Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2014) Volume 4, Issue 1
Keywords: Calcium, Homeostasis, Saccharomyces cerevisiae, Vacuole
Calcium signaling
Calcium is a universal signaling molecule used by all eukaryotic cells. The concentration of Ca2+ ions in the extracellular fluid relative to the intracellular concentration is approximately 1000 times higher. The higher concentration of Ca2+ outside of the cell creates a downhill electrochemical gradient that allows Ca2+ ions to move passively and quickly across the membrane. This also occurs with select organelles inside the cell. Organelles such as the Endoplasmic Reticulum (ER), Sarcoplasmic Reticulum (SR) and the vacuole maintain a higher concentration of Ca2+ ions in the lumen versus the cytosol. Ca2+ pumps are located in the membranes of the cell and organelles, which is used to pump Ca2+ ions out of the cell or into the lumen of the various organelles [1]. The cellular processes regulated by intracellular Ca2+ binding include the activation of secondary messenger molecules; exocytosis of signaling molecules; muscle contraction; egg cell fertilization; and cell division [2].
| Name | Genome | Source |
|---|---|---|
| BY4742 | Mat α, his3Δ1, leu2Δ0, lys2Δ0, ura3Δ0 | ResGen |
| BY4742 Dyvc1 | BY4742 yvc1::URA3 | This project |
| jgy149 | Mat a ade2-10c can1-100 his3-11,15 leu2-3,112 trp1-1 ura3-1 cmd1-6 URA3 5FOA | [20] |
| ykl190w | BY4742 cnb1::kanMX | ATCC |
| ynl027w | BY4742 CRZ1::kanMX | ATCC |
| ykl190w Δyvc1 | ykl190w yvc1::URA3 | This project |
| yfr014c | BY4742 cmk1::kanMX | ATCC |
| yfr014c Δyvc1 | yfr014c yvc1::URA3 | This project |
| yol016c | BY4742 cmk2::kanMX | ATCC |
| ydl128w | BY4742 vcx1::kanMX | Invitrogen |
Table 1: Yeast strains used in this study.
Saccharomyces cerevisiae is a model organism
The budding yeast, Saccharomyces cerevisiae, serves as a model organism for laboratory research. Yeast cells and higher eukaryotic cells also share many homologous proteins [3]. In fact, Cui et al. [4], compared key homologues of the regulatory proteins involved in intracellular calcium homeostasis and signaling transduction in both S. cerevisiae and mammalian cardiomyocytes. Aberrant changes in the mechanisms involved in intracellular calcium homeostasis and signaling transduction in mammalian cardiomyocytes are suspected to play a part in congestive heart failure and mechanical dysfunction. One study found that a decrease in the expression of the SR Ca2+-ATPase, which is responsible for removing Ca2+ from the cytosol following contraction, and an increase in a Na+-Ca2+ exchanger (NCX) expression, which is responsible for bringing Ca2+ into the cell, was seen in instances of heart failure [5]. This makes yeast calcium research important for gaining greater insight into the mechanics of mammalian cardiomyocyte functions. The identification of the homologous regulatory proteins between S. cerevisiae and mammalian cardiomyocytes is an advantage to assist in understanding how both cells function.
Calcium homeostasis in yeast
Ca2+ signal transduction in the budding yeast, S. cerevisiae, is involved in cell division and pheromone signaling [6]; responses to hypertonic shock and cold stress resistance [7]; and the secretory pathways [1]. The basal concentration of intracellular Ca2+ in yeast is approximately 50-200 nM [8]. In yeast the extracellular concentrations can range from <1 μM to >100 mM, yet the cytosolic Ca2+ is maintained at 50-200 nM. Thus, yeast cells must have mechanisms in place to tightly maintain the concentration of intracellular Ca2+ and not allow it to vary with extracellular concentration. The lower the concentration of free Ca2+ in the cytosol, the more sensitive the cell is to an increase, which lends to increased effectiveness of Ca2+ as a signaling molecule.
Much work has been done in an attempt to elucidate the mechanisms used by S. cerevisiae for maintaining this intracellular Ca2+ homeostasis. One pathway that has been studied extensively is that of the High Affinity Ca2+ System (HACS). Located in the cell plasma membrane, HACS consists of a putative voltage-gated Ca2+ channel of which two identified proteins are encoded by CCH1 and MID1. It is postulated Cch1p makes up the pore of the channel, while the function of Mid1p is that of a regulatory subunit [9]. HACS is only activated to bring in extracellular Ca2+ under certain cellular conditions: α-factor pheromone signaling [6] that may require potassium as well [10], low secretory Ca2+ [11], and hyperosmotic shock [7].
Upon activation of HACS, extracellular Ca2+ enters through the high affinity Cch1p/Mid1p channel. From there, depending on what the cell needs, cytosolic Ca2+ has a multitude of destinations. Ca2+ is taken up by the vacuole and ER/Golgi apparatus through membrane transporters. The yeast gene PMC1encodes a localized vacuolar Ca2+ -ATPase transporter (Pmc1p) for the uptake of cytosolic Ca2+. An H+/ Ca2+ exchanger, encoded by Vcx1, is also localized to the yeast vacuole for the uptake of cytosolic Ca2+ [4]. The yeast vacuole is the main storage center for cellular calcium holding approximately 90% of the cell’s calcium reserves [8]. This calcium storage is used for intracellular Ca2+ signaling.
The ER/Golgi apparatus sequesters cytosolic Ca2+ through another Ca2+ -ATPase, encoded by PMR1. The ER/Golgi apparatus is not only a minor storage unit for cellular calcium, but the signal transduction induced by Ca2+ binding in the ER lumen activates a number of secretory pathway processes, such as required proteases for activation or modification of newly synthesized proteins [8].
Further work has demonstrated additional calcium channels in the plasma membrane. Muller et al. [6] also delineate the possible existence of a Low Affinity Ca2+ Influx System (LACS) located in the cell plasma membrane. Their study found HACS and LACS are both activated by α-factor mating pheromones under varied circumstances. In the presence of α-factor, HACS is activated under low nutrient media when the activity of the calmodulin-dependent phosphatase calcineurin is inhibited, and LACS is activated in rich media or high levels of extracellular Ca2+ [6]. Furthermore, a Mid1p-dependent Cch1p independent calcium flux was shown in response to amiodarone [12]. Finally, Cui and Kaandorp [13] postulated the existence of an unidentified plasma membrane Ca2+ channel called Channel X that is separate from the Cch1p/Mid1p channel and through which extracellular Ca2+ enters the cell during normal conditions, although this channel has not yet been identified.
In response to an elevated cytosolic Ca2+ level, the plasma membrane channel(s) responsible for the influx into the cell is/are inhibited and cytosolic Ca2+ is sequestered by the vacuole H+/Ca2+ exchanger (Vcx1p), the vacuolar ATPase, Pmc1p, and Pmr1p [1]. Cytosolic Ca2+ also initiates a phosphatase signaling cascade by binding to a Ca2+ responsive protein calmodulin. Ca2+-bound calmodulin binds to the phosphatase protein calcineurin complex. Calcineurin has multiple functions relating to Ca2+ signaling pathways. It inhibits the action of Vcx1p; activates the transcription factor Crz1p, which translocates to the nucleus to induce gene transcription of relevant target proteins; and regulates the Cch1p/Mid1p channel in addition to the regulation of other membrane ion transporters. Gene transcription of target proteins activated by Crz1p is relative to the response the cell requires to return to homeostasis of its physiological condition. There are approximately 160 genes activated by Crz1p in response to cellular stress including increasing expression of PMC1 and PMR1 [14]. Two protein kinases, Cmk1p and Cmk2p are activated by Ca2+-bound calmodulin as well [1]; although their exact substrates are unknown in this process. Excess Ca2+ is able to leave the cell through the secretory pathway via Pmr1p [4,15] and a plasma membrane exporter is hypothesized to exist as well [16].
The Yeast vacuole and its role in calcium homeostasis
The yeast vacuole is responsible for protein degradation, cellular ion and pH regulation, and the storage of amino acid precursors [17]. The lumen of the vacuole is kept to a lower pH than the surrounding cytosol at approximately pH 3-5 [18]. Many metabolites and ions cross the vacuolar membrane through antiporters utilizing the proton motive force created by the activity of the vacuole H+-ATPase. The V-H+-ATPase uses the energy of ATP hydrolysis to move H+ ions into the vacuole, which maintains a proton gradient.
The calcium concentration in the vacuole lumen is in the millimolar range compared to the nanomolar range of the cytosol. The H+/ Ca2+ exchanger, Vcx1p, uses the energy provided by the proton motive force to uptake Ca2+ into the vacuole lumen. Two H+ ions are released to the cytosol for each Ca2+ ion sequestered. The active transport of Ca2+ ions across the vacuole membrane also occurs through the Ca2+-ATPase. Pmc1p uses hydrolysis of ATP to move Ca2+ ions up their concentration gradient into the vacuole [18]. Pmc1p is sensitive to the cytosolic Ca2+ concentration, and has a binding affinity of approximately 4.3 μM [18]. Small changes in the cytosolic Ca2+ concentration can activate the binding of ATP to Pmc1p to induce Ca2+ transport [18]. Vcx1p is only activated during high levels of cytosolic Ca2+.
The vacuole is also responsible for expelling Ca2+ as a secondary messenger in signal transduction, and the expulsion is activated by hypertonic shock. Calcium is expelled through an ion channel encoded by Yvc1 [19]. Immediately after the expulsion, the Ca2+ is taken back up into the vacuole through the H+/Ca2+ exchanger, Vcx1p [19].
The complete mechanisms of all of the factors involved in yeast calcium homeostasis have not been elucidated. We study the key proteins involved in the mechanisms of cell response and return to homeostasis after calcium shock in Saccharomyces cerevisiae. Our assay consists of treating cells with a single pulse of 100 mM (final concentration) CaCl2 and measuring the cytosolic aequorin luminescence over a thirteen minute timeframe. Our work has verified key players involved in a Ca2+ induced stress response to be CMD1, CNB1, CRZ1, CMK1, and CMK2. We, and others, have shown that a rapid feedback response sequesters cytosolic Ca2+ into the vacuole via both Vcx1p and Pmc1p and into the ER/Golgi via Pmr1p [4,15,21,22]. We are interested in identifying the roles of each key player. Cytosolic aequorin luminescence of strains harboring deletion mutants of the genes (cmd1-6, Δcnb1, Δcmk1, and Δcmk2), demonstrated anomalous behavior after a single calcium pulse of 100 mM CaCl2 (final concentration). Cytosolic Ca2+ in these cells peaked after the pulse, returning to just above baseline, but then deviated from wild type response with a second rise in cytosolic calcium at approximately 6 minutes (Results). We wanted to identify the origin of the second rise in cytosolic Ca2+. We ascertained the source could be an extracellular or intracellular pool of Ca2+. Previous work also showed a peak in cytosolic calcium after a single pulse of 100 mM CaCl2 in Δcch1 Δmid1 strain (data not shown). We concluded from those results that HACS was not the origin; therefore we speculated if the leak was an extracellular Ca2+ source then it could be the unidentified putative Channel X proposed by Cui et al. [4]. Since Yvc1p is the localized vacuole exporter of Ca2+ during times of stress [19], and there is an unidentified hypothesized Ca2+ channel X [13], we hypothesized the origin of the cytosolic leak in Ca2+ was coming from Yvc1p, or the plasma membrane through the unidentified channel. We tested this by measuring cytosolic aequorin luminescence of double mutant strains, Δcnb1 ΔYvc1 and Δcmk1 ΔYvc1 after an extracellular single pulse of 100 mM CaCl2. Additional work focused on whether Vcx1p was acting backwards and producing the second rise in cytosolic calcium. We show that Δcmk1 ΔYvc1 cells no longer demonstrate the second rise in calcium, implicating Cmk1p in regulating Yvc1p, and that Vcx1p does not work in reverse to expel calcium from the vacuole into the cytosol.
DNA transformations
Yeast DNA transformations were conducted by using a Zymo Research Frozen-EZ Yeast Transformation II Kit™ (Zymo Research Corporation, Irvine, CA), following the manufacturer’s instructions. The yvc1 gene knockout was made by transforming the cells with linearized pdy3u DNA (cut with SacII, New England Biolabs, Ipswich, MA), a kind gift of Dr. Yoshiro Saimi from University of Wisconsin, Madison. This plasmid contained the 5’ and 3’ ends of the Yvc1 gene with a complete deletion of the ORF replaced with URA3. Cells utilized for the luminometry assay were transformed with a pEVPII/Aeq plasmid containing the APOAEQUORIN gene for the bioluminescence assays [7]. The transformed cells were grown on the appropriate dropout media.
Confirmation of yvc1 knockout through PCR analysis
Following transformation of the cells with the Yvc1 knockout plasmid (pdy3u), a single colony from the transformed culture plate was isolated on a sterile –URA synthetic dextrose agar plate using the streak plate technique. Approximately 10 μl of cells from each quadrant of the isolation plate were directly placed into the PCR reaction (Invitrogen Platinum®, Life Technologies, Carlsbad, CA) with the YVC1-1 and YVC1-3 oligonucleotides. Agarose gel electrophoresis was used to analyze the PCR reactions. A 2.8 kb band size indicated that the knockout was made and compared to a PCR with the same oligonucleotides on the original pdy3u plasmid as a positive control.
PCR primers
The oligonucleotide primers, forward primer: YVC1-1: 5’CTTCGAGGATCAGCTAGAGCC and reverse primer: YVC1-3: 5’GCAGTATTTGAACACATGTCG, were obtained from Integrated DNA Technologies (Coralville, Iowa) and used to determine integration of the yvc1 knockout construct. These oligonucleotides reside in the 5’ and 3’ untranslated regions of the Yvc1 gene, respectively.
Bioluminescence assays
Sample Preparation: 2-3 colonies of the transformed strain were grown overnight at 30°C in YNB glucose media plus the appropriate supplements. Then the cells were back diluted and grown until the concentration reached an OD600 of 0.4. Then, 5 mL of cells were pelleted by centrifugation and resuspended in 400 μL of YNB glucose. 200 μL cell aliquots were transferred to microcentrifuge tubes. 10 μL of 590 μM coelenterazine (Gold Bio Technology, St. Louis, MO) (resuspended in methanol) was added to each sample to reconstitute the aequorin protein [8]. The samples were then incubated in a shaker in the dark at 30°C for 20 minutes. After incubation, the samples were harvested by centrifugation and resuspended in 200 μL of YNB glucose and kept in the dark until the assay was run.
Bioluminescence: Each sample was treated with a CaCl2 solution using a Sirius Luminometer 9600 (Berthold Detection Systems – USA, Oak Ridge, TN). The cells were pulsed with final concentration 100 mM CaCl2 at 30 seconds, and the cytosolic aequorin luminescence was followed for 13 minutes. The cells were then lysed with 5% Triton X-100 supplemented with 2.5 M CaCl2 to calculate the total cytosolic calcium concentration [7]. Each graph is a representative of at least 6 experiments
Hyperosmotic shock test: The bioluminescence assay protocol was followed above, with the exception that the cells were treated with 800 mM of NaCl in place of the CaCl2 as an additional method to serve as a control to verify the inactivity of Yvc1p by putting the cells into hyperosmotic shock [19] (data not shown).
Inhibition of calcineurin: In order to determine a possible reversal of Vcx1p action when calcineurin is inhibited, FK506 was used to treat wild type cells (BY4742) and the strain ydl128w with a genotype of ΔVcx1. 4μg ml-1 of FK506 (Sigma, St. Louis, MO, dissolved in DMSO) was added to the back dilution and coelenterazine treatment to achieve calcineurin inhibition.
Statistical analysis
A ratio was calculated for each replicate of the calcium concentration at 4.5 minutes / the calcium concentration at 11 minutes. These replicate ratios were compared between wild type cells and each variant using an unpaired T-test, equal variance, two tails, to determine if the calcium rise were statistically different than the wild type change
Several mutants in the calcium signaling pathway demonstrate a calcium leak after approximately six minutes. Our goal of this study was to identify the mechanisms of cell response and return to homeostasis after calcium shock in yeast cells. We assayed this by treating aequorin expressing cells with an extracellular pulse of 100 mM CaCl2 (final concentration) at 30 seconds into the assay, and measuring the cytosolic luminescence over a thirteen minute timeframe. Wild type cells (Figure 1A) increased cytosolic calcium after the CaCl2 pulse, and then cytosolic levels decreased too slightly above the pre-pulse baseline. As seen in Figure 1B-F, mutant strains (cmd1-6, a mutated calmodulin that can no longer bind calcium [20]; cnb1; crz1; cmk1; and cmk2; respectively) had a cytosolic calcium spike after the CaCl2 pulse, and the cytosolic levels decreased temporarily, followed by a second rise in cytosolic calcium after approximately six minutes. The second rise did not display the sharp increase that the initial cytosolic rise does. To determine if this second rise is statistically significantly different than the wild type cytosolic calcium level, since the rise is reproducible but not large, a ratio of the calcium concentration at 4.5 minutes (after the initial cytosolic rise and subsequent fall)/ the calcium concentration at 11 minutes (during the second rise on the graph) was calculated for each replicate and these ratios were compared to the wild type using a two-tailed equal variance unpaired T-test. In each case, the ratio is statistically significantly different than wild type, P<0.05. Interestingly, the second rise appears to be seepage or leakage rather than an extrusion as the initial rise is.
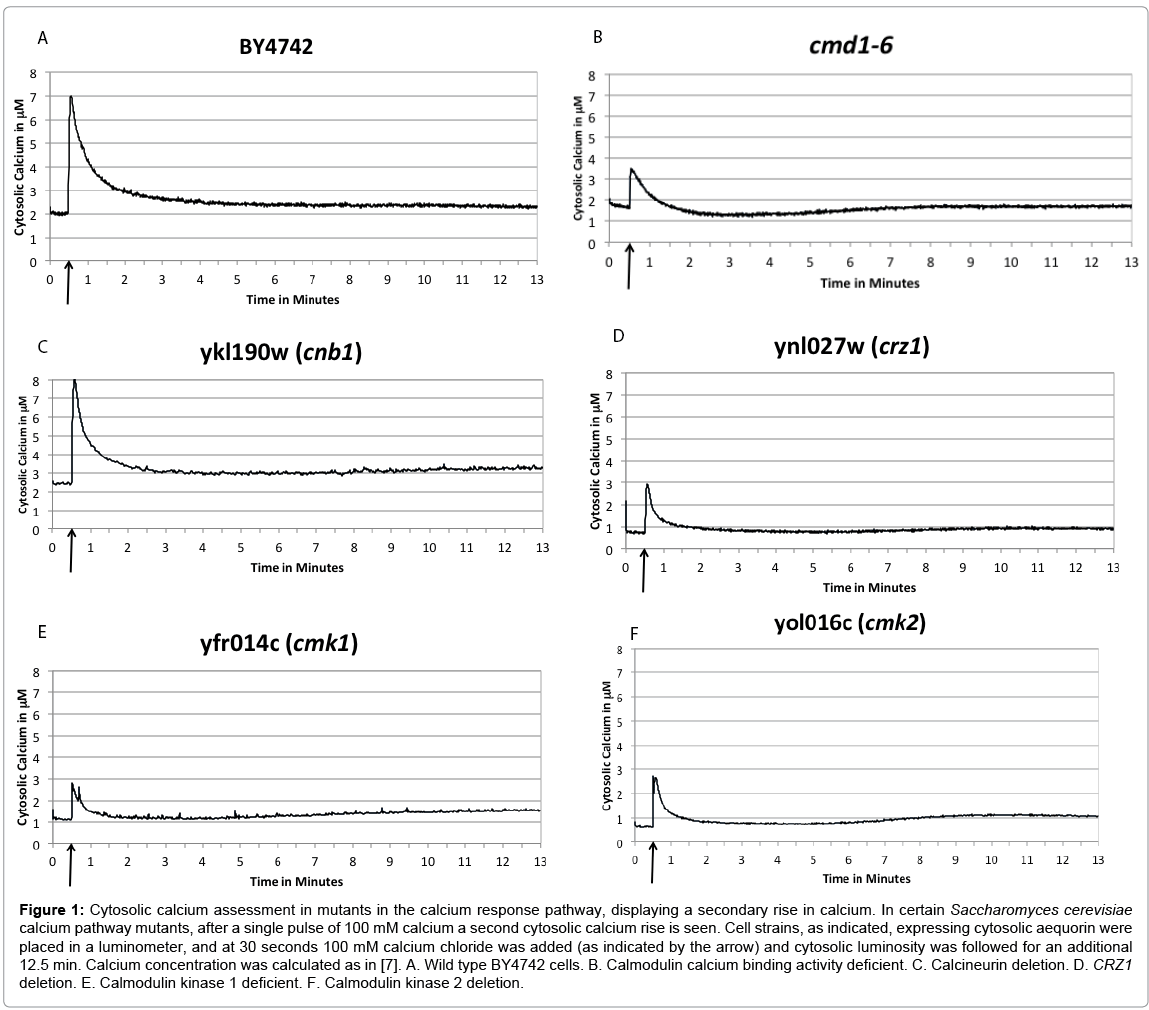
Figure 1: Cytosolic calcium assessment in mutants in the calcium response pathway, displaying a secondary rise in calcium. In certain Saccharomyces cerevisiae calcium pathway mutants, after a single pulse of 100 mM calcium a second cytosolic calcium rise is seen. Cell strains, as indicated, expressing cytosolic aequorin were placed in a luminometer, and at 30 seconds 100 mM calcium chloride was added (as indicated by the arrow) and cytosolic luminosity was followed for an additional 12.5 min. Calcium concentration was calculated as in [7]. A. Wild type BY4742 cells. B. Calmodulin calcium binding activity deficient. C. Calcineurin deletion. D. CRZ1 deletion. E. Calmodulin kinase 1 deficient. F. Calmodulin kinase 2 deletion.
Determination of the source of the calcium leak
As seen in Figures 1B-F, mutant strains (cmd1-6, cnb1, crz1, cmk1, and cmk2) increased cytosolic calcium after the CaCl2 pulse, and the cytosolic levels decreased temporarily, followed by an increase in cytosolic calcium after approximately six minutes. We hypothesized that the Ca2+ leak was from the vacuole and deleted the sole vacuole outward Ca2+ channel, Yvc1, to determine if this were the case. We analyzed the double mutants, Δcnb1 ΔYvc1 and Δcmk1 ΔYvc1, for the origin of the calcium leak.
Synthetic lethal interactions in calcium homeostasis
We explored the origin of the Ca2+ leak in the mutant strains. We hypothesized it was either originating from the vacuole through Yvc1p or through the uncloned plasma membrane Channel X [13]. We created double mutants of cmd1-6 ΔYvc1, Δcnb1 ΔYvc1, Δcmk1 ΔYvc1, and Δcmk2 ΔYvc1. We did not test a double mutant of Δcrz1 ΔYvc1, as Crz1p is a transcription factor that acts after the initial response to calcium to up regulate key players, and we wished in this initial analysis only to explore the protein-mediated response that restores calcium homeostasis in yeast. To ensure the integration of the Yvc1 knockout construct was successful we tested the strains in two ways.
(1) PCR analysis of the transformed cells tested positive for the band specific to the pdy3u Yvc1 knockout construct, indicating successful integration.
(2) We induced hyperosmotic shock to ensure the vacuolar Ca2+ channel (Yvc1p) was not expressed in the cells. In a wild type cell with a fully functioning vacuolar Ca2+ pump (Yvc1p) expressed, we saw an expulsion of intracellular Ca2+ through Yvc1p when the cell is under hyperosmotic shock [19]. This resulted in a rise in cytosolic Ca2+ in cells with an intact Yvc1p and is missing in cells lacking that transporter (data not shown). We found that the Yvc1 knockout construct was not integrating properly into the genome of cmd1-6 ΔYvc1 and Δcmk2 ΔYvc1 cells and we were not able to isolate double knockouts of these genes, implying that in the BY4742 background cmd1-6 Yvc1 and cmk2 Yvc1 are possible synthetic lethal combinations.
Cmk1p inhibits the action of Yvc1p
The cells were pulsed with 100 mM (final concentration) CaCl2 at 30 seconds into the assay, and the cytosolic aequorin luminescence was measured for 13 minutes. BY4742 ΔYvc1 shows an initial spike in cytosolic Ca2+ followed by a return to approximately pre-pulse baseline, which is maintained for the remainder of the thirteen minute assay (Figure 2A), similar to wild type BY4742 cells (Figure 1A) and not statistically significantly different from the wild type in the ratio comparison analysis (P=0.3).
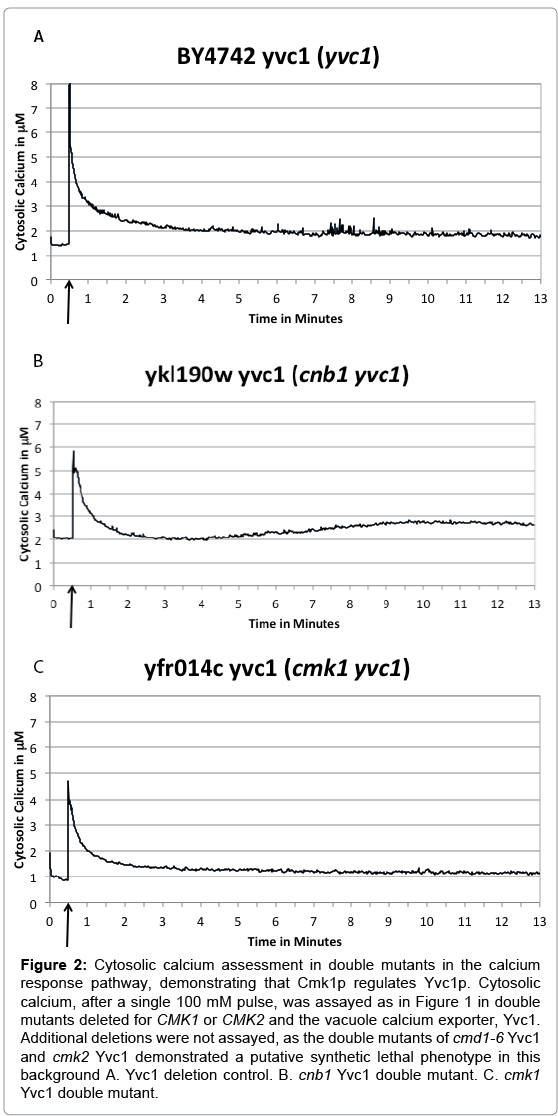
Figure 2: Cytosolic calcium assessment in double mutants in the calcium response pathway, demonstrating that Cmk1p regulates Yvc1p. Cytosolic calcium, after a single 100 mM pulse, was assayed as in Figure 1 in double mutants deleted for CMK1 or CMK2 and the vacuole calcium exporter, Yvc1. Additional deletions were not assayed, as the double mutants of cmd1-6 Yvc1 and cmk2 Yvc1 demonstrated a putative synthetic lethal phenotype in this background A. Yvc1 deletion control. B. cnb1 Yvc1 double mutant. C. cmk1 Yvc1 double mutant.
The double mutants demonstrated different phenotypes, however. The Δcnb1 ΔYvc1 strain (Figure 2B) showed a leak in cytosolic calcium at approximately six minutes (which was significantly different than wild type (Figure 1A) and the Yvc1 single mutant (Figure 2A) in the ratio analysis, P<0.01). This behavior is similar to what was observed in the Δcnb1 single mutant (Figure 1C) (and not statistically significant from the Δcnb1 single mutant in the ratio analysis), indicating that the calcium leak in the cnb1 mutant is not arising from the vacuole through Yvc1p. The Δcmk1 ΔYvc1 strain (Figure 2C) showed the absence of a leak in cytosolic calcium throughout the thirteen minute assay, which was significantly statistically different from the single Δcmk1 mutant (Figure 1E) in the ratio analysis P< 0.05. This behavior is similar to that of BY4742 ΔYvc1 (Figure 2A) and in contrast to what was observed in the Δcmk1 single mutant (Figure 1E). This data is consistent with the second calcium rise coming from the vacuole in the cmk1 mutant. We hypothesize that Cmk1p is regulating Yvc1p either directly or indirectly, and in the absence of that kinase, Yvc1p releases calcium in response to the hyperosmotic changes in the cells after 100 mM calcium treatment, much in the same way Yvc1p releases calcium in response to sodium [19].
Determination of the persistent calcium leak in CNB1 mutants
We continued to explore the origin of the calcium leak in the calcineurin mutant strain. We then hypothesized that either calcium is leaking in through the plasma membrane in calcineurin mutants or is leaking through Vcx1p, a calcium/proton exchanger in the vacuole previously shown to be regulated by calcineurin [23], functioning in reverse to extrude calcium from the vacuole.
We inhibited calcineurin in a wild type and ΔVcx1 strain using FK506 and probed for the calcium leak. To inhibit calcineurin, cells were treated with 4 μg ml-1 FK506. As before, the cells were pulsed with extracellular 100 mM (final concentration) CaCl2 at 30 seconds into the assay, and the cytosolic aequorin luminescence was followed for 13 minutes. BY4742 exhibited an initial increase of cytosolic calcium; a return to approximate pre-pulse baseline level; and then followed by a secondary rise at approximately six minutes (Figure 3A) similar to the cnb1 mutant strain (Figure 1C). This indicates successful inhibition of calcineurin by the inhibitor FK506. Assaying a Vcx1 deletion strain, Ydl128w ΔVcx1, with no treatment served as the control, and the cells showed an immediate rise in cytosolic calcium followed by a return to approximate pre-pulse baseline and then maintained this behavior for the 13 minute timeframe (Figure 3B). However, ydl128w ΔVcx1 treated with FK506 before the assay exhibited a secondary rise at approximately six minutes as in the wild type treated with FK506, indicating the leak is not caused by the reversal of the vacuole H+/Ca2+ exchanger, as the Vcx1p protein is missing in this strain. As in the analysis of Figure 1 and 2, we analyzed the replicates by calculating a ratio between calcium concentration at 4.5 minutes/11 minutes and comparing this ratio to the wild type ratio utilizing an unpaired T-test (equal variance, two tailed), to determine if the rise were statistically significantly different than the change in wild type (Figure 1A). There is no difference between wild type cells and cells lacking Vcx1 in this ratio (Figure 3A) (P=0.2). However, comparing wild type cells versus cells treated with FK506 (Figures 3B and 3C), there is a statistically significant difference in the ratio (P<0.01) indicating FK506 treatment of cells induces the secondary calcium rise. We thus ruled out the possibility of the vacuole H+/Ca2+ exchanger, Vcx1, running backwards in the calcineurin mutants by inhibiting calcineurin in a Vcx1 strain before commencing the cytosolic aequorin luminescence assay and demonstrating that the rise still occurs in this strain with Vcx1p missing and calcineurin inhibited.
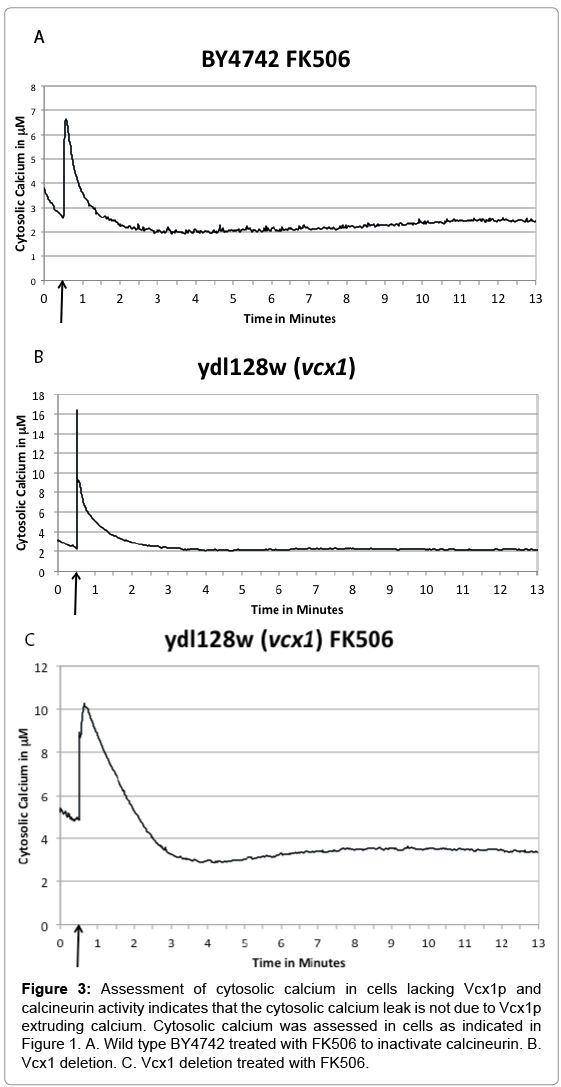
Figure 3: Assessment of cytosolic calcium in cells lacking Vcx1p and calcineurin activity indicates that the cytosolic calcium leak is not due to Vcx1p extruding calcium. Cytosolic calcium was assessed in cells as indicated in Figure 1. A. Wild type BY4742 treated with FK506 to inactivate calcineurin. B. Vcx1 deletion. C. Vcx1 deletion treated with FK506.
We have demonstrated a unique cytosolic calcium increase in select mutants of the calcium homeostasis response. As can be seen in Figure 1, the cytosolic calcium leak occurred at approximately six minutes. We hypothesize that the time delay in the calmodulin, calcineurin and crz1 mutants can be explained by the fact that induced gene expression from the activation of calcineurin to dephosphorylate Crz1p by Ca2+- calmodulin is part of the secondary feedback response in returning the cell to homeostasis after a large increase in cytosolic calcium [4,15]. Gene expression is a slower regulatory process. The behavior of the mutant strains show the cell’s initial feedback response is sufficient to maintain homeostasis for approximately six minutes, while gene expression is needed to maintain the lower cytosolic Ca2+ levels for longer duration.
Our additional collective work demonstrates a novel regulation of Yvc1p by a kinase, calmodulin kinase 1 (Cmk1p). We show here that CaCl2 treatment of the double mutant, Δcmk1 ΔYvc1, eliminates the Ca2+ leak that occurs at approximately six minutes after the CaCl2 treatment of the single mutant Δcmk1; therefore, the deletion of Yvc1 stops the leak (compare Figures 1E and 2C). Based upon these results, we conclude that cells with a deletion in cmk1 are leaking calcium through Yvc1p. Furthermore, we conclude that in the return to homeostasis after an extracellular calcium shock, Cmk1p is a negative regulator of Yvc1p. Intriguingly, Yvc1p has a phosphorylation site in its C terminus, T636 [24] and both the N and C terminus of Yvc1p are hypothesized to be exposed to the cytosol [25]. Perhaps the residual calcium in the cytosol after the extracellular calcium pulse, or even the pulse and calcium wave itself, activate Yvc1p, as calcium has been shown to activate Yvc1p at milimolar concentrations [26]. We hypothesize Cmk1p functions to inhibit this calcium leak, as the cells have no need for a second wave of calcium signaling after such a large transient increase has activated calmodulin and downstream effectors.
We hypothesize that calcineurin, either directly or indirectly, regulate a plasma membrane calcium channel. This activity would serve as a feedback inhibition loop to halt calcium from entering the cell after the initial cytosolic calcium increase. Indeed, in our assay, the extracellular calcium after the transient increase is still close to 100 mM in concentration (data not shown) and the cells would need a mechanism to stop any more calcium from being transported into the cell. Following the CaCl2 treatment of Δcnb1 ΔYvc1 cells, the leak in cytosolic Ca2+ was seen just as occurs in the Δcnb1 single mutant (compare Figures 1C and 2B). This indicates that in the Δcnb1 single mutant, Ca2+ does not leak through Yvc1p and could be leaking through an unidentified plasma membrane channel. Intriguingly, yeast cells lacking the CNB1 gene have a higher than wild type cytosolic calcium level after growth in rich media [27], lending credence to this hypothesis.
Calcineurin is a known inhibitor of Vcx1p [23] and Bagar and Bencina [28] demonstrated elevated levels of cytosolic Ca2+ induce an increase in intracellular acidity, indicating additional potential roles for the vacuole in calcium homeostasis in addition to calcium sequestration and extrusion through Yvc1p; thus, we wondered if, in the absence of its regulator calcineurin, Vcx1p were pumping calcium out of the vacuole under the stress conditions after 100mM extracellular calcium treatment, (Figure 1C). Therefore, we tested the calcium phenotype in a Vcx1 deletion treated with FK506 to inhibit calcineurin (Figure 3C). We ruled out the possibility that Ca2+ is leaking from the vacuole through a reverse action via the H+/Ca2+ exchanger (Vcx1p) when Cnb1p is inhibited. As can be seen in Figure 3C, the inhibition of CNB1 coupled with the deletion of Vcx1 does not eliminate the cytosolic calcium leak seen after 6 minutes.
We have demonstrated a novel behavior, an additional cytosolic calcium increase at 6 minutes, after a 100 mM calcium pulse in a subset of deletion mutants whose proteins have been implicated in the calcium homeostasis pathway. Interestingly, the leak comes from the vacuole in cmk1 mutants but not from the vacuole in cnb1 mutants. Thus we have shown two separate phenomenon in our deletion mutant analysis that seems to demonstrate locus heterogeneity. We have shown that Yvc1p, the yeast vacuolar calcium exporter, is regulated by calmodulin kinase 1 (Cmk1p) and that Yvc1p is not controlled by calcineurin. We also have demonstrated that in the absence of its calcineurin, Vcx1p does not function in reverse to extrude calcium into the cytosol. We will continue studying the phenomenon of the secondary cytosolic calcium increase in order to determine additional mechanisms of regulation.
We thank Eden Tanzosh, Belinda Miguel, Jane Kruchowsky, and Lorenzo Sanchez for excellent technical help. This work was in part supported by a grant to PAM from the New College of Interdisciplinary Arts and Sciences. We give special appreciation to Dr. David Bedwell (University of Alabama), Dr. Martha Cyert (Stanford), Dr. Trisha Davis (University of Washington), and Dr. Yoshiro Saimi (University of Wisconsin, Madison) for plasmids and/or strains. JK wishes to thank Dr. Becky Ball (ASU) for help with her Honors thesis, from which this paper arose. We also thank Dr. William Courchesne (University of Nevada Reno) for his help with the thesis and the suggestion of the FK506 experiments.