Journal of Agricultural Science and Food Research
Open Access
ISSN: 2593-9173
ISSN: 2593-9173
Research Article - (2018) Volume 9, Issue 1
Seven fungal agents were isolated from native Lycium arabicum plants growing in the Tunisian Centre-East and were screened for their ability to inhibit Fusarium oxysporum f. sp. radicis lycopersici (FORL) development and to promote plant growth. They were shown able to colonize tomato roots, crowns and stems. Seedlings inoculated or not with FORL and treated with these fungal isolates showed significant increments in all studied growth parameters (root length, shoot height, and fresh weight of roots and shoots). Tested as conidial suspensions or cell-free culture filtrates, I15 and I18 isolates were found to be the most active leading to 85.7-87.5% decrease in leaf and root damage index and to 93.6-98.4% lowered vascular browning extent over FORL-inoculated and untreated control. These two bioactive and growth-promoting isolates (I15 and I18) were microscopically and macroscopically characterized and identified using rDNA sequencing gene as being Alternaria alternata (MF693801) and Fusarium fujikuroi (MF693802). FORL mycelial growth was inhibited by 36.4-77.4 and 62.2-81% using their conidial suspensions and cell-free culture filtrates, respectively. Both isolates showed chitinolytic, proteolytic and amylase activities but lipolytic activity was displayed by F. fujikuroi (MF693802) only. To the best of our Knowledge, this is the first study that shows the potential use of fungi naturally associated to L. arabicum for suppression of Fusarium Crown and Root Rot and enhancement of tomato growth.
<Keywords: Antifungal activity; Associated fungi; Fusarium oxysporum f. sp. radicis-lycopersici; Tomato growth
Fusarium Crown and Root Rot (FCRR) is serious soilborne disease of tomato worldwide [1].Thecausal agent, Fusarium oxysporum f. sp. radicis-lycopersici (FORL) [2], is a long-lasting pathogen in soil due to its airborne dissemination to neighboring plants and to its survival as resting structures i.e., chlamydospores [1]. The lack of specific chemical fungicides and resistant cultivars led to increased interest to search for more innovative and effective alternatives. Biological control is considered as a key alternative for sustainable agriculture. Different fungal agents were shown to be efficient in controlling FORL such as nonpathogenicFusarium oxysporum [3], Trichodermaharzianum [4,5], binucleateRhizoctoniasolani [6], and F.equiseti [7]. A significant decrease by 50-73% in FORL radial growth and to 5.5% in disease incidence were achieved using some biofungicides based T. harzianun strain T22 [8].
Endophytic fungi naturally associated to wild or grown plants were also widely explored as promising antagonists and environmentally friendly tools for biocontrol of various plant diseases [9-11].Once established within tissues, fungal endophytes could protect their plant hosts from biotic and abiotic stresses [12]. These effects may be accomplished through the activation of their defense mechanisms [13] or by the inactivation of target pathogens and consequently disease development and severity[14]. In fact, various bioactive secondary metabolites were involved in these effects such as auxins [15], indole and its derivatives, and others [16].
Some endophytic fungi were shown effective in protecting tomato crops from fungal infections [17]. In fact, endophytic F. oxysporum strain Fo47, applied as root treatment, had significantly controlled Fusarium wilt of tomato caused by F. oxysporum f. sp. lycopersici [18]. Moreover, Qui et al. [19] noted 20% decrease in Verticillium wilt on tomato plants colonized by Piriformosporaindica. Rodriguez et al. [20] showed that the ability of F. oxysporum to inhibit Sclerotiniasclerotiorum growth in infected tomato plants through the release of the antifungal metabolite cyclosporine. Interestingly, F. equiseti GF191 had also successfully controlled FCRR disease through the secretion of antifungal compounds [21]. Also, endophytic F. solani had significantly limited root infection by FORL and subsequent disease development [13].
Moreover, fungal endophytes are well known for their contribution in plant fitness by promoting its growth and by enhancing its tolerance to environmental stresses [22,23] and its nutrient uptake and translocation. These effects may be achieved by the release of phytohormones [24], abscisic acid [25] and plant-growth regulatory compounds [26]. For instance, Johnson et al. [24] showed that endophytic Piriformosporaindica was able to improve plant growth not only by the improvement of nutrient uptake and translocation but also through the modulation of phytohormones as demonstrated on Nicotiana tabacum, Arabidopsis thaliana, and Hordeum vulgare test plants. Other fungal endophytes such as F. fujikuroi, SphacelomamanihoticolaCladosporium sp., Penicillium sp., and Aspergillus fumigatus had also enhanced tomato growth by releasing hormones [27-29]. Growth promotion may be also achieved through the enhancement of nitrogen fixation by producing nitrogenase as demonstrated for Scolecobasidiumhumicolas involved in [30].
Some previous studies demonstrated that wild Solanaceae plants are valuable sources of isolation of beneficial bioagents and extraction of bioactive compounds [31,32]. In fact, Veira et al. [33] showed the diversity of fungal agents isolated from Solanum cernuum. Vell and the strong antifungal potential displayed by Glomerellaacutata, Leptosphaeria sp., and Phomaglomerata. This is also the case of Aspergillusustusassociated with S. tuberosum which conferred induced resistance to Arabidopsis thaliana against various pathogens and its promoted growth [34]. Chaetomium spp. isolated from S. tuberosum plants was also shown able efficiently control late blight disease caused by the fungus-like pathogen Phytophthora infestans[35].
LyciumarabicumSchweinf. ex Boiss. is a wild Solanaceous species which is widely distributed mainly in African and Asian countries as well as in the Mediterranean region [36]. This species is known to contain alkaloids, steroids, saponins, coumarins and glycoalkaloids [37] but its exploration for isolation of potent biocontrol agents was not previously investigated. Thus, this study was performed to evaluate the capacity of conidial suspensions and cell-free culture filtrates of endophytic fungi recovered from L. arabicum to control Fusarium Crown and Root Rot disease, to improve tomato growth and to suppress FORL in vitro growth.
Pathogen isolation and culture
FORL isolate used in the bioassays was originally recovered from tomato plants showing typical Fusarium Crown and Root Rot (FCRR) disease symptoms i.e., plant wilting, vascular discoloration, and serious crown and root rots. Pathogen isolate used belonged to the Fungi collection of the Phytopathology Laboratory of the Regional Center of Research on Horticulture and Organic Agriculture at Chott-Mariem, Sousse, Tunisia.
Before being used for biological activities, FORL isolate was grown at 25°C for 5 days on Potato Dextrose Agar (PDA) medium amended with streptomycin sulphate (300 mg/L).
For mass-production of pathogenic inoculum, a mycelial plug (5 mm in diameter) of FORL, removed from 5-day-old colonies, was inoculated in 150 mL of Potato Dextrose Broth (PDB) medium injected in 500 mL Erlenmeyer flasks and incubated for 5-7 days under continuous shaking at 150 rpm in a rotating incubator. The obtained liquid culture was filtered through sterile Whatman No. 1 filter paper to remove mycelium and the collected conidial suspension was adjusted to 107 conidia/mL using a hemocytometer [8].
Tomato seedling preparation and growth conditions
Tomato cv. Rio Grande seeds were surface sterilized by immersion into 70% (v/v) ethanol for 2 min, then in 0.2% (v/v) sodium hypochlorite (NaOCl) for 3 min [38]. They were rinsed several times with sterile distilled water (SDW) and sown in alveolus plates (7×7 cm) filled with sterilized peat TM (FloragardVertriebsGmbHfürgartenbau, Oldenburg). Seedlings were grown the two-true-leaf growth stage under controlled conditions (24-26°C, 12 h photoperiod and 70% relative humidity) for about 28 days and watered regularly to avoid water stress.
Wild material processing and isolation of associated fungi
Fresh and healthy L. arabicum leaves, stems, fruits and flowers were collected from Tunisian littoral, Monastir (latitude 35°42'32.4"N, longitude E10°49'19.9") in November 2013.Collected samples were washed thoroughly under running tap water to remove any adhering soil particles.
Five leaf, stem, fruit and flower samples were surface disinfected according to Kjer et al. [39] method. They were immersed for 1 min in 70% (v/v) ethanol, then for 5 min in 10% (v/v) sodium hypochlorite, again for 30 s in a 70% (v/v) ethanol, and finally rinsed three times in SDW (3 min each). Sterility checks were performed for each sample to verify the efficiency of the disinfecting process. For these tests, 0.1 mL from the last rinse water was spread on solid PDA medium previously poured in Petri plates. Cultures were incubated for 6 days and observed regularly for the presence of any growing fungal colonies. Absence of such searched colonies is an indicator of the efficiency of the disinfecting process [40]. The surface-disinfected plant tissues were blotted dry on sterilized filter papers. They were transversely sectioned into pieces of 1 cm in length, using a sterile razor blade, placed in Petri plates containing PDA. Ten pieces were deposited in each plate and three plates were used per each organ sample. Plates were incubated at 25°C and examined daily for any fungal colonies growing from the incubated fragments. Once observed, growing fungal colonies were individually transferred to new PDA plates and incubated at 25°C. The collected fungal cultures were purified using the single-spore isolation technique and stored at 4°C or in 20% glycerol (v/v) at -20°C until future use. Morphological traits of purified colonies were noted, and spore-producing structures were examined microscopically to determine the taxonomic status of each isolate. Fungal isolates recovered from L. arabicum were affiliated to 10 distinct morphotypes. One isolate from each morphotype was selected for the screening of the endophytic colonization ability.
Preparation of conidial suspensions of associated fungi
Conidia of fungal isolates recovered from L. arabicum samples were harvested from actively growing colonies and suspended in 100 mL Potato Dextrose Broth (PDB) already poured in 250 mL Erlenmeyer flask. Cultures were incubated at 25°C for 12 days under continuous shaking at 150 rpm [41]. Liquid cultures were filtered through Whatman No. 1 filter paper to remove mycelium and the obtained conidial suspension was adjusted to 106conidia/mL using a Malassezhaemocytometer [42].
Preparation of cell-free culture filtrates of associated fungi
Fungal isolates were grown on PDA medium. An agar plug (6 mm in diameter) of each isolate was suspended in 100 mL PDB in a 250mL Erlenmeyer flask and incubated for 15 days at 28°C under continuous shaking at 150 rpm [43]. Fungal liquid cultures were filtered through Whatman No. 1. filter paper and the obtained filtrates were centrifuged thrice for 10 min at 10,000 rpm. Filtrates were further sterilized by filtration through a 0.22 μm pore size filter [44] before being used.
Test of the endophytic colonization ability
Fungal isolates recovered from L. arabicum were evaluated for their endophytic behavior and ability to colonize the internal tomato tissues. A group of five tomato cv. Rio Grande seedlings (at two-true-leaf stage) were dipped for 30 min into 25 mL of the conidial suspension (106 conidia/mL) of each tested isolate [45]. Control seedlings were soaked in an equal volume of SDW.Tomato seedlings were transferred to individual pots (12.5×14.5 cm) filled with commercialized peat and grown for 60 days at 20-25°C, with 70-85% relative humidity and a 12 h photoperiod. To check their ability to colonize tomato tissues, tested fungal isolates were recovered from tomato roots, crowns and stems according to Hallmann et al. [46] procedure. Plates were maintained at 25°C and examined daily for any growing fungal colonies. Colonies exhibiting similar morphological traits as the wild-type ones were selected and isolates originally used for seedling soaking were considered as endophytes.The colonization frequency (CF) was calculated according to Kumareson and Suryanarayanan [47] formula as follows:
CF (in %)=(Number of segments colonized by the test fungus/total number of segments plated)×100.
The percent of fungal colonization per target organ was arsine transformed before performing statistical analysis.
Assessment of FCRR suppression ability displayed by selected isolates
Fungal colonies exhibiting macro-morphological diversity and re-isolated onto PDA medium with a frequency exceeding 20% were picked separately onto PDA. Conidial suspensions and cell-free culture filtrates of eight selected fungal isolates were screened for their ability to suppress Fusarium Crown and Root Rot disease on tomato cv. Rio Grande plants under greenhouse conditions.
Tomato cv. Rio Grande seedlings were transplanted into individual pots (12.5×14.5 cm) containing commercialized peat. The tested biological treatment was applied to seedlings as culture substrate drench with 20 mL of a conidial suspension (106conidia/mL) or a cell-free supernatant prepared as described above. Inoculation was performed, one-week post-treatment, as substrate drench with 20 mL of FORL conidial suspension (107conidia/mL) [21]. Pathogen-free control (negative control or NC) seedlings were watered with SDW only. Pathogen-inoculated control (positive control IC) plants were challenged with a same volume of FORL conidial suspension and watered with SDW. All plants were cultured in a greenhouse at 20-25°C, 70-85% relative humidity and 12 h photoperiod. Five replicates of one seedling each were used for each individual treatment. The whole experiment was repeated two times. At 60 days post-inoculation with FORL (DPI), all plants were uprooted and peat adhering to roots was carefully washed off under running tap water. The parameters noted were disease severity, root length, shoot height, roots and shoot fresh weights and FORL re-isolation frequency (percentage of pathogen isolation from roots, collars and stems) on PDA medium.
FCRR severity was evaluated based on the above- and below-ground damage and on the vascular browning extent (from collar). Disease damage was assessed based on a 0-3 rating scale [48] where:0=no symptoms; 1=light yellowing of leaves, light or moderate rot on taproot and secondary roots and crown rot; 2=moderate or severe yellowing of leaves with or without wilting, stunting, severe rot on taproot and secondary roots, crown rot with or without hypocotyls rot and vascular discoloration in the stem; and 3=dead seedlings.
The frequency of FORL re-isolation from roots, collars and stems was calculated using following formula:
IR (%)=(r/R) × 100
Where r=Number of fragments showing pathogen growing colonies and R=total number of fragments plated on PDA medium.
Assessment of growth-promoting ability displayed by endophytic isolates
Eight selected endophytic fungal isolates were screened in vivo for their ability to improve tomato growth using their conidial suspensions or their cell-free culture filtrates.
Biological treatments were performed by dipping roots of a group of five tomato cv. Rio Grande seedlings (at two-true-leaf growth stage) for 30 min into fungal conidial suspensions and another group into cell-free filtrates [45]. Seedlings were transferred to individual pots (12.5×14.5 cm) containing commercialized peat. Control seedlings were similarly challenged using SDW. All seedlings (treated and controls) were grown under greenhouse conditions and regularly watered with tap water to avoid water stress. Five replicates of one seedling each were used for each individual treatment and the whole experiment was repeated twice. At 60 days post-treatment, parameters noted were root length, shoot height and fresh weight of roots and shoots.
Assessment of the in vitro antifungal activity of selected isolates
Eight endophytic isolates were evaluated for their capacity to inhibit the in vitro growth of FORL using the dual culture technique. Two agar plugs (6 mm in diameter) one colonized by the pathogen (removed from a 5-day-old culture at 25°C) and a second by the test fungus (removed from a 7-day-old culture at 25°C) were deposed equidistantly 2 cm apart on PDA medium supplemented with streptomycin sulfate (300 mg/L) [49]. Three replicates of one plate each were considered for each individual treatment and the whole experiment was repeated twice. Control plates were inoculated each by only one FORL plug. Cultures were incubated at 25°C for 5-6 days. Mean diameter (cm) of FORL colony was recorded when pathogen growth reached the center of control plates. Pathogen growth was estimated by averaging the two perpendicular diameters of each growing colony. Growth inhibition percentage was calculated according to the following formula [50]:
Growth inhibition (%)=[(dc – dt)/ dc]×100
Where dc=Mean colony diameter in control plates; dt=Mean colony diameter in treated plates.
Assessment of the in vitro antifungal activity of fungal cell-free culture filtrates
Five Fungal isolates were chosen based on their ability to suppress Fusarium Crown and Root Rot disease severity by more than 50% over control and to reduce FORL mycelial growth by more than 60%.
The selected isolates were grown on PDA medium at 25°C in the dark for 5 days [51]. A mycelial agar plug (5 mm in diameter) removed from the edge of colony of each endophytic fungus was inoculated individually in 100 mL of PDB poured in 250-mL Erlenmeyer flasks. Cultures were incubated under continuous shaking at 150 rpm at 25°C for 30 days [41].
A 2 mL-sample of each tested culture filtrate was centrifuged thrice at 10,000 rpm for 10 min. Collected supernatant fluids were sterilized by filtration through a 0.22 μm pore size filter. Control treatment was the PDB filtrate. Filtrates were aseptically added at the concentration of 10% (v/v) to Petri plates containing molten PDA medium amended with streptomycin sulfate (300 mg/mL) (w/v). After medium solidification, three 6 mm agar plugs colonized by FORL were placed equidistantly in each Petri plate. Three replicate plates for each tested treatment were used and all the experiment was repeated twice. Cultures were incubated at 25°C for 5 days. The diameter of pathogen colony (in treated and control plates) was measured and the pathogen growth inhibition rate was calculated as described above.
Identification of the most bioactive fungal isolates by ITS sequencing gene
The genomic DNA extraction of the four selected fungal isolates was performed using the DNA Mini Kit (Analytik Jena, Biometra) according to manufacturer instructions. For each test fungus, the ITS region, the widely used for general fungal identification [52], was amplified by polymerase chain reaction (PCR) using both universal fungal primers: ITS1 (TCCGTAGGTGAACCTGCGG) and ITS4 (TCCTCCGCTTATTGATATGC). The PCR reaction was performed in a total reaction volume of 25 μL containing 5 μL of buffer (5×), 2.5 μL of dNTP (2 mM), 1.5 μL of MgCl2(25 mM), 0.25 μLTaq polymerase (5 U/μL), 2.5 μL of each primer (6 μM), 5.75 μL of ultra-pure water and 5 μL of genomic DNA templates (10 ng).
The amplification program, performed in an OpticonII (Biorad) Thermal Cycle, included an initial denaturation at 94°C for 5 min, followed by 30 cycles of denaturation at 94°C for 1 min, annealing at 50°C for 1 min and extension at 72°C for 1 min. Amplification was terminated by a final extension step of 7 min at 72°C. The obtained PCR products were electrophoresed in agarose gel 1% (w/v) stained with ethidium bromide, and visualized under UV light. gene sequencing was carried out in a private laboratory (Biotools, Tunisia). ITS sequences were analyzed with Basic Local Alignment Search Tool (BLAST) through GenBank (http://www. blast.ncbi.nlm. nih.gov/).
Enzymatic activity displayed by the selected bioactive isolates
The most effective fungi (I15 and I18 isolates) in suppressing FCRR disease were screened for their ability to produce extracellular enzymes (namely amylases, lipases, proteases, and chitinases) using qualitative techniques as described below. All assays were carried out in triplicates.
Amylase activity
Amylase activity was tested by growing fungal isolates (plugs of 3 mm in diameter) on Glucose Yeast Extract Peptone Agar (GYEP) medium amended with 0.2 g starch. After incubation at 25°C for 4 days, plates were flooded with 1% iodine in 2% potassium iodide and the formation of white zones around colonies, induced by the digestion of starch added to medium, indicated a positive reaction [53].
Lipolytic activity
For lipase activity, fungal isolates (plugs of 3 mm in diameter) were grown on Peptone Agar (PA) medium amended with sterilized tween 20 diluted at 1% v/v. Plates were incubated at 25°C for 3-7 days. The presence of a visible precipitate around the colony, due to the formation of calcium salts of the lauric acid released by the enzyme, indicated a positive lipase activity [53].
Proteolytic activity
For protease activity, fungal agar plugs (3 mm in diameter) removed from 10-day-old cultures were spot inoculated on Casein Starch Agar with 1% skimmed milk and incubated at 25°C for 96 h. After incubation, the formation of clear halos around fungal colonies indicated a positive proteolytic activity [53].
Chitinolytic activity
Chitinase activity was tested by inoculating fungal plugs (3 mm in diameter) on chitin-based medium. Cultures were maintained at 25 ± 2°C for 10 days. Isolates displaying chitinolytic activity grew on the medium while no growth indicates absence of chitinase activity [54].
Statistical analysis
Data were subjected to one-way analysis of variance (ANOVA) using Statistical Package for the Social Sciences (SPSS) software for Windows version 20.0. Each experiment was repeated twice. As no significant interactions between treatment and experiments were noted, one representative trial of each experiment was presented in the current study. Data were analyzed according to a completely randomized design. Means were separated using LSD or Duncan Multiple Range tests (at P ≤ 0.05). Correlations between FCRR severity and plant growth parameters were tested using bivariate Pearson’s test at P ≤ 0.05.
Endophytic fungal isolation frequency and diversity
Data given in Table 1 revealed that a total of 40 fungal isolates were recovered from L. arabicum leaves, stems, flowers, and fruits. There was a difference in the isolation frequency of isolates depending on plant parts explored. In fact, 13 isolates (32.5% of the total collected) were originated from leaves, 11 (27.5%) from stems, 9 (22.5%) from fruits, and 7 (17.5%) from flowers. Interestingly, a macroscopic variability was also noticed between the 40 fungal isolates. They were affiliated to 5 genera namely Fusarium, Alternaria, Penicillium, Aspergillus, and Trichoderma.
| Identification | Leaves | Stems | Flowers | Fruits | N Total (%) | F Total (%) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | F (%) | N | F (%) | N | F (%) | N | F (%) | |||
| Fusarium | 2 | 1.67 | 2 | 1.67 | 1 | 0.83 | 2 | 1.67 | 7 | 5.83 |
| Alternaria | 6 | 5 | 2 | 1.67 | 2 | 1.67 | 2 | 1.67 | 12 | 10 |
| Penicillium | 1 | 0.83 | 2 | 1.67 | 1 | 0.83 | 1 | 0.83 | 5 | 4.17 |
| Aspergillus flavus | 1 | 0.83 | 2 | 1.67 | 1 | 0.83 | 2 | 1.67 | 6 | 5 |
| Aspergillus niger | 1 | 0.83 | 1 | 0.83 | 1 | 0.83 | 1 | 0.83 | 4 | 3.33 |
| Trichoderma | 2 | 1.67 | 2 | 1.67 | 1 | 0.83 | 1 | 0.83 | 6 | 5 |
| N Total | 13 | - | 11 | - | 7 | - | 9 | - | 40 | - |
| F Total | - | 32.5 | - | 27.5 | - | 17.5 | - | 22.5 | - | 100 |
Table 1: Fungal isolates from Lycium arabicum leaves, stems, flowers and fruits on Potato Dextrose Agar medium and their relative isolation frequency. N: Number of isolates; F: Isolation frequency (%) based on their macro- and micro-morphological traits. It should be highlighted that Alternaria was the mostly isolated genus (10%). The isolation frequency of Aspergillus , Fusarium , Trichoderma and Penicillium were estimated at 8.33, 5.83, 5 and 4.17%, respectively (Table 1).
Endophytic colonization ability
Based on the colony characteristics and morphology, the 40 fungal isolates recovered from L. arabicum plants were classified into 10 different morphotypes. One isolate from each morphotype was selected for endophytic colonization screening. Results revealed that all treated plants remained healthy until the end of the experiment. The ten isolates tested were found to be non-pathogenic and were selected for further screenings.
ANOVA analysis revealed that tomato colonization frequency, noted 60 days post-treatment, depended significantly (at P ≤ 0.05) upon fungal treatments tested. Data shown in Table 2 showed that colonization frequency ranged between 16.6 and 83.3% from roots, between 16.6 and 76.6% from crowns, and between 6.6 and 73.3% from stems. The highest colonization frequencies from roots, crowns and stems (83.3, 76.6 and 73.3%, respectively) were noted on plants treated with I15 and I18 isolates. I11, I14, I16, I19 and I20 isolates exhibited significantly similar tomato colonizing ability where their respective colonization frequencies were estimated at 60-66.6, 46.6-60, and 36.6-53.3%, from roots, crowns and stems. The lowest colonization ability, ranging between 6.6 and 16.6%, was noted on plants treated with I12, I13 and I17 isolates.
| Isolate | Isolation origin | Roots | Crowns | Stems |
|---|---|---|---|---|
| NC | - | 0 d | 0 c | 0 d |
| I11 | Flowers | 66.67 b | 60.00 b | 53.33 b |
| I12 | Leaves | 16.67 c | 16.67 c | 6.67 d |
| I13 | Stems | 16.67 c | 16.67 c | 6.67 d |
| I14 | Stems | 66.67 b | 60.00 b | 53.33 b |
| I15 | Leaves | 83.33 a | 76.67 a | 73.33 a |
| I16 | Fruits | 60.00 b | 46.67 b | 36.67 b |
| I17 | Stems | 16.67 c | 16.67 c | 6.67 d |
| I18 | Stems | 83.33 a | 76.67 a | 73.33 a |
| I19 | Fruits | 66.67 b | 60.00 b | 53.33 b |
| I20 | Leaves | 66.67 b | 60.00 b | 53.33 b |
Table 2: Re-isolation frequency (%) of endophytic fungal isolates from tomato cv. Rio Grande roots, crowns and stems noted 60 days postinoculation. NC: Untreated control; I11: Isolate from flowers; I15, I20: Isolates from leaves; I18, I14: Isolates from stems; I16, I19: Isolates from fruits. Within each column, values followed by the same letter are not significantly different according to Duncan Multiple Range test (at P ≤ 0.05).
Fungal isolates inoculated to tomato seedlings, successfully re-isolated onto PDA medium with a frequency exceeding 20%, and showing similar traits as the wild type ones were classified as endophytes. Thus, 7 isolates out of the 10 tested and fulfilling the above-mentioned conditions (namely I11, I14, I15, I16, I18, I19 and I20) were selected for the in vivo screening of their antifungal potential against FORL and their plant growth-promoting ability.
Effect of endophytic isolates on disease severity
Disease-suppressive potential of conidial suspensions: ANOVA analysis revealed that FCRR severity, based on above- and below-ground damage and noted on tomato plants 60 days post-inoculation with FORL, varied significantly (at P ≤ 0.05) depending on biological treatments tested which were applied as conidial suspensions. Data given in Figure 1A (a) showed that the seven isolates tested had significantly decreased leaf and root damage index by 28.5 to 85.7% relative to pathogen-inoculated and untreated control (IC). I15- and I18-based treatments were found to be the most effective in suppressing FCRR severity by 85.7% on tomato plants challenged with FORL compared to control. Moreover, I11, I19 and I20 isolates exhibited significantly similar ability to decrease FCRR severity, by 57.1% as compared to FORL-inoculated and untreated control (IC) and by 40% relative to hymexazol-treated control (or FC).The lowest disease-suppressive potential (28.1%) was noted on plants treated with I14 and I16 conidial suspensions (Figure 1).
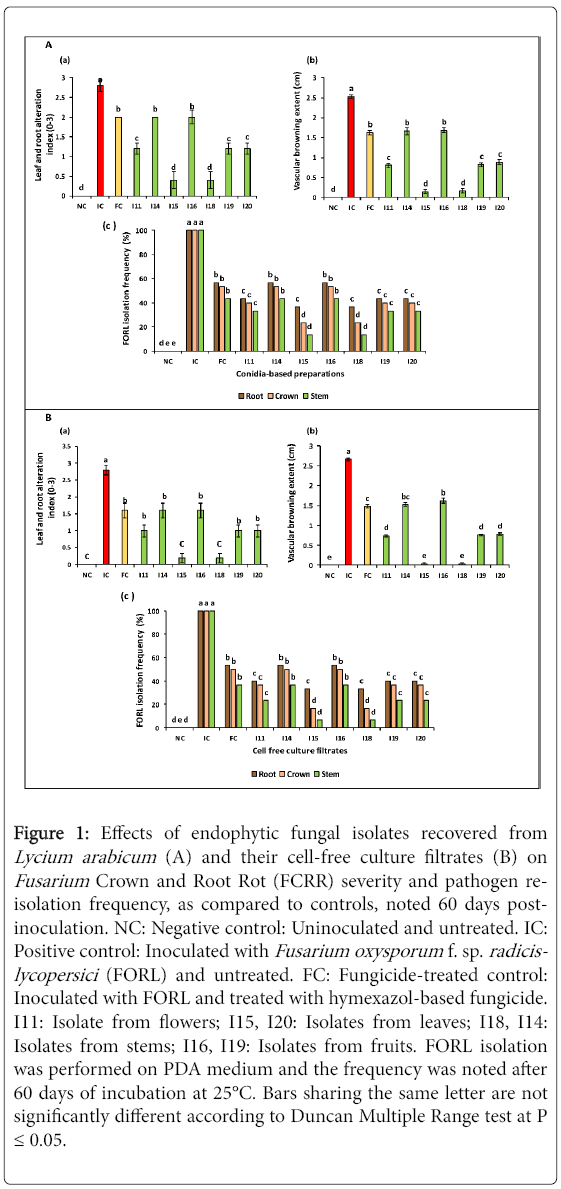
Figure 1: Effects of endophytic fungal isolates recovered from Lycium arabicum (A) and their cell-free culture filtrates (B) on Fusarium Crown and Root Rot (FCRR) severity and pathogen reisolation frequency, as compared to controls, noted 60 days postinoculation. NC: Negative control: Uninoculated and untreated. IC: Positive control: Inoculated with Fusarium oxysporum f. sp. radicislycopersici (FORL) and untreated. FC: Fungicide-treated control: Inoculated with FORL and treated with hymexazol-based fungicide. I11: Isolate from flowers; I15, I20: Isolates from leaves; I18, I14: Isolates from stems; I16, I19: Isolates from fruits. FORL isolation was performed on PDA medium and the frequency was noted after 60 days of incubation at 25°C. Bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P ≤ 0.05.
Also, as shown in Figure 1A (b), the vascular discoloration extent (measured from collar) was lowered by 33.3 to 94.4% compared to pathogen-infected control (IC) following treatments using conidial suspensions of tested isolates. Similarly, I15- and I18-based treatments were found to be the most efficient in suppressing the vascular discoloration extent by 93.6-94.4% versus control (IC). Also, interestingly, treatments with I11, I19 and I20 conidial suspensions had lowered this parameter by 65-68.2% relative to FORL-inoculated and untreated control (IC) and by 45.6-50.6% compared to hymexazol-treated control (or FC).
Re-isolation frequency of FORL onto PDA medium from roots, crowns and stems of treated tomato plants varied depending on tested biological treatments. Data given in Figure 1A (c) showed a reduction in FORL re-isolation frequency by 43.3-63.3, 46.6-76.6, and 66.6-86.6% from roots, crowns and stems, respectively, as compared to FORL-inoculated and untreated control where pathogen was recovered at 100%. It should be highlighted that the least pathogen re-isolation frequency was achieved using treatments with I15 and I18conidial suspensions.
Disease-suppressive potential of cell-free culture filtrates: The disease-suppressive potential of cell-free culture filtrates, noted 60 days post-inoculation with FORL, varied significantly (at P ≤ 0.05) depending on biological treatments tested. Results presented in Figure 1B (a) showed strong decrease in FCRR severity, as estimated based on leaf and root damage intensity, ranging between 42.8 and 92.8% compared to FORL-inoculated and untreated control (IC). Interestingly, cell-free culture filtrates of I15 and I18 were found to be the most efficient treatments by suppressing FCRR symptoms, by 92.8% relative to control (IC) and by 87.5% relative to pathogen-inoculated and hymexazol-treated control (FC). In addition, cell-free filtrates from I11, I14, I16, I19 and I20 isolates exhibited significantly similar ability in decreasing disease severity by 42.8-64.2% relative to FORL-inoculated and untreated control (IC).
Data shown in Figure 1B (b) revealed that FCRR severity, as measured by the vascular discoloration extent (from collar), was significantly (at P ≤ 0.05) reduced by 39-98.4% compared to FORL-inoculated and untreated control. Cell-free culture filtrates from I15 and I18 isolates were found to be the most effective in reducing this disease severity parameter by 98.4% over FORL-inoculated and untreated control (IC) and by 97.2% versus pathogen-inoculated and hymexazol-treated control (FC). Treatments with I11, I19 and I20 culture filtrates showed significantly similar potential in decreasing the vascular discoloration extent by 70-72.1% as compared to IC control and were found to be more efficient than hymexazol (FC).
Pathogen re-isolation frequency onto PDA medium from treated tomato plants also varied depending on tested cell-free filtrates. Figure 1B (c) showed 46-66.6, 50-83.3, and 63.3-93.3% decrease in FORL re-isolation frequency from tomato roots, crown and stems, respectively, compared to control (100%), following treatments with filtrates of tested isolates. It should be highlighted the least FORL re-isolation frequency (estimated at 6.6, 16.6 and 33.3%) was noted on stems, crowns and roots of plants treated with I15 and I18 cell -free filtrates.
Growth-stimulating effect of selected endophytic isolates on tomato plants inoculated with FORL
Stimulating effect of conidial suspensions: ANOVA analysis performed for all tomato growth parameters (root length, plant height, root and aerial part fresh weights), noted 60 days post-inoculation, revealed a significant variation (at P ≤ 0.05) depending on biological treatments tested. In fact, as shown in Figure 2A (a), an important increment in tomato root length, by 34.2-85.3% over FORL-inoculated and untreated control (IC), was recorded on treated tomato plants. Treatments with I15 and I18 conidial suspensions led to the highest increase (by 84-85.3% over control (IC)) in root length. Interestingly, I11- and I20-based treatments had also significantly improved this parameter by 65.4-66% over IC control and by 16.8-17.2% over hymexazol-treated control (FC).
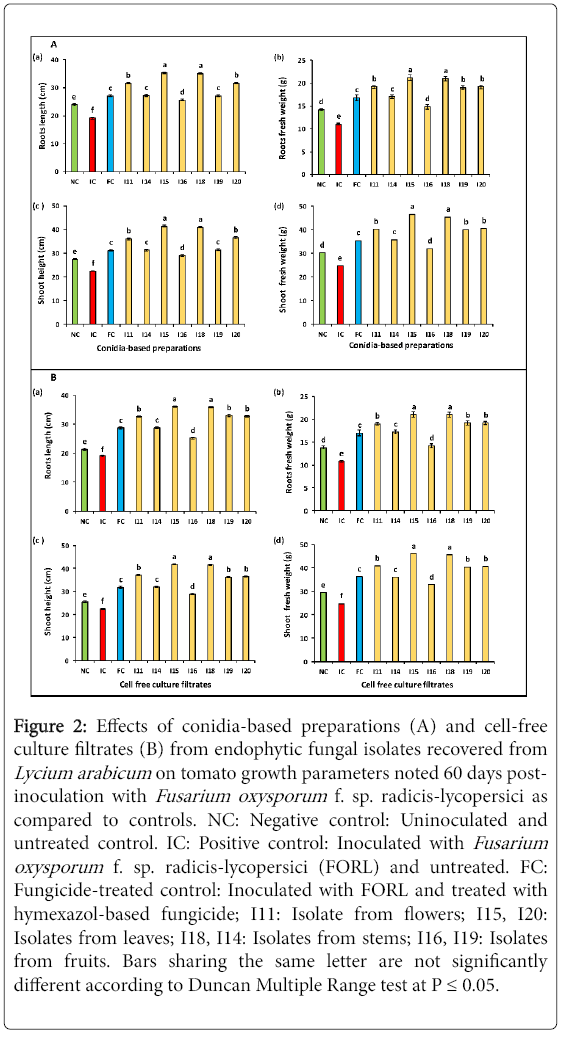
Figure 2: Effects of conidia-based preparations (A) and cell-free culture filtrates (B) from endophytic fungal isolates recovered from Lycium arabicum on tomato growth parameters noted 60 days postinoculation with Fusarium oxysporum f. sp. radicis-lycopersici as compared to controls. NC: Negative control: Uninoculated and untreated control. IC: Positive control: Inoculated with Fusarium oxysporum f. sp. radicis-lycopersici (FORL) and untreated. FC: Fungicide-treated control: Inoculated with FORL and treated with hymexazol-based fungicide; I11: Isolate from flowers; I15, I20: Isolates from leaves; I18, I14: Isolates from stems; I16, I19: Isolates from fruits. Bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P ≤ 0.05.
Results given in Figure 2A (b) showed a variable ability to increase the root fresh weight depending on biological treatments tested by 34.5-92.7% over positive control (IC) and by 25-26.1% over hymexazol-treated control (FC). The highest increment (by about 92.7%) was recorded on plants treated with I15 and I18 conidial suspensions. Treatments with I11, I19 and I20 conidial suspensions had significantly similar effect on the root fresh weight which was improved by 72.7-74.5% relative to infected control (IC) and by 13-14.2% versus hymexazol-treated control (FC).
Data graphed in Figure 2A (c) showed that all tested biological treatments had significantly improved shoot height by 34.7 to 90.3% versus FORL-inoculated and untreated control (IC) and by 29-83.9.7% over pathogen-free control (NC). The highest shoot height increments (82.2 and 83.9%) were recorded on plants treated with I15 and I18 conidial suspensions, respectively. Moreover, a significant increase in this parameter, by 60.4-62.6% compared to inoculated and untreated control (IC), and by 15.5-17.1% relative to hymexazol-treated control (FC) was achieved using I11 and I20 conidial suspensions.
Figure 2A (d) illustrated the significant (at P ≤ 0.05) increments in the shoot fresh weight noted using all tested biological treatments compared to FORL-inoculated control (IC) (29-87.9%) or to pathogen-free and untreated control plants (NC) (5.9-54.3%). The highest improvement of shoot fresh weight (83-87.9% higher than IC control) was recorded on plants treated with I15 and I18 conidial suspensions. Moreover, I11-,I19- and I20-based treatments had also enhanced this parameter by 61.2-63.7% over FORL-inoculated and untreated control (IC) and by 43.3-49% as compared to hymexazol-treated control (FC).
Stimulating effect of the cell-free culture filtrates: Growth parameters (root length, shoot height, roots and shoot fresh weights), noted on tomato seedlings 60 days post-inoculation with FORL, varied significantly depending on tested biological treatments.
All cell-free culture filtrates tested had significantly (at P ≤ 0.05) improved root length of FORL-inoculated and treated tomato seedlings by 31.6-88.6% compared to the untreated control (IC) and by 17.9-69% over pathogen-free ones (NC) (Figure 2B (a)). The highest increment in this parameter, by 87.7-88.6% over IC control and by 24.9-25.6%, relative to hymexazol-treated control (FC), was induced by I15 and I18 culture filtrates. Treatments based on I11, I19 and I20 cell-free filtrates had also increased this parameter by 70.5-72.7% compared to FORL-inoculated and untreated control (IC) and by 13.5-25.6% relative to FORL-inoculated and fungicide-treated control (FC).
Data given in Figure 2B (b) showed that all tested cell-free culture filtrates had significantly (at P ≤ 0.05) improved root fresh weight over controls. The recorded increments, compared to FORL-inoculated control, ranged between 31.4 and 94.4% and the highest one was recorded on tomato plants treated with I15 and I18 filtrates. Those from I11, I15 and I18 isolates had also enhanced this growth parameter by 75.9-77.7% over FORL-infected and untreated control (IC) and by 11.7-12.9% over fungicide-treated control (FC).
Results presented in Figure 2B (c) revealed that all tested filtrates had significantly (at P ≤ 0.05) stimulated by 28.9-86.1% the height of tomato shoots, as compared to FORL-inoculated and untreated control (IC) and by 13.4-64.2% versus pathogen-free control (NC). Cell-free filtrates from I15 and I18 isolates were found to be the most effective treatments leading to in 86.1% increase in shoot height. Moreover, an improvement by 61.1-65.2% was also achieved using I11, I15 and I18 filtrates as compared to pathogen inoculated and untreated control (IC) and by 13.8-16.7% over FORL-inoculated and fungicide-treated control (FC).
Data given in Figure 2B (d) showed that all cell-free filtrates tested had significantly (at P ≤ 0.05) increased shoot fresh weight by 32.2-85.4% compared to pathogen-inoculated and untreated control (IC) and by 11.5-56.4% versus pathogen-free control (NC). The highest improvement (83.8-85.4% higher than IC control) was noted following treatments with I15 and I18 cell-free filtrates. Those from I11, I15 and I18 isolates had also significantly enhanced this parameter by 61.1-65.2% compared to FORL-inoculated and untreated control (IC) and by 11.6-12.7% compared to FORL-inoculated and hymexazol-treated control (FC).
Correlation between FCRR severity and growth parameters
Treatments with conidial suspensions: Pearson’s correlation analysis performed for growth and disease severity parameters revealed that tomato root length and shoot height were significantly and negatively related to the leaf and root damage intensity (r=-0.7; P=0.024; r=-0.682; P=0.030) and to the vascular browning extent (r=-0.757; P=0.011; r=-0.742; P=0.014). Also, root and shoot fresh weights were significantly and negatively linked to the leaf and root damage scores (r=-0.654; P=0.040; r=-0.649; P=0.042) and to the vascular browning extent (r=-0.742; P=0.014; r=-0.721; P=0.019).
This correlation analysis indicated that the lowered Fusarium Crown and Root Rot severity, observed on tomato plants treated with endophytic conidial suspensions, was associated to the recorded decrease in FORL colonization ability leading, thus, to the noted plant growth stimulation.
Treatments with cell-free culture filtrates: Pearson’s correlation analysis performed for cell-free culture filtrate data showed that tomato root length and shoot height were significantly and negatively correlated to the leaf and root damage rate (r=-0.70; P=0.024; r=-0.682; P=0.030) and to the vascular browning extent (r=-0.757; P=0.011; r=-0.742; P=0.014). Root and shoot fresh weights were also significantly and negatively related to the leaf and root damage scores (r=-0.654; P=0.040; r=-0.649; P=0.042) and to the vascular browning extent (r=-0.730; P=0.017; r=-0.721; P=0.019).
These present correlation results indicated that the disease-suppressive effect achieved using fungal cell-free filtrates was also associated to the recorded decrease in FORL colonization ability leading consequently to the noted promotion of plant growth.
Growth-stimulating effect of selected endophytic isolates on pathogen-free tomato plants
The seven fungal isolates tested did not induce any disease symptoms when inoculated to tomato plants which remained healthy till 60 days post-inoculation. As they were found to be non-pathogenic, their conidial suspensions and their cell-free culture filtrates were further screened for their ability to stimulate growth of pathogen-free tomato plants.
Effect of fungal conidial suspensions: ANOVA analysis revealed that plant growth parameters (root length, root fresh weight, shoot height, and shoot fresh weight), noted 60 days post-treatments, varied significantly (at P ≤ 0.05) depending on tested biological treatments.
Data given in Figure 3A (a) revealed that all conidial suspensions tested had significantly enhanced tomato root length 44.2-76.7% over pathogen-free and untreated control (NC). The highest improvement, estimated at 73.7-76.7%, was achieved following treatments using I15 and I18 conidial suspensions. In addition, I11- I19- and I20-based treatments had also significantly improved root length by 58.9-59%. For the remaining fungal treatments, the recorded increments ranged between 42.4 and 44.7%.
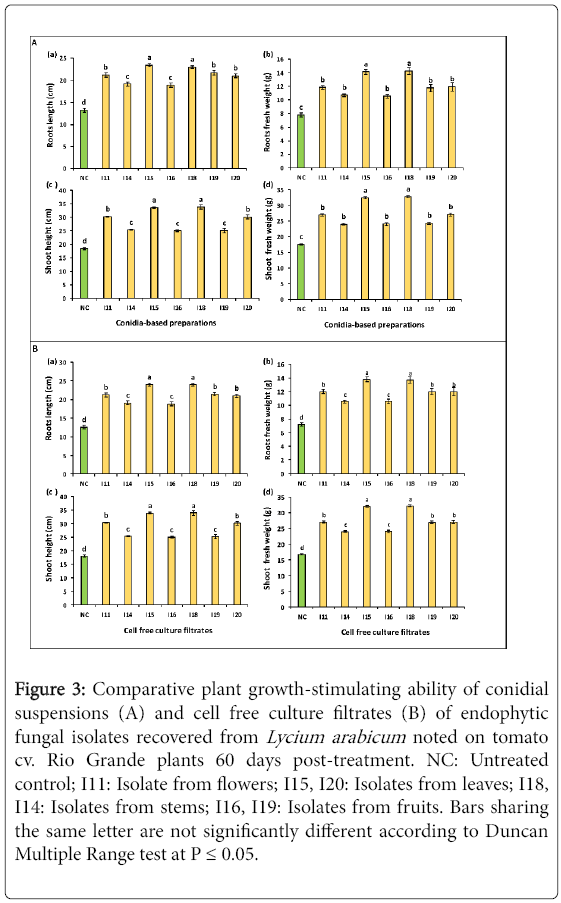
Figure 3: Comparative plant growth-stimulating ability of conidial suspensions (A) and cell free culture filtrates (B) of endophytic fungal isolates recovered from Lycium arabicum noted on tomato cv. Rio Grande plants 60 days post-treatment. NC: Untreated control; I11: Isolate from flowers; I15, I20: Isolates from leaves; I18, I14: Isolates from stems; I16, I19: Isolates from fruits. Bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P ≤ 0.05.
As measured based on root fresh weight, all conidial suspensions tested had significantly enhanced this parameter by 35.3-83% relative to control (NC). Treatments with I15 and I18 conidial suspensions led to the highest increase in this parameter (81.7-83%). The remaining treatments (I11, I14, I16, I19 and I20 suspensions) exhibited significantly similar effect where the recorded root fresh weight promotion was of about 35.2-53.3% relative to control (Figure 3A (b)).
Results graphed in Figure 3A (c) showed that all conidial suspensions tested had significantly enhanced shoot height by 36-83.7% as compared to pathogen-free and untreated control (NC). The greatest increase (82.6-83.7%) was achieved using I15 and I18 conidial suspensions. Furthermore, treatments with I11 and I20 conidial suspensions had significantly similar effect on this parameter where the recorded promotion varied between 63.3 and 64.2%. The least enhancement (36-37.6%) was noted on tomato plants treated with I14, I16 and I19 conidial suspensions.
Data given in Figure 3A (d) revealed that shoot weight increase achieved following biological treatments tested ranged between 36 and 86.4% as compared to untreated control (NC) where I15- and I18-based treatments were the most effective leading to 84-86% increase in this parameter. Furthermore, the remaining conidial suspensions (from I11, I14, I16, I19 and I20 isolates) had significantly improved shoot weight by 36-53.8% over control.
Effects of cell-free culture filtrates: ANOVA analysis performed for tomato growth parameters revealed a significant (at P ≤ 0.05) variation in the root length, root fresh weight, shoot height, and shoot fresh weight), noted 60 days post-treatments, depending on treatments tested.
As shown in Figure 3B (a), the root length was increased by 49.4-90.3% over control (NC) using all tested culture filtrates. The highest enhancement of root length, by 90-90.3% over pathogen-free control (NC), was noted on plants treated with I15 and I18 cell-free filtrates. Interestingly, I11, I19, and I20 filtrates were also effective in improving this parameter by about 66.7-69.7%. The least but relatively interesting increment, estimated at 49.4-51.8%, was induced by I14 and I16 filtrates.
As estimated based on their effects on the root fresh weight, all biological treatments tested had significantly (at P ≤ 0.05) enhanced this parameter by 46.6-91.3% over control. I15 and I18 filtrates induced the highest stimulating effect by increasing this parameter by 90.2-91.3% versus control. The least promoting effect, of about 46.6%, was noted on plants treated with I14 and I16 filtrates.
Results presented in Figure 3B (c) showed that all treatments tested had significantly (at P ≤ 0.05) increased shoot height by 39.5-88.7% relative to the untreated control (NC). The highest enhancement, by 88.5-88.7% versus control, was induced by I15 and I18 filtrates. Interestingly, I11 and I20 filtrates had also significantly improved this growth parameter by about 67.7-68.9%. The least growth-stimulating effect, estimated at 39.5-41.2%, was obtained following treatments with I14, I16 and I19 filtrates.
Data provided in Figure 3B (d) revealed that all culture filtrates tested had significantly (at P ≤ 0.05) enhanced shoot fresh weight by 43.2-91.6% in treated plants compared to the untreated control ones (NC). Treatments with I15 and I18 filtrates were found to be the most effective in enhancing this parameter by 80.7-85.3% over control. Furthermore, I11, I19 and I20 filtrates had also improved shoot growth by about 60-61.1%. The least growth-promoting effect, of about 43.2-43.4%, was recorded on plants treated with I14 and I16 cell-free culture filtrates.
In vitro antifungal potential of the selected endophytic isolates against FORL
Effect by direct confrontation: ANOVA analysis revealed a significant (at P ≤ 0.05) decrease in FORL colony diameter, noted after 5 days of incubation at 25°C,depending on biological treatments tested as compared to the untreated control. As shown in Figure 4 (a), the recorded reduction in FORL mycelial growth varied from 36.4 to 77.4% depending on treatments tested. The highest inhibition, of about 77.2-78.1% versus control, was noted on pathogen colonies co-cultured withI15 and I18 isolates (Figure 5A). Mycelial growth of pathogen co-cultured with I11, I19 and I20 isolates showed 60.5-61.5% less growth as compared to the control. The lowest decrease in this parameter (by 36.4-37% over control) was recorded on pathogen colonies co-cultured with I14 and I16 isolates.
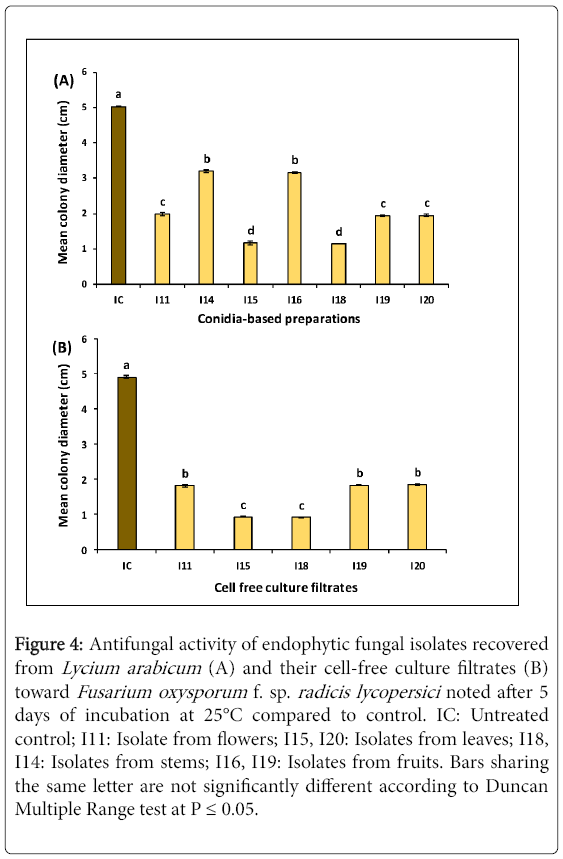
Figure 4: Antifungal activity of endophytic fungal isolates recovered from Lycium arabicum (A) and their cell-free culture filtrates (B) toward Fusarium oxysporum f. sp. radicis lycopersici noted after 5 days of incubation at 25°C compared to control. IC: Untreated control; I11: Isolate from flowers; I15, I20: Isolates from leaves; I18, I14: Isolates from stems; I16, I19: Isolates from fruits. Bars sharing the same letter are not significantly different according to Duncan Multiple Range test at P ≤ 0.05.
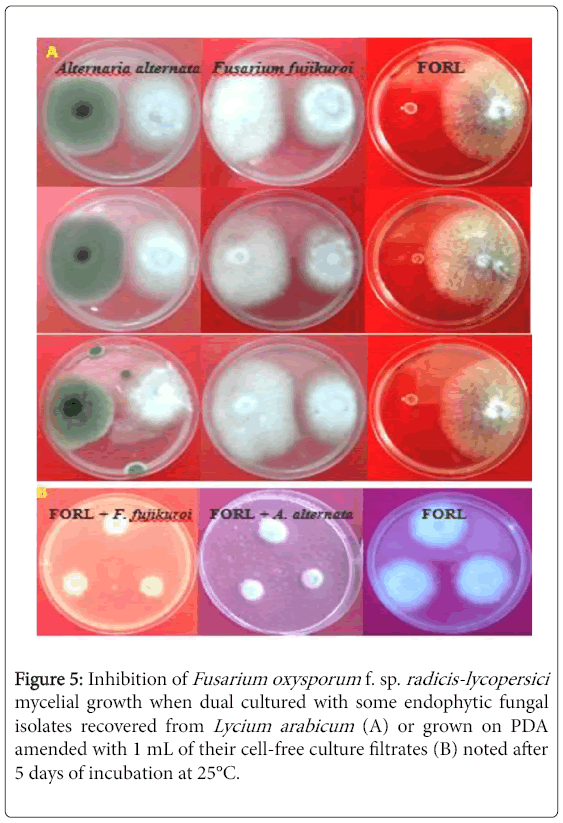
Figure 5: Inhibition of Fusarium oxysporum f. sp. radicis-lycopersici mycelial growth when dual cultured with some endophytic fungal isolates recovered from Lycium arabicum (A) or grown on PDA amended with 1 mL of their cell-free culture filtrates (B) noted after 5 days of incubation at 25°C.
Effect of cell-free culture filtrates:Five selected endophytic isolates were screened for their in vitro antifungal activity against FORL using their cell-free culture filtrates. They were chosen based on their ability to suppress Fusarium Crown and Root Rot disease severity by more than 50% and to inhibit FORL mycelial growth by direct confrontation by more than 60% over control.
ANOVA analysis revealed a significant (at P ≤ 0.05) variation in FORL mycelial growth depending on cell-free filtrates tested. In fact, Figure 4 (b) showed that the decrease in FORL mycelial growth varied from by 62.2-81.2% depending on treatments. The highest decrease in FORL mycelial growth, by about 80.9-81.2% versus control, was recorded on pathogen cultures grown on PDA medium amended with I15 and I18 filtrates. Moreover, the remaining culture filtrates tested had significantly similar effect on this parameter by decreasing FORL mycelial growth by 62.2-62.9% versus control.
Characterization of most bioactive endophytic isolates: Two isolates (namely I15 and I18) were selected based on their interesting potential to suppress the in vitro and the in vivo growth of target pathogen and their capacity to stimulate growth of tomato plants inoculated or not with FORL.
Morphologicalcharacterization: Colonies of the most bioactive fungal isolates were morphologically characterized based on colony appearance, mycelial texture and pigmentation on PDA medium when grown at 25°C.
Macrospically, colonies of I15 isolate showed a rapid growth (estimated at 7-8 mm/day). The surface texture is velvety and colony color is black. The plate reverse color is also black. As for micro-morphological traits, hyphae are septate. Conidia are formed in long chains, obclavate, obpyriform, ovoid or elipsoidal, with up to 3-5 transverse and several longitudinal septa. Conidiophores are single or in small groups, simple or branched, straight or flexuous, pale to mid olvaceous or brown. Dimension of conidia is of about 9-11×20-32 μm (Figure 6).
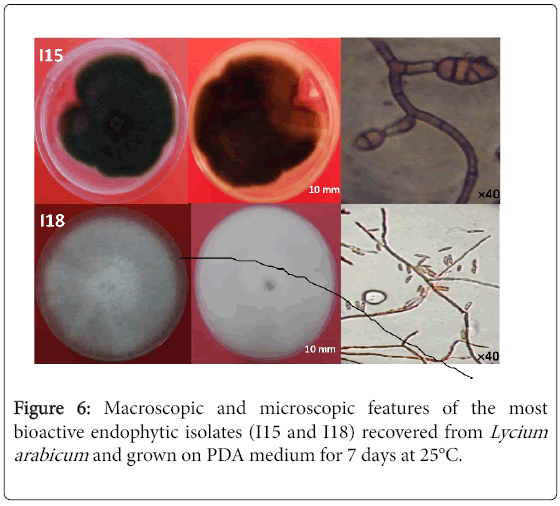
Figure 6: Macroscopic and microscopic features of the most bioactive endophytic isolates (I15 and I18) recovered from Lycium arabicum and grown on PDA medium for 7 days at 25°C.
I18 isolate grown on PDA medium showed abundant aerial mycelia that are initially white in color and later change to gray, purple or magenta. Sporodochia are orange. The macroconidia are generally uniform in shape and size, relatively thin, of medium length without perfect curvature. Apical cells have a conical morphology. Basal cells have an underdeveloped morphology. Macroconidia are septate with 3 to 5 septa. The microconidia have an oval or club-shaped shape with a flattened base without or with a partition. Chlamydospores are absent.
Molecular identification: The electrophoresis of PCR products of genomic DNA samples on 1.0% (w/v) agarose gel using a 100 bp size marker as a reference, showed bands of 600 bp for each fungus. Blast analysis of sequenced rDNA gene homology and the phylogenetic analysis based on neighbor-joining (NJ) method with 1000 bootstrap sampling revealed that the isolate I15 belonged to the genus Alternaria with 100% of similarity to Alternaria alternata (MF693801) (Figure 7A).
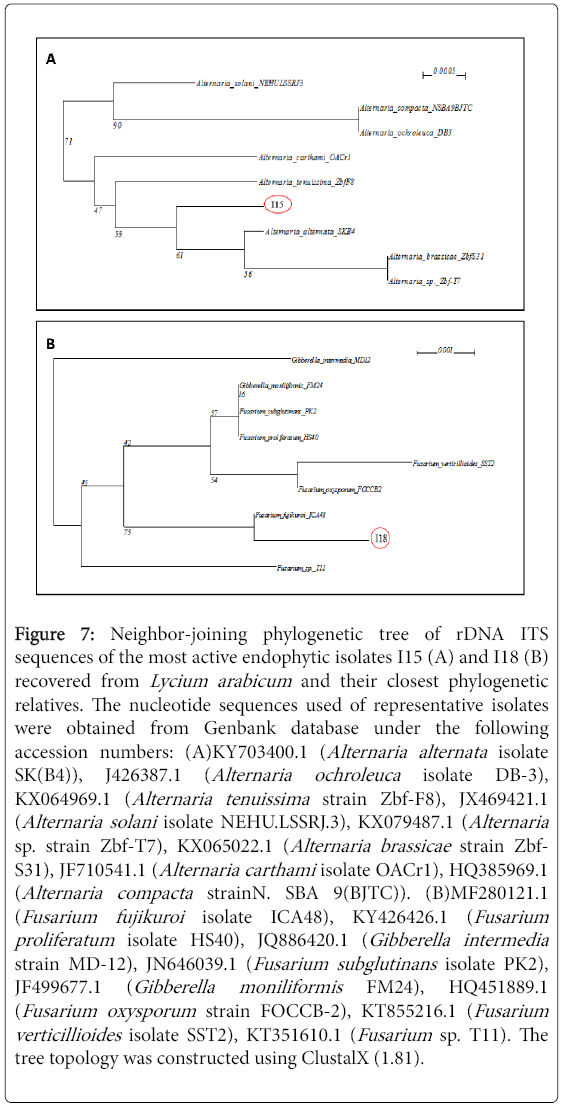
Figure 7: Neighbor-joining phylogenetic tree of rDNA ITS sequences of the most active endophytic isolates I15 (A) and I18 (B) recovered from Lycium arabicum and their closest phylogenetic relatives. The nucleotide sequences used of representative isolates were obtained from Genbank database under the following accession numbers: (A)KY703400.1 (Alternaria alternata isolate SK(B4)), J426387.1 (Alternaria ochroleuca isolate DB-3), KX064969.1 (Alternaria tenuissima strain Zbf-F8), JX469421.1 (Alternaria solani isolate NEHU.LSSRJ.3), KX079487.1 (Alternaria sp. strain Zbf-T7), KX065022.1 (Alternaria brassicae strain Zbf- S31), JF710541.1 (Alternaria carthami isolate OACr1), HQ385969.1 (Alternaria compacta strainN. SBA 9(BJTC)). (B)MF280121.1 (Fusarium fujikuroi isolate ICA48), KY426426.1 (Fusarium proliferatum isolate HS40), JQ886420.1 (Gibberella intermedia strain MD-12), JN646039.1 (Fusarium subglutinans isolate PK2), JF499677.1 (Gibberella moniliformis FM24), HQ451889.1 (Fusarium oxysporum strain FOCCB-2), KT855216.1 (Fusarium verticillioides isolate SST2), KT351610.1 (Fusarium sp. T11). The tree topology was constructed using ClustalX (1.81).
Blast analysis of sequenced rDNA gene homology and the phylogenetic analysis based on neighbor-joining (NJ) method with 1000 bootstrap sampling revealed that the isolate I18 belonged to the genus Fusarium with 100% of similarity to Fusarium fujikuroi (MF693802) (Figure 7B).
Hydrolytic enzymes production potential of the bioactive endophytic isolates
Proteolytic activity: Both isolates I15 and I18 formed clear zones around their colonies, after 4 days of incubation at 25°C, when grown on skim milk agar medium. This confirms their ability to produce protease (Table 3).
| Isolate | Amylase | Lipase | Protease | Chitinase |
|---|---|---|---|---|
| I15 | + | - | + | + |
| I18 | + | + | + | + |
Table 3: Enzymatic activity displayed by both endophytic fungi (I15 and I18) recovered from Lycium arabicum .+: Presence of enzymatic activity; -: Absence of enzymatic activity.
Amylase activity: I15 and I18 isolates grown in the Glucose Yeast Extract Peptone Agar (GYEP) medium formed a white zone around their colonies after for 4 days of incubation at 25°C. This indicates the release by the tested isolates of amylase enzyme involved in the digestion of starch in the culture medium (Tables 3 and 4).
| Isolate | Accession number | Most related species | Sequence homology (%) |
|---|---|---|---|
| I15 | MF693801 | Alternaria alternata SKB4 | 100 |
| I18 | MF693802 | Fusarium fujikuroi | 100 |
Table 4: Identification of the two most bioactive endophytic isolates (I15 and I18) by DNA sequencing genes. I15 and I18: Fungal isolates recovered from surface-sterilized Lycium arabicum leaves.
Chitinolytic activity: The degradation of the chitin is initiated by the chitinolytic mechanism process which involves the biosynthesis of chitinase. Both I15 and I18 isolates were shown able to grow on the chitin-based medium after 10 days of incubation at 25 ± 2°C. This indicates that they can produce chitinase enzyme (Tables 3 and 4).
Lipolytic activity: The presence of visible precipitate around the colony of isolate I18 indicated a positive lipase activity due to the formation of calcium salts of the lauric acid released by the lipase enzyme) after 3-7 days of incubation at 25°C. However, the isolate I15 did not (Tables 3 and 4).
Attempts of biological control of Fusarium Crown and Root Rot disease in tomato has been widely accomplished using several bacterial agents such as Bacillus subtilis [55], Pseudomonas fluorescens [56,57], and Paenibacilluspolymyxa SC09-21 [58]. Trichoderma harzianum [8], non-pathogenic Fusarium oxysporum [3], and F.equiseti [7] were also widely explored as potent bio-control agents against target pathogen.
Another bio-resource, endophytic fungi, was explored as promising antifungal and growth-promoting agents. They are known by their ability to release various bioactive metabolites involved in plant protection [59] growth promotion [60]. However, the involvement of endophytic fungi as biocontrol agents against FCRR disease is rarely considered [13].
Searching for potential source of natural compounds useful in agriculture, the Lycium genus was widely reported as an interesting source of extraction of bioactive molecules and isolation of biocontrol agents [19,32]. In the present study, a collection of fungal isolates naturally associated with L. arabicum was screened for its ability to suppress FCRR and to stimulate tomato growth when applied as conidial suspensions or cell-free culture filtrates.
Forty fungal isolates were recovered from L. arabicum leaves, stems, flowers and fruits. The frequency of isolates collected varied depending on host organs targeted for isolation. Leaves harbored 32.5% of recovered isolates compared to 27.5, 22.5, and 17.5% collected from stems, fruits and flowers, respectively. Similarly, previous studies have demonstrated that colonization rate of endophytic fungi is more prevalent in leaves of Tinosporacordifolia (29.3%), Justiciaadhatoda (69.3%) [61], and Mansoaalliacea (72.2%) [62] than in the other explored organs. In fact, this may be due to the large leaf area, which is directly exposed to the environment. In addition, the leaves contain stomata that act as a pathway for fungal mycelium penetration and further tissue colonization [63].
According to their macroscopic and microscopic traits, fungal isolates recovered from L. arabicum were affiliated to five genera namely Fusarium, Alternaria, Penicillium, Aspergillus, and Trichoderma. Alternaria was found to be the most dominant genus with a relative isolation frequency of 10% followed by Fusarium (5.83%). Alternaria and Fusarium associated to Lycium species were reported in previous studies. For instance, these both genera were recovered from L. chinense leaves [64] and from L. barbarum leaves, stems, and fruits [65]. Furthermore,A. alternata was also found to be the dominant endophyte associated to Aegle marmelos [66] and Justiciaadhatoda (17.03%) [61].F. fujikuroiwas also frequentlyrecovered fromPaepalanthuschiquitensis [67,68].
Seven fungal isolates associated to L. arabicum were evaluated for their capacity to control Fusarium Crown and Root Rot disease under greenhouse conditions using their conidial suspensions or cell-free culture filtrates. Results from the current study clearly demonstrated that I15 and I18 isolates exhibited the highest disease-suppressive effects, by reducing leaf and root damage intensity by 85.7-92.8% and the vascular browning extent by 94.4-98.4% when applied as conidial suspensions or filtrates. These both isolates were identified based on rDNA sequencing as A. alternata (MF693801) and F. fujikuroi(MF693802). They were also found to be the most efficient in stimulating growth of tomato plants already challenged with FORL. Furthermore, Pearson’s correlation analysis confirmed the link between the decrease in FCRR severity and FORL colonization of tomato tissues and the recorded growth promotion. This indicates that wild L. arabicum could be an effective source of isolation of effective bio-agents, able to colonize and to protect cultivated tomato plants and to protect them against FCRR disease. In this regard, many fungal endophytes are shown capable to produce a variety of extracellular metabolites responsible for the protection of their host plants from their associated phytopathogens [64,69]. Interestingly, Alternaria spp. was reported to be a potential producer of bioactive metabolites such steroids, terpenoids, pyrones, quinones and polyphenols and which are involved in their antimicrobial activities [69,70]. Other previous studies demonstrated that A. alternata (KP714382) and A. tenussima (KP714380 and KP714381), endophytes associated with Panaxnotoginseng leaves and seeds, had significantly reduced disease severity of Panaxnotoginseng caused by Phomaherbarum [71].
In the present study, the ability of conidial suspension preparations and cell-free culture filtrates from the tested endophytic fungi to improve root and shoot growth of treated tomato seedlings compared to pathogen-free ones was clearly demonstrated. I15 (A. alternata; MF693801) and I18 (F. fujikuroi; MF693802) conidial preparations were found to be the most effective in improving by 73.7-76.7% and 82.6-83.7% root and shoot fresh weights, respectively, over the untreated control. In addition, cell-free filtrates from these both isolates led to the highest enhancement of tomato growth by 88.5-91.3% over control. This growth promotion can be achieved directlythrough the antagonistic activity of endophytic agents against target pathogenic fungi or indirectly through an activation of plant defense. In fact, this improvement in treated plants is due to an enhancement of nutrient uptake and ability to produce various bioactive metabolites and enzymes, including phytohormones [30,72]. Indeed, fungi are well known as a potential producer of phytohormones which are involved in the promotion of plant growth [25]. In this context, similar findings had also demonstrated the ability of F. fujikuroi to enhance the growth of Oryza sativa and Atriplexgemelinii plants through the production of gibberellins (GA3) [73,74]. In other studies, endophytic A. alternata recovered from Solanum nigrum leaves had significantly improved root elongation, height and biomass of rice plants as well as growth of maize plants. These effects have been explained, by the ability of this endophyte to release indole acetic acid [74]. Moreover, endophytic A. alternata isolated from Asclepiassinaica leaves had also significantly improved root elongation of Zea mays plants [75].
Tested using the dual culture method, the tested endophytic fungal isolates exhibited a significant antifungal activity toward FORL. In fact, the highest inhibition of pathogen radial growth, by about 77.2-78.1%, was achieved using I15 (A. alternata; MF693801) and I18 (F. fujikuroi; MF693802) isolates. Also, these both isolates were found to be potential protease-, chitinase-, and amylase-producing agents. Thus, this interesting antifungal potential could be explained, among others, to the capacity of these fungal isolates to inhibit FORL growth via the biosynthesis of extracellular cell wall-degrading enzymes such as chitinases, proteases and amylases. In fact, based on previous studies, endophytic fungi can produce extracellular hydrolases as a resistance mechanism against pathogenic invasion. Such enzymes include pectinases, cellulases, lipases, and laccase [76]. For example, the endophytic fungus A. alternata (PCTS21) had reduced the growth of F. solani and this effect was explained by its ability to produce extracellular enzymes such as amylase, lipase, cellulase, and chitinase [77]. On the other hand, extracellular metabolites present in cell-free culture filtrate of the endophytic fungi, tested at 10% (v/v) in this study, were found to be effective in suppressing FORL in vitro growth. Interestingly, filtrates of I15 (A. alternata) and I18 (F. fujikuroi) induced the highest decrease in FORL mycelial growth (by 80-81.4% over control). In this context, several antifungal metabolites produced by the endophytic A. alternata have been widely reported. For example, the cyclo- (Phe-Ser) metabolite produced by the endophytic Alternaria sp. FL25 isolated from Ficuscarica had significantly inhibited the growth of many fungal plant pathogens such as F. graminearum, F. oxysporum f. sp. cucumerinum, F. oxysporum f. sp. niveum, Phytophthora capsici, and Colletotrichumgloesporioides with a minimal inhibitory concentration (MIC) of 6.25-25 μg/mL [78]. In another study, this fungal endophyte was found able to release another metabolite identified as helvolic acid and displayed an important antagonistic activity against various pathogens [78]. Other antifungal metabolites identified as herbarin A and Chromone A were released by A. brassicicola ML-P08 [79]. Alterperylenol and dihydroalterperylenol were also released by Alternaria sp. [80] displaying antifungal activity against Aspergillus niger and Valsaceratosperma. In another study, the endophytic fungus F. fujikuroi was shown able to produce other antimicrobial metabolites such as alkaloid 2-(4-butylpicolinamide) acetic acid, fusaric acid, indole acetic acid, a sesquiterpene identified as terpestacine which may be involved in its antifungal potential [67].
Endophytic fungi are potentially interesting sources of bio-based products useful in sustainable agriculture. To the best of our knowledge, L. arabicum was firstly explored in the current study as a potential source of isolation of endophytic fungi with antifungal potential against FORL. The present study led to the selection of two bioactive endophytes which were shown efficient for Fusarium Crown and Root Rot control and for the bio-stimulation of tomato growth. According to rDNA gene sequencing, the most bioactive endophytic fungi were identified as Alternaria alternata (Isolate I15; MF693801) and Fusarium fujikuroi (Isolate I18; MF693802). Interesting enzymatic activities (chitinase, protease, lipase and amylase) were demonstrated for these two selected isolates and seemed to be responsible for their antifungal potential against FORL. Our study suggests that wild Solanaceous species are interesting sources of promising endophytic fungal isolates with FCRR suppression and bio-fertilizing abilities.
This work was funded by the Ministry of Higher Education and Scientific Research of Tunisia through the funding allocated to the research unit UR13AGR09-Integrated Horticultural Production in the Tunisian Centre-East, The Regional Center of Research on Horticulture and Organic Agriculture of Chott-Mariem, Tunisia.