering
Open Access
ISSN: 2329-6674
ISSN: 2329-6674
Research Article - (2013) Volume 2, Issue 2
Keywords: Lipase, Rhizopus oryzae, Purification, Characterizatio,Low molecular weight
The quest for developing an environment friendly approach for synthesis of different products of human interest has lead to the employment of enzymes in varied industrial processes. Lipases, one of the major enzymes have proven to be extremely versatile. They have been extensively used in a number of industrial sectors including food, detergent, oleochemical, pharmaceutical, cosmetics, textiles etc. In recent years, research on lipases, mainly of microbial origin, has increased because of their great commercial potential [1,2].
Lipases are carboxyl-esterases acting on long chain acylglycerides. They possess several unique features viz. enantioselectivity, chemoselectivity and regioselectivity [3] which make them one of the prominent biocatalysts. Being ubiquitous in nature lipases are found in animals, plants and microorganisms. Since the industries look for lipases with desired properties for its efficient applications, new sources of enzyme are being explored. Fungal lipases are preferred due to easy recovery of enzyme from the fermentation broth [1]. Fungal species widely used for lipase production include Candida sp., Mucor sp., Rhizopus sp., Aspergillus sp., Penicillium sp. etc [2].
Apart from this certain degree of purity of the enzyme is essential for several biotransformation processes which necessitate development of efficient purification strategies. Varied purification schemes have been proposed based on the prime requirement of attaining maximum purity in lesser number of steps for process economization. Most purification processes employ concentration step, followed by chromatographic separation which can be single or multiple. Most of the conventional purification attained through multistep processes has been found to yield 2-20% of enzyme [4]. For large scale application purification strategies are required which are fast, inexpensive, high yielding and simple in operation.
The suitability and applicability of the purified enzyme in different industries is determined by a study of effect of various physical and chemical factors that help to understand its nature and properties [5]. Lipases derived from different sources show variations in their properties like thermostability, pH stability, and effect of metal ions, solvents and modifiers on the enzyme activity. For example different Aspergillus sp. lipases show variations in their characteristics. Aspergillus carneus had optimum pH of 9 [6] whereas Aspergillus niger showed pH 7 as its optimum pH [7]. Thus characterisation of enzymes is essential for widening its applicability.
The present study deals with purification of lipase produced by Rhizopus oryzae NRRL 3562 through solid state fermentation in minimal steps with the aim of increasing the yield as well as fold purification. Different biochemical characteristics of the enzyme have been explored and compared with other fungal lipases. The lipase was found to be smallest among the fungal lipases of various Rhizopus sp. till date and showed distinct biochemical properties which suggest its usefulness in transesterification processes.
Materials
p-Nitrophenyl esters (pNP-esters) and standard peptide markers for gel electrophoresis (Molecular weight of 16.95, 14.44, 10.6, 8.16, 6.21, 3.48 kDa) were procured from Sigma Alderich (St. Louis, MO, USA). SEPHADEX G-100 and solution of ampholytes (pH 3-10) were purchased from GE Healthcare Biosciences (Pittsburgh, PA, USA) and Bio-Rad (Hercules, CA, USA), respectively. AMICON CELL with 10 kDa molecular weight cut off was acquired from Milipore (Billerica, MA, USA). All the reagents were of analytical grade.
Microorganism
In the present study Rhizopus oryzae NRRL 3562, previously isolated from local soils of IIT Kharagpur, India was selected for lipase production. It was maintained on potato dextrose agar slants at 30°C with regular subculturing.
Enzyme production by solid state fermentation and extraction
For enzyme production wheat bran (25g) moistened with Czapekdox medium (1:1 ratio) supplemented with dextrose and coconut oil, was used as substrate. The flask with the substrate was then autoclaved and inoculated with a spore suspension (106 spores/ml) prepared from 5 day old culture slant and incubated at 30°C, 90% humidity for 6 days. Post incubation for extraction of enzyme, distilled water was added to the fermented biomass, which was then allowed to stand for 2 h at room temperature. Following this the contents were squeezed through cheese cloth and the extract was centrifuged for 15 min at 5000 rpm. The supernatant was collected and used for lipase assay, protein estimation and purification.
Assay of lipase
Lipase assay was done using standard method developed by Pencreach and Baratti [8] using p-Nitrophenyl palmitate (pNPP) as substrate. One unit (IU) of lipase activity equals to the amount of enzyme that releases 1 μmole of p-nitrophenol per minute under the assay conditions.
Protein estimation
Protein estimation was carried out following Bradford [9] method using bovine serum albumin as internal standard.
Purification of lipase
The crude extract was concentrated by two step ammonium sulphate precipitation. First ammonium sulphate was added to the saturation of 20% followed by centrifugation for 20 min at 8000 rpm. The pellet was discarded and supernatant was used for further precipitation. 55% of ammonium sulphate was added to the supernatant and thereafter the contents were kept overnight at 4°C and then centrifuged for 20 min at 8000 rpm. The pellet obtained after centrifugation was dissolved in 20 mM phosphate buffer (pH 7). The resuspended pellet was used for lipase assay and total protein estimation. The concentrated enzyme was dialyzed overnight against the same buffer. Post dialysis the enzyme solution was subjected to gel filtration chromatography (GFC) using SEPHADEX G-100 packed column (50 cm X 1.6 cm) previously equilibrated in 20 mM phosphate buffer (pH 7). Elution was done using the same buffer at the rate of 1ml/min. Fractions of 1.5 ml volume were collected and further assayed for presence of protein and lipase. Fractions showing lipase activity were pooled together.
Determination of lipase purity through native PAGE and zymography
Purity of the lipase enzyme was analysed by native polyacrylamide gel electrophoresis (native PAGE) using a 15% separating and 5% stacking gel [10]. The GFC pooled active fraction was concentrated using AMICON CELL and then subjected to gel electrophoresis. Coomassie Brilliant Blue staining was employed for visualization of bands. Activity of lipase was detected by zymography following method developed by Castro-Ochoa et al. [11].
Determination of molecular weight and pI
For determination of molecular weight sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) was carried out using a 20% separating and 5% stacking gel [10]. Molecular weight was estimated by comparing the relative mobility of purified enzyme with that of standard peptide markers ranging from 16.95-3.48 kDa.
The pI was determined by isoelectric focussing using ROTOFOR (Bio-Rad, Hercules, CA, USA). After 4 h run at 4°C, fractions were collected and checked for the enzyme activity.
Effect of temperature on activity and stability of lipase
The activity of the enzyme was assayed at varied temperatures from 30-90°C. For checking the stability of enzyme, purified enzyme was incubated at different temperatures varying from 30°C to 90°C. The incubation was done for 1 h and then the residual activity of the enzyme was measured using pNPP as substrate.
Effect of pH on activity and stability of lipase
To determine the effect of pH on enzyme activity, assay of the enzyme using pNPP as substrate was done at varied pH ranging from 2-10. The buffers used were Glycine-HCl (2-3), Acetate (4-5), phosphate (6-7), Tris-HCl (8) and Glycine-NaOH (9-10). All the buffers used were of same strength, 100mM. pH stability was studied by incubating the purified enzyme at different pH varying from 2-10. Incubation was done for 1 h at 30°C and then the residual activity of the enzyme was measured.
Determination of half life of lipase at pH 7, 30°C and 50°C
The half life of lipase was determined by incubating the enzyme at pH 7, 30°C and 50°C and measuring the residual activity of the enzyme at regular time intervals.
Determination of substrate specificity
The enzyme assay was carried out using p-Nitrophenyl acetate (pNPA), p-Nitrophenyl butyrate (pNPB), p-Nitrophenyl caprate (pNPC), p-Nitrophenyl laurate (pNPL), p-Nitrophenyl palmitate (pNPP) and p-Nitrophenyl stearate (pNPS) to determine substrate specificity.
Effect of metal ions on stability of lipase
Effect of different metal ions (Cu2+, Ca2+, Mg2+, Na+ , K+, Zn2+ , Hg2+, Pb2+, Co2+) at 1 mM concentration on the purified enzyme was determined by incubating the enzyme at 30°C in presence of the metals. The incubation was done for an hour and then the residual activity of the enzyme was measured using standard method employing pNPP as substrate against a control without any additions.
Effects of surfactants and inhibitors on stability of lipase
To study the stability of the purified lipase in different surfactants (SDS, CTAB, Triton-X-100, Tween- 20, Tween-80) and inhibitors (EDTA, urea, DTT and guanidium HCl) the enzyme was incubated in respective chemicals at 1 mM concentration, 30°C for an hour and then the residual activity was measured.
Effect of solvents on stability of lipase
Stability of the purified lipase in different solvents was studied by incubating the enzyme in presence of 10% solvents (methanol, ethanol, propanol, isopropanol, butanol, acetone, diethyl ether and hexane) for an hour at 30°C and then the residual activity was measured using pNPP as substrate.
Purification of lipase
The present work provides a two step purification of lipase from R. oryzae NRRL 3562 with high yield as well as fold purification. Summary of the purification has been given in Table 1 The methodology followed here has three major advantages. Firstly, number of steps has been significantly reduced to two steps as compared to earlier reported studies, four steps [12,13]; six steps [14] etc. Secondly, increased yield of 51.36% has been attained which is considered to be improved one compared to other reported literature [7,15-17]. Finally, notable increase in fold purification up to 37 times has been achieved, which is much higher than earlier findings [7,15-17]. Although, the recovery (63%) and fold purification (53.8) of the enzyme was higher for Aloulou et al. [18] and Kambe et al. [7] respectively, yet their corresponding fold purification [18] and yield [7] was found to be lower than the present study. Thus, in this case successful purification and recovery was attained in lesser steps and simpler process. Purification to homogeneity was confirmed by the presence of a single band in the native PAGE. Presence of a single orange fluorescent band in zymographic study confirmed the lipolytic activity of the purified protein [Supplementary figure 1] The enzyme was further used for biochemical characterization studies.
| Purification steps | Total Activity (IU) | Total Protein (mg) | Specific Activity (IU/mg) | Fold Purification | Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 980.20 | 80.51 | 12.17 | 1.00 | 100 |
| Ammonium sulphate precipitation | 854.24 | 7.98 | 107.05 | 8.80 | 87.15 |
| Sephadex G-100 | 503.43 | 1.12 | 450.29 | 37.00 | 51.36 |
Table 1: Purification of lipase from Rhizopus oryzae NRRL 3562.
Determination of molecular weight and isoelectric point of purified lipase
One of the unique aspects of the presently studied lipase is its molecular weight. It has been found to be 14.45 kDa (Figure 1) which is smallest among fungal lipases especially of Rhizopus sp. till date reported [12,13,19-22]. A 20 kDa lipase was reported in fungus Yarrowia lipolytica by Yadav et al. [23]. Recently, a small lipase of 17 kDa produced from Rhizopus sp. has been reported by Kantak and Prabhune [24]. Though small lipase of 11 kDa has been reported in bacterial system by Castro-Ochoa et al. [11], presence of such small lipase has not been so far reported in fungal system.

Figure 1: SDS-PAGE: Lane 1 GFC pooled lipase purified to homogeneity; Lane 2 standard pepetide markers.
Isoelectric focussing, showed localization of the enzyme at pH 6.5, concluded to be its isoelectric point. The pI of lipase from different sources has been found to be varying [12,20,21]. The enzyme’s pI is found to be similar to lipase derived from Mucor sp. by Abbas et al. [25].
Effect of temperature on activity and stability of lipase
Temperature plays a crucial role in maintenance of structural stability and activity of enzymes. As shown in Figure 2A the purified enzyme was found to have the optimum temperature range of 30-40°C (100%). 80% retention of activity was observed at 50°C. Similarly, stability study showed that the enzyme was fairly stable (> 80%) from 30-50°C. At 60°C the enzyme exhibited 44% of residual activity and becomes nil at 90°C due to thermal denaturation. The enzyme was considered to be thermostable compared to other lipases derived from R. oryzae, R. chinenesis, Yarrowia lipolytica and Mucor sp. [12,13,25-27]. The optimum temperature of the enzyme was found to be more than that of most of Aspergillus lipases [7,28]. A newly isolated Rhizopus strain also shows same optimum temperature, but beyond 50°C its activity falls sharply [24]. Thus, the present enzyme could be efficiently used for processes requiring stability at moderately high temperatures.
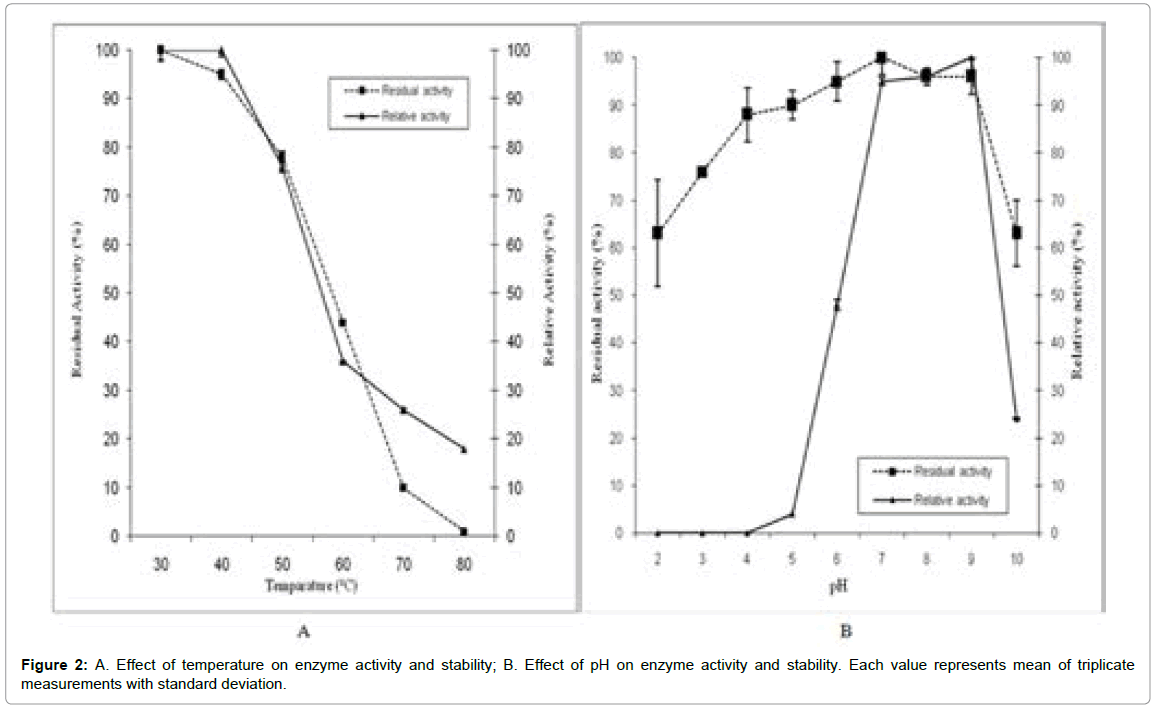
Figure 2: A. Effect of temperature on enzyme activity and stability; B. Effect of pH on enzyme activity and stability. Each value represents mean of triplicate measurements with standard deviation.
Evaluation of pH on activity and stability of lipase
Another remarkable aspect of this enzyme was its stability through wide range of pH. Figure 2B reveals the stability and retention of activity (>80%) between pH 4-9. Though the optimum pH of the enzyme was found to be 9 (100%), the stability was found to be maximum at pH 7 (100%). Compared to most of the Rhizopus lipases, whose optimum pH lie in the range 7- 8.5 [20,21,24], the present lipase was found to be most active in alkaline pH. Its optimum pH was equal to that of Pseudomonas fluorescens JCM5963, Pseudomonas sp. strain S5 [29] and comparable to Geotrichum sp. SYBC WU-3 [30] whose pH is 9.5. Although, Sayari et al. [31] reported a bacterial lipase fairly stable in the pH range of 4-9, yet for fungal system similar stability is sparsely observed. The wide pH stability of the present lipase proposes its applicability in range of industrial processes.
Half life of lipase at pH 7, 30°C and 50°C
Half life study of enzymes is carried out to estimate their stability and thus infer their usefulness as biocatalyst in industrial processes. As in the effect of temperature studies, the enzyme was found to retain its 100% activity at 30°C, pH 7 and about 80% of its activity at 50°C, its half life has been studied for these two temperatures and pH. The half life of lipase at 50°C was found to be 36 h. An interesting observation has also been recorded that the same enzyme can retain 77% of its activity till 2 h of incubation (Figure 3A) In case of 30°C enzyme was found to be more stable and the activity reached nearly 52% at the end of 30 days incubation (Figure 3B) In a similar study by Yao et al. lipase derived from Burkholderia ambifaria YCJ01 also showed high stability at 55°C and 30°C [16]. The observations in the current study was quite impressive and much better than half life of lipase from Burkholderia multivorans V2, which at 50°C and 60°C was found to be one hour and 10 min, respectively [32].
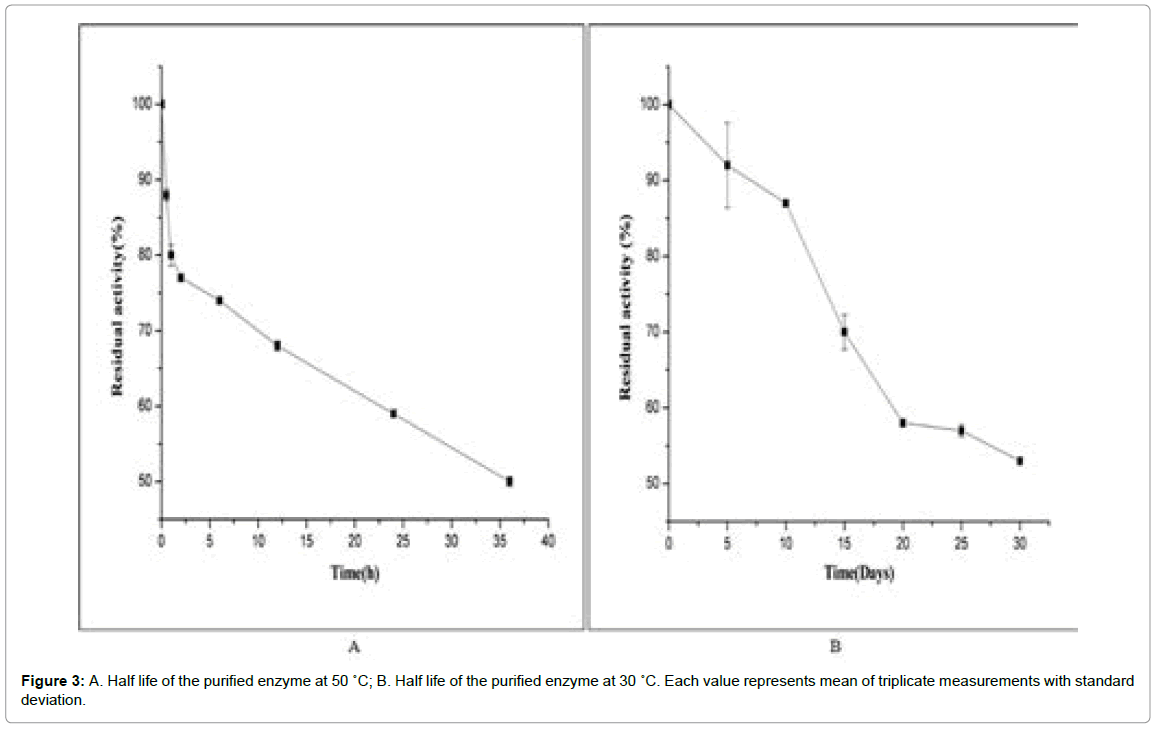
Figure 3: A. Half life of the purified enzyme at 50 ˚C; B. Half life of the purified enzyme at 30 ˚C. Each value represents mean of triplicate measurements with standard deviation.
Determination of substrate specificity
Substrate specificity of lipolytic enzymes helps to determine it application in desired sector. Lipases show preference for long chain acylglycerols (C8-18) and eterases hydrolyse short chain fatty acids (< C8) [23]. In the present study highest activity was shown towards pNPP (C16) followed by pNPS (C18) by the enzyme (Figure 4). Variation in the substrate specificity has been depicted by the lipases derived from Rhizopus sp. R. homothallicus [33] and R. oryzae [12] reported to have highest activity on C8 chain length substrates viz trioctanoin, whereas high hydrolytic activity against methyl laurate and palmitate has been reported in R. chinensis lipase [22].
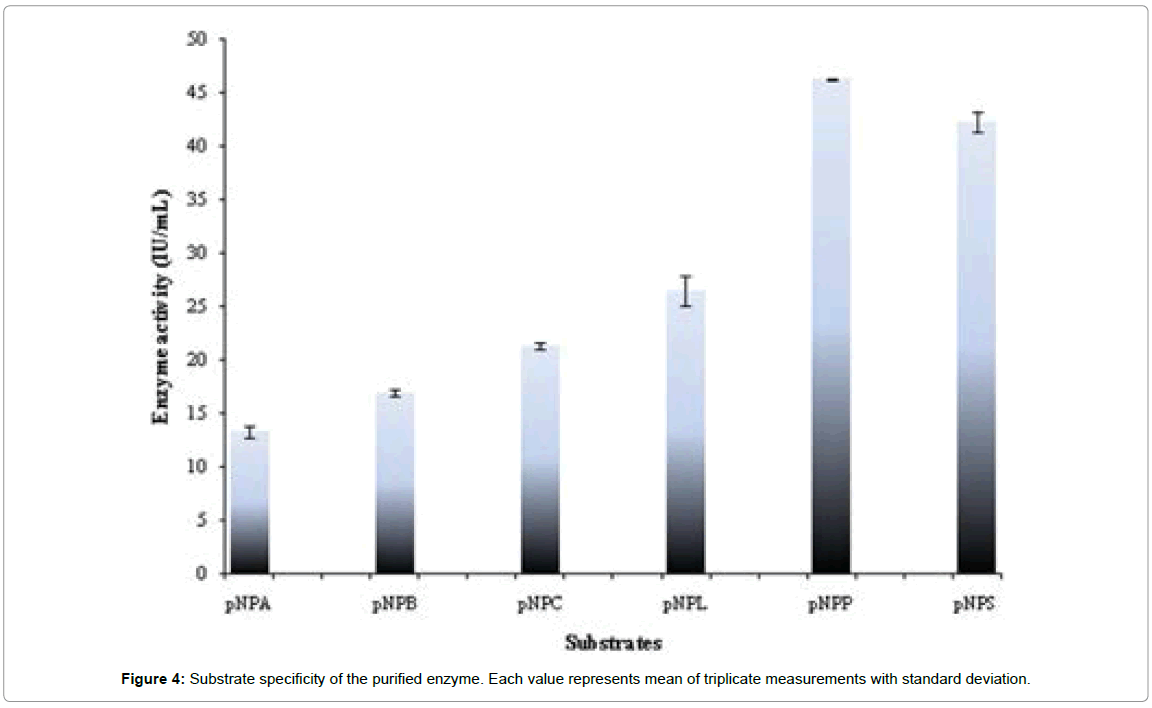
Figure 4: Substrate specificity of the purified enzyme. Each value represents mean of triplicate measurements with standard deviation.
Effect of metal ions on stability of lipase
The details of the effect of metal ions on the enzyme stability have been given in Table 2 Ca2+ was found to increase the activity of enzyme at the given concentration. The enhancement in activity could be attributed to complex formation between Ca2+ and fatty acids, changing the solubility of fatty acids and its behaviour at interface. Hiol et al. [27], Sayari et al. [31] and Sarkar et al. [34] reported similar effect of Ca2+ on the lipase activity of Mucor sp. and Staphylococcus sp., repectively. Yarrowia lipolytica lipase also showed enhancement in activity in presence of Ca2+ and inhibition by Hg2+ [35]. Na+ has shown no effect on the activity while, Zn2+ and Hg2+ both decreased the activity of the enzyme drastically. Others reduced the activity about 20%. Reduction in enzyme activity by Hg2+ suggests the involvement of a putative cysteine residue in the activity of enzyme [34]. Inhibition of enzyme by transition metals has been attributed to the interaction of ions with charged side chain groups of surface amino acids thus effecting overall enzyme structure [32].
| Metal ions (1mM) | Residual activity (%)a |
|---|---|
| Control | 100 ± 1.45 |
| Na+ | 100 ± 0.18 |
| K+ | 77 ± 0.28 |
| Ca2+ | 108 ± 1.28 |
| Mg2+ | 76 ± 1.16 |
| Cu2+ | 80 ± 0.8 |
| Zn2+ | 64 ± 0.83 |
| Hg2+ | 38 ± 0.62 |
| Co2+ | 82 ± 0.59 |
| Pb2+ | 84 ± 0.28 |
a100% residual activity of control enzyme corresponds to 16.78 IU/ml of enzyme activity. Each value represents mean ± standard deviation (n=3).
Table 2: Effect of metal ions on lipase stability.
Effect of surfactants and inhibitors on stability of lipase
Industrial application requires robust lipases which can retain their activity in presence of various detergents and contaminants. In order to probe in to this aspect, its residual activity in presence of various detergents and standard inhibitors has been measured. Table 3 gives the details of the effect of different surfactants. Tween 80 and Tween 20 slightly increased the enzyme activity post one hour incubation. Surfactants stimulate enzyme activity by probable change in conformation of enzyme thus increasing substrate accessibility [32]. Lipases from R. homothallicus and B. ambifaria YCJ01 showed similar trend and were activated by Tween 80 and inhibited by SDS [16,33]. The enzyme was not much affected by the other surfactants and retained more than 90% of its activity in their presence except CTAB. On the other hand, DTT drastically reduced the enzyme activity which infers involvement of cystein or thiol group in maintaining active enzyme structure, while, EDTA did not affect the enzyme activity. It shows the enzyme to be independent of any metals, hence not a metalloprotein [14].
| Residual activity (%)a | ||
| Control | 100 ± 0.95 | |
| Surfactants(1mM) | SDS | 99 ± 0.45 |
| CTAB | 40 ± 0.9 | |
| Triton-X-100 | 90 ± 0.82 | |
| Tween-80 | 106 ± 0.59 | |
| Tween-20 | 101 ± 0.84 | |
| Inhibitors (1mM) | EDTA | 97.01 ± 8.11 |
| Urea | 44.78 ± 4.4 | |
| DTT | 37.31 ± 11.0 | |
| Guanidium HCl | 54.73 ± 16.2 |
a100% residual activity of control enzyme corresponds to 16.78 IU/ml of enzyme activity. Each value represents mean ± standard deviation (n=3).
Table 3: Effect of surfactants and inhibitors on enzyme stability.
Effect of solvents on stability of lipase
Industrial processes like transesterification reactions require presence of solvents, good tolerance to them is the perquisite for any enzyme to be employed for biocatalysis of such processes. In contrast to R. oryzae lipase reported by Hiol et al. [12], which showed high stability towards long chain alkenes and alcohols the present enzyme showed stability in presence of polar solvents, short chain alcohols than non-polar ones (Table 4). It was found to retain about 100% of its activity in methanol and more than 90% activity in ethanol, propanol and acetone. In hexane and diethyl ether the enzyme was found to be more than 65% active. Methanol and ethanol stability was also reported by Kantak and Prabhune [24] and Yoo et al. [15] in Rhizopus sp. and Ralstonia sp., respectively. The purified lipase showed stability against most of the solvent with high resistance to methanol and ethanol indicating its suitable application in biodiesel production.
| Solvents (10% V/V) | Residual activity (%)a |
|---|---|
| Control | 100 ± 1.25 |
| Methanol | 99 ± 0.42 |
| Ethanol | 92 ± 1.26 |
| Propanol | 91 ± 0.58 |
| Isopropanol | 86 ± 1.37 |
| Butanol | 65 ± 2.5 |
| Hexane | 65 ± 0.2 |
| DMSO | 83 ± 0.86 |
| Diethyl ether | 69 ± 0.22 |
| Acetone | 91 ± 1.76 |
a100% residual activity of control enzyme corresponds to 16.78 IU/ml of enzyme activity. Each value represents mean ± standard deviation (n=3).
Table 4: Effect of different solvents on enzyme stability.
In the present work a simple, two step purification process was developed which gave both high fold purification (37 fold) and yield (51.36%) of lipase from R. oryzae NRRL 3562. The results are better than so far reported two step purification strategies which either gave high yield or fold purification but never both together. The enzyme also found to be smallest, only of 14.45 kDa, lipase till date reported from any fungus and especially of Rhizopus sp. Along with it, the enzyme also showed to be most active at alkaline pH (9) so far not seen in Rhizopus sp. and extremely resistant to changes in wide range of pH and moderately thermostable, with a month long half life at pH 7, 30°C. It also exhibited other industrially important characteristics such as stability in presence of most of metals, surfactants and solvents especially polar solvents, thus proving to be a robust enzyme suitable for varied applications including transesterification, detergent formulation etc.
The authors gratefully acknowledge CSIR, New Delhi for the financial support provided to Ms. Sunita Adak (CSIR-SRF) for the grant of her fellowship.