Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Editorial - (2013) Volume 2, Issue 6
Barley is a short-season early maturing crop, which is cultivated in a variety of climates, including both irrigated and dryland production areas. In fact, barley is the third largest feed grain crop produced in the United States (US), after corn and sorghum. However, extensive application of imidazolinone (IMI) herbicides resulted in significant drop in its production due to continuous decline in the barley acreage over the last two decades. In 2011, barley touched the all time low acreage of 2.24 million acres in the US with an average yield of 69.6 bushels/acre, which was approximately one-half of the year 2000 production level. Washington State, which was ranked fourth in the whole nation for barley production in 1999 with 10.3% of the US crop planted over 500,000 acres, the acreage declined to 85,000 planted acres in 2010 [1], USDA Economic Research Service-Feed Grains [2]. This significant drop in the acreage during the last two decades can be attributed to wide scale application of imidazolinone herbicides, in combination with the introduction of IMI-resistant crops. The decline in acreage can also be explained by the overlapping distribution of regions under barley cultivation in the US Pacific Northwest (PNW) and the regions under extensive application of Imazamox (Beyond), and/or Imazethapyr (Pursuit) (Figure 1). Collectively, the major reason for the decline in barley acreage is its sensitivity to commonly used herbicides. Many of the widely used herbicides imposing barley plant-back restrictions belong to the group B herbicides [3]. Group of herbicides belonging to imidazolinone family targets acetolactate synthase (ALS) or acetohydroxyacid synthase (AHAS), an octameric enzyme with four catalytic and four regulatory subunits. AHAS catalyses two parallel reactions in the synthesis of branched chain amino acids. The first reaction is condensation of two pyruvate molecules to yield acetolactate leading to the production of valine and leucine, and the other reaction is the condensation of pyruvate and α-ketobutyrate yielding acetohydroxybutyrate leading to the production of isoleucine. The AHAS-inhibiting herbicides are known to bind at the substrate access channel, blocking the path of substrate to the active site. When AHAS is inhibited, deficiency of the amino acids (valine, leucine and isoleucine) causes a decrease in protein synthesis, which in turn, slows down cell division rate. This process eventually kills the plant showing symptoms in meristematic tissues, where biosynthesis of amino acids primarily takes place. Resistant plants in most cases depend on reduced sensitivity to these herbicides by an isoform of AHAS, which does not severely affect its catalytic activity. Most AHAS isoenzymes resistant to the herbicides carry substitutions for the amino acid residues Ala122, Pro197, Ala205, Asp376, Trp574 or Ser653 (amino acid numbering refers to the sequence in Arabidopsis thaliana). The amino acid residues Ala122, Pro197 and Ala205 are located at the N-terminal end of the enzyme, whereas Asp376, Trp574 and Ser653 are located at the C-terminal end. Amino acid substitutions at Ala122 and Ser653 confer high levels of resistance to imidazolinone herbicides, whereas substitutions at Pro197 endow high level of resistance against sulfonylureas and provide low-level resistance against imidazolinone and triazolopyrimidine herbicides. Substitutions at Trp574 endow high levels of resistance to imidazolinones, sulfonylureas and triazolopyrimidines, while substitutions at Ala205 provide resistance against all AHAS-inhibiting herbicides [4].
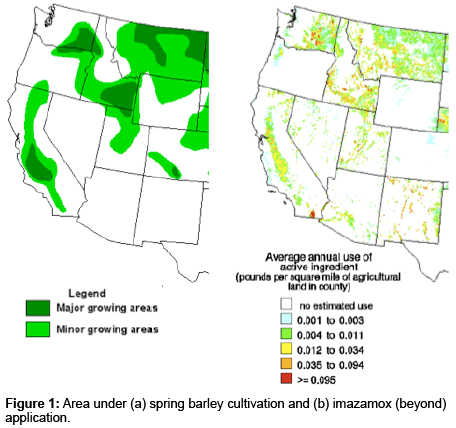
Figure 1: Area under (a) spring barley cultivation and (b) imazamox (beyond) application.
In the case of barley, which is a common highly profitable rotational crop following wheat, no IMI-resistance is reported for any of the varieties cultivated in the PNW. Thus, introduction of barley varieties with resistance to imidazolinone herbicides will provide greater flexibility as a rotational crop. An IMI-resistant mutant was isolated from an extensive screen of 2 million seeds of a sodium azide treated ‘Bob’ population. Molecular characterization of the mutant revealed an amino acid substitution in the substrate access channel of the catalytic subunit of AHAS enzyme, changing a Serine to Asparagine at amino acid location 653 [5]. This mutation in substrate access channel does not allow imazamox to block the path of substrate to the active site, and thus, functions normally and allows plants to survive the field recommended dose of imidazolinone (4 oz/Acer for winter wheat), with no obvious influence on the plant fitness. Development of IMI-resistant barley cultivars by crossing Bob AHAS mutant with barley cultivars adapted to the US PNW will have many advantages: For instance, the lines are already adapted to the PNW, have excellent food, feed or malting quality and the final product will be non-transgenic in origin. In addition, IMI-resistant barley has lesser risk of introducing herbicide resistance to the most common monocotyledonous weeds in the barley fields. In contrast, IMI-resistant wheat has a potential risk of transferring herbicide resistance to jointed goat grass (Aegilops cylindrica), which shares its ‘D’ genome with common wheat.
To effectively breed for IMI-resistance, it is a prerequisite to develop resources, which will allow precise transfer of the trait of interest in a single generation to the desired genotype, without a need of backcrossing. Towards this end, the following experiments were undertaken: i) Estimation of genetic diversity among 13 two-rowed spring barley cultivars/breeding-lines adapted to the US PNW, using 61 carrier chromosome 6H-specific microsatellite markers. Based on this genotypic-information, two genotypes per market class (food, feed and malting) were selected and crossed with the ‘Bob’ AHAS mutant. (ii) A micro-scale enzyme extraction and in vitro colorimetric enzyme activity assay for quick detection of the AHAS mutant allele in the segregating population was optimized.
Building upon the above results, a range of 29 to 53 crosses were made per genotype combination with Bob AHAS mutant. The ‘WAS4’ and Bob mutant crosses showing maximum genetic dissimilarity are currently used for genetic mapping of the AHAS gene on barley chromosome 6H. More than 2100-2800 F2 grains obtained per cross combination were screened for IMI-resistance in the glasshouse, and the survivors showing vigorous phenotype will be examined for carrier chromosome recovery.
So far, successfully transfer of IMI-resistance from ‘Bob’ AHAS mutant was confirmed in two feed barley cultivars Lyon and Muir, two food barley cultivars Clearwater and WAS4 and a malting barley cultivar Radiant. We confirmed transfer of trait by the herbicide spray at the rate of 8 oz/Acer with methylated seed oil, which is equivalent to the 2× field, recommended dose on winter wheat and by genotyping the point mutation providing IMI-resistance by DNA sequencing. The segregation rations from all six crosses obtained from the greenhouse spray test suggested dominant nature of the mutant. We have also successfully standardized the colorimetric enzyme activity assay and confirmed that the mutant enzyme can survive up to 40 oz/Acer beyond (i.e. 10 times field recommended dose on winter wheat), without any effect on the enzyme activity. This can be monitored in the test by production of red colour in presence of herbicide by the mutant enzyme. To bring the IMI-resistance barley genotypes from laboratory to field, it is now necessary to follow the following objectives: 1) screen progeny of F2 lines that survived 8oz/Acer beyond with methylated seed oil and showed early vigor a month after spray, for the genotype at the acetohydroxyacid synthase locus and for carrier chromosome (6H) recovery. 2) Fix all segregating loci by converting lines showing good carrier chromosome recovery into doubled haploids. This will avoid confounding of results of phenotypic evaluations in field, and 3) evaluate the doubled haploid lines for their performance on herbicide residue and under spray trials in field.
In a nutshell, this research will result in the development of IMIresistant food, feed and malting barley cultivars adapted to the US PNW and will re-establish barley as a rotational crop with winter wheat.
This project was supported by the Washington Grain Commission Grant 13C- 3019-3590.