Journal of Osteoporosis and Physical Activity
Open Access
ISSN: 2329-9509
ISSN: 2329-9509
Review Article - (2015) Volume 3, Issue 3
Osteoporosis is a chronic, systemic, metabolic disease characterized by loss of bone mass, disruption of the micro architectural structure of bone tissue and changes in the physical properties of bone matrix that leads to an increased risk of fracture. The rise in average age in western society increases the need for novel therapeutic strategies for this disease; therefore, in order to identify novel drugs, systems for in vitro screening for molecules capable of stimulating osteoblast differentiation are of increasing relevance. Indeed, detailed analysis and characterization in cellular and animal models are necessary preclinical steps towards the development of novel therapeutic options.
Here we present a review of cellular models for osteoblast differentiation and animal models for osteoporosis as well as available methodologies to evaluate bone parameters in vivo. These models are useful to identify novel targets and to develop safer and more effective therapeutic strategies with new drugs able to reduce fractures risk in human.
<Keywords: Osteoporosis; Animal model; Cellular models; New drugs
BMD: Bone Mineral Density; SERMs: Selective Estrogen Receptor Modulators; rhPTH: Recombinant Human Parathyroid Hormone; RANKL: Receptor Activator of Nuclear Factor κ-B Ligand; MSCs: Mesenchymal Stem Cells; CFU-f: COLONYForming Unit-Fibroblasts; HSCs: Hematopoietic Stem Cells; BMMSCs: Bone Marrow-Derived MSCs; DEXA: Dual-Energy X-ray Absorptiometry
Osteoporosis is a chronic, systemic, metabolic disease characterized by loss of bone mass, disruption of the micro architectural structure of bone tissue and changes in the physical properties of bone matrix that leads to an increased risk of fractures. This condition has a multifactorial etiology and it can be primary (postmenopausal and senile) or secondary; the underlying mechanism in all cases is an imbalance between bone resorption by osteoclasts and bone formation by osteoblasts that leads to a decreased bone mineral density (BMD). Indeed, bone is a dynamic tissue that undergoes a continuous physiological process of remodeling [1]. Bone resorption occurs throughout life and in normal conditions is counterbalanced by bone formation, in order to maintain a balance in bone mass. Alterations in bone remodeling favoring bone resorption over de novo synthesis cause diseases such as osteoporosis; conversely, a deficit in osteoclast activity and, therefore, in bone resorption, leads to a pathological condition known as osteopetrosis.
Osteoporosis is a multifactorial disease and its clinical manifestation depends upon the complex interrelation between environmental and genetic factors [2-4]. Risk factors for osteoporosis development include positive family history, age, female gender, hormonal status, long termuse of medications such as glucocorticoid, and lifestyle (diet, physical activity, vitamin D deficiency, smoke and alcohol abuse) [5,6]. The disorder can be localized or involve the entire skeleton with different consequences for patients’ health and healthcare costs; in fact, vertebral compression fractures may have a lower economic impact compared to hip fractures, but they have a substantial negative effect on the patient’s function and quality of life [7].
The increase in lifespan in western society has led to a consequent increase of osteoporosis substantially decreasing quality of life and increasing healthcare costs and the need for additional therapeutic strategies. Even obesity, a major health issue in industrialized countries, has recently been associated to deteriorated bone microarchitecture and increased prevalence of vertebral fractures [8].
Currently approved therapeutic options to treat osteoporosis are several and include the estrogen replacement therapy, the use of selective estrogen receptor modulators (SERMs) such as raloxifene, the bisphosphonates (e.g., alendronate, risedronate, ibandronate, and zoledronic acid), calcitonin, recombinant human parathyroid hormone (rhPTH) and its derivatives (e.g., teriparatide), the monoclonal antibody denosumab that binds the receptor activator of nuclear factor κ-B ligand (RANKL), and strontium ranelate (not approved in the United States) [9,10].
However, there are particularly challenging categories of osteoporotic patients that show an inadequate response to therapy and patients with pre-existing conditions, such as renal or gastrointestinal diseases, that may not tolerate the existing therapies [11].
Indeed, most of the drugs currently available present contraindications and even severe side effects. For example, biphosphonates, that represent often the first-choice therapy for osteoporosis, although effective in reducing bone loss and vertebral fracture risk and generally well tolerated, are contraindicated in patients with renal impairment and gastrointestinal problems and present several short and long-term safety concerns, including two rare but severe adverse effects, the osteonecrosis of the jaw and atypical femoral fractures [11-13]. Moreover, severe hypocalcemia has been reported in a number of patients treated with the recently developed anti-resorptive drug denosumab [14].
Another important aspect that point out the need to develop novel drugs for the treatment of osteoporosis is that most of the currently available therapeutic options are represented by anti-resorptive agents, with the exception of rhPTH, that currently represents the only approved osteoanabolic drug, and few other still in development but promising anabolic agents, such as anti-sclerostin antibodies [15]. Indeed, anabolic drugs stimulating bone formation are useful not only to reduce both vertebral and non-vertebral fracture risk but also to improve bone parameters, such as bone microarchitecture and connectivity, whereas anti-resorptive drugs are not effective in decreasing non-vertebral fractures and their use leads to a reduced bone turnover and to an impaired bone remodeling [16,17].
Therefore, it is of paramount interest the identification of novel safer and more effective drugs capable of inducing bone synthesis. At this aim cellular and animal models are essential in order to maximize the efficacy of drug identification process.
Here we describe and analyze the currently available cellular models for osteoblast differentiation, represented by mesenchymal stem cells (MSCs) and derived cell lines, useful to identify new targets and to develop and test the efficacy of novel anabolic drugs for the treatment of osteoporosis. We also provide an overview of the most used rodent models of osteoporosis, essential to validate the efficacy of newly developed drugs in vivo as well as to assess their safety profile. Other animal models, such as non-human primates useful to evaluate the risk of vertebral fractures, will not be described in the present review. Finally, we present the methods that allow to evaluate the bone mass, the microarchitecture and the skeleton metabolism in animal models, underlying the differences, the strengths and the limits of the different methodologies.
MSCs are adult, multi-potent stem cells able to self-renew and generate, after appropriate stimuli, several mesodermal cell lineages, including adipocyte, osteoblasts, chondrocytes, myoblasts, tenocytes and haematopoiesis-supporting stroma [18,19]. MSCs were identified by Alexander Friedenstein and co-workers in the 1960s. The investigators plated bone marrow cells at low density and identified a population of fibroblastoid, clonogenic plastic-adherent cells they named colony-forming unit-fibroblasts (CFU-f). Each colony, seeded under the kidney capsule of mice, was able to give rise in few weeks to fibrous tissue, bone and bone marrow; bone cells were of donor origin, whereas the hematopoietic tissue was of recipient origin, probably due to the homing of host circulating blood hematopoietic stem cells (HSCs) [20,21]. These data showed that in the bone marrow there is a population of cells distinct from hematopoietic precursors, able to generate bone tissue and to provide an appropriate microenvironment for HSCs homing and hematopoiesis. In the subsequent years, further studies demonstrated that the inherently bone-forming cells discovered by Friedenstein and named “osteogenic stem cells” were actually multipotent and capable of generating not only osteoblasts, but also adipocytes, condrocytes and myoblasts [18,22].
In the last few years, several studies have shown that MSCs could exhibit a broad degree of plasticity, since they seem to be able to differentiate also into non-mesodermal cell types, including neural cell lineages, endothelial cells, hepatocytes and pancreatic progenitor cells [23-26]. However, there are still controversies regarding the true plasticity of MSCs; in fact, differentiations observed in vivo could be due to the fusion of MSCs with endogenous cells, whereas in vitro nonstandardized culture conditions may lead to the selection of rare cell populations with greater differentiation potential. Furthermore, recent reports have shown that, at least in some circumstances, the ability of MSCs to promote in vivo tissue regeneration in response to disease or injury is unlikely due to their ability to differentiate, since they exhibit very low or transient levels of engraftment [27]. These observations suggest that MSCs could promote tissue repair through the secretion of molecules that modify tissue microenvironment and stimulate regeneration through resident cells. Indeed, several studies showed that MSCs produce a variety of cytokines and adhesion molecules that promote endogenous cell survival and proliferation, induce angiogenesis and have immune-modulatory and anti-inflammatory effects [28-31].
Although MSCs were originally isolated from bone marrow, it has soon become clear that these cells are present in different tissues, including adipose tissue, peripheral blood, skeletal muscle, tendon and several fetal tissues [32-37]. However, several studies have reported that MSCs isolated from different sources are not equivalent and that show differences in many aspects, including proliferation rate, differentiation potential and expression of specific markers [38]. Therefore, further studies are required to better characterize MSC populations from different sources in order to be able to select the most suitable ones for the specific purpose of the study as well as for selected clinical applications.
During the last years several techniques for MSCs isolation and culture-expansion have been developed and MSCs have been successfully harvested from several tissues of multiple species; so far, bone marrowderived MSCs (BM-MSCs) are the most used cells for clinical applications and, therefore, the most intensely studied. The conventional isolation protocol, widely used for the purification of BM-MSCs, is based on the ability of these cells to adhere to tissue culture plates [39]. However, in the last years, great efforts have been made to characterize MSCs immunophenotype in order to identify a surface antigen panel that could be useful for their isolation and purification [40,41].
The definition of MSCs is extremely complicated as such cells are morphologically, phenotypically and functionally heterogeneous. For this reason, minimal criteria for the definition of human MSCs have been established: MSCs have to be plastic-adherent, express CD105, CD73 and CD90, and lack expression of CD45, CD34, CD14 or CD11b, CD79alpha or CD19 and HLA-DR surface molecules and, finally, they must be able to differentiate at least into osteoblasts, adipocytes and chondroblasts in vitro [42].
MSCs can be successfully isolated from several tissues and organs of humans and animals [18,39]. However, although primary cultures present several advantages compared to cell lines, since they keep the distinctive features of the native organ or tissue and reflect better the biochemical activities of cells in vivo, some experimental procedures require a greater standardization and reproducibility of the results and the possibility to extensively expand the cells in an undifferentiated state. Furthermore, primary cultures often represent a highly heterogenous population composed by osteoblastic cells at different stages of differentiation and cells of other lineages, such as fibroblasts [43]. For these reasons, cell lines with mesenchymal characteristics have been established, including non-transformed, experimentally immortalized and osteosarcoma-derived cell lines.
Non-transformed cell lines
Mouse cell lines, such as W20-17, C3H10T1/2 and MC3T3-E1 cells, are widely used for in vitro screening and research. The murine stromal cell line W20-17 was obtained from mouse bone marrow using a limited dilution method. Bone morphogenetic protein 2 (BMP-2) is able to increase alkaline phosphatase expression in W20-17 cells in a dose-responsive manner without affecting cellular proliferation, so this cell line was initially developed and used to evaluate the effect of recombinant human BMP-2 (rhBMP-2) on osteogenic differentiation [44]. W20-17 cells are also capable of differentiating into osteoblasts when cultured in a conditioning medium supplemented with the synthetic glucocorticoid dexamethasone, L-ascorbic acid-2-phosphate and sodium β-glycerophosphate (Figure 1, panels A and B) as well as of giving rise with great efficiency to adipocytes when treated with insulin, dexamethasone, indomethacin and 3-isobutyl-1-methylanxthine (unpublished data, Figure 1, panels C and D) [45].

Figure 1: Osteogenic and adipogenic differentiation of W20-17 cell line. Authors’ unpublished images.
W20-17 cells were culture in regular (A) and osteogenic medium (B). After 21 days cells were stained with the calcium-binding dye Alizarin Red S to detect mineralized nodules of bone matrix.
W20-17 cells treated for 2 weeks with regular (C) and adipogenic medium (D) were stained with Oil Red O to visualize lipid droplets.
The other cell line commonly used, namely C3H10T1/2, is a murine embryonic fibroblastic cell line extremely sensitive to post confluence inhibition of cell division, originally established by Reznikoff and colleagues from C3H mouse embryos [46]. This cell line is able to differentiate into osteoblasts, adipocytes and chondrocytes [47-49]. MC3T3-E1 cells, instead, are pre-osteoblasts derived from newborn mouse calvaria [50]. Different sub-clones were isolated from the MC3T3-E1 cell line, selected for high or low osteoblast differentiation and mineralization in presence of ascorbic acid. Highly differentiating subclones express several osteogenic markers (bone sialoproteins, osteocalcin, and the PTH receptor) and are able to induce the formation of bone-like ossicles resembling woven bone in immunodeficient mice [51].
In addition, there are several other murine cell lines that, under appropriate stimuli, can be induced to differentiate into osteoblasts in vitro. For instance, the mouse myoblast cells C2C12 differentiate along the osteogenic lineage in presence of BMPs and the embryo-derived fibroblasts NIH3T3 adopt an osteoblast-like phenotype when treated with 1α,25-dihydroxyvitamin D(3) and dexamethasone [52-54].
Another non transformed cell line commonly used is represented by the UMR-201 cells established from neonatal rat calvaria; this clonal cell line has a limited lifespan (12 passages in culture), exhibits a pre-osteoblast phenotype, an alkaline phosphatase (AP) activity undetectable in basal condition and significantly prompted by the treatment with retinoic acid [55].
Experimentally immortalized cell lines
Non transformed cell lines have a limited lifespan in culture and this could represent an obstacle for long-term projects or studies in which large amounts of cells are needed. In order to overcome this issue, experimentally immortalized and osteosarcoma-derived cell lines with osteoblast features have been developed. Nonetheless, it is important to consider that, although a valuable tool, proliferation in these cell lines is non-physiological as several mechanisms, included senescence and contact inhibition, are disrupted.
KS-4 clonal cell line was obtained from fetal mouse calvaria through the immortalization with the c-HA-ras-1 proto-oncogene. These cells produce type-1 collagen, present increasing AP activity when approaching confluence, form mineralized nodules without addition of β-glycerophosphate and stimulate osteoclastogenesis in co-culture with spleen cells [56]. The same group subsequently established from KS-4 cells three sub-clones which represent phenotypically different osteoblasts, but able to support osteoclast differentiation to the same extent as the parental cell line [57].
Also few human immortalized mesenchymal cell lines have been developed, including the adult human osteoblast-like (hOB) cell line, the human fetal osteoblast (hFOB) cells and the HM3. B10 cell line. hOB cells were established from normal osteoblastlike cells isolated from a 68-year old woman and immortalized with a plasmid encoding for SV40 small and large T antigens. The cells express alpha(I)-pro-collagen, osteopontin 1a, transforming growth factor beta and interleukin-1 beta mRNAs, present functional estrogen and androgen receptors, produce calcified deposits when treated with β-glycerophosphate and, therefore, represent a welldifferentiated cell line with the phenotype of mature osteoblasts [58]. The hFOB cell line was derived through the immortalization of primary fetal tissues cultures from a spontaneous miscarriage with a gene coding for a temperature-sensitive mutant (tsA58) of SV40 large T antigen. Differentiated cells express high levels of AP, type I collagen, osteopontin, osteonectin and bone sialoprotein, and form mineralized nodules [59]. The HM3.B10 cell line was obtained through the immortalization of primary cell cultures of fetal human BM-MSCs with a v-myc-encoding retroviral vector. The authors have demonstrated that this cell line is able to differentiate not only into mesoderm-derived lineages (osteoblasts, adipocytes and chondrocytes), but also into neural cell types [60]. More recently an additional human mesenchymal cell line derived from amniotic fluid with interesting biological properties and differentiation potential has been developed and characterized. These cells, called mesenchymal progenitor (MePr) cells, were obtained through the immortalization of human MSCs from amniotic fluid with lentiviral vectors encoding human papillomavirus type 16 genes (HPV16-) E6/E7 and human telomerase reverse transcriptase (hTERT) and are able to differentiate into chondrocytes, adipocytes, osteoblasts and neuronal cells [61].
Osteosarcoma-derived cell lines
Several osteosarcoma-derived cell lines have been developed and characterized and they could represent a valuable tool to investigate the molecular mechanisms altered in cancer as well as to identify targets and test novel drugs for therapy.
ROS clonal cell lines were established from a spontaneous transplantable rat osteosarcoma on the basis of PTH-responsiveness. These cells express high AP levels, produce bone-matrix proteins and mineralized tumors in host rats [62]. UMR-106 is a cell line derived from a 32P-orthophosphate-induced transplantable rat osteosarcoma that has been extensively studied; the cells express high AP levels and are responsive to PTH, prostaglandins and 1α,25-dihydroxyvitamin D(3) [63-65].
Also few human osteosarcoma cell lines have been isolated; the most widely used ones are represented by the well-characterized Saos-2 and MG-63 cell lines. Saos-2 cell line was established from a 11-year old Caucasian female patient and exhibit a mature osteoblast phenotype. The cells express high AP levels, that increase with time in culture and cell density, are not tumorigenic when injected in immunosuppressed mice, are able to form a calcified matrix in vivo and express a panel of cytokines and growth factors highly similar to primary human osteoblastic cells [66,67]. MG-63 cells were isolated from the juxtacortical osteosarcoma of a 14-year old Caucasian male and were considered a promising source for interferon production [68]. These cells show an immature osteoblast phenotype, form cell aggregates in culture since they lack contact inhibition, and are responsive to 1α,25-dihydroxyvitamin D(3) that is able to modulate cell phenotype and morphology [69,70]. Recently, Di Fiore and colleagues selected from MG-63 cells by long-term treatment with 3-aminobenzamide (3AB) a new stable and heterogeneous cell population named 3ABOS, that present features such as self-renewal, pluripotency in vitro and tumorigenicity in vivo characteristic of cancer stem cells (CSCs) [71-73]. The potential molecular mechanism underneath the de-differentiation of 3AB-OS compared to the parental cell line and the acquisition of stem cell-like properties has been identified in a gain of function mutation in the tumor-suppressor gene p53 [74]. The isolation and the molecular and phenotypical characterization of CSCs has important implications in oncologic research, since CSCs are believed to be responsible for initiating and maintaining tumors [75]; therefore CSCs-like cell lines, such as 3AB-OS, could be of paramount importance to get insight into the biology of the “cancer-initiating cells” and to provide targets for the development of more effective therapeutic strategies.
MSCs are considered a promising source for cell-based therapy and regenerative medicine, since they can be extensively expanded in vitro retaining their undifferentiated phenotype and differentiation potential [76], they do not cause teratoma formation in vivo and are well-tolerated in clinical trials. Moreover, they show unique features that largely contribute to their attractiveness: MSCs are immuneprivileged, have the ability to migrate to injured tissues after systemic administration, produce several trophic factors and are able to regulate inflammatory and immune responses [30,31,77,78]. For these reasons, MSCs were largely employed in preclinical and clinical studies to treat a broad spectrum of diseases, including auto-immune and inflammatory diseases, graft versus host disease (GvHD), myocardial infarction and spinal cord injuries [79]. The MSCs potential to differentiate into several cell types has opened a variety of tissue engineering applications: undifferentiated or differentiated MSCs seeded into a variety of natural or synthetic biomaterial scaffolds have been successfully applied in repair of cartilage and bone defects [79-81].
Most interestingly, MSCs and their derived cell lines represent a valid in vitro differentiation system and a valuable tool for the identification of molecular mechanisms underlying cellular differentiation: the identification of genes that regulate osteoblast differentiation can directly provide new pharmacological targets for the induction of bone neo-synthesis and, therefore, the treatment of osteoporosis. Osteogenesis is a complex and not entirely elucidated biological process regulated by intrinsic cellular signals and extrinsic micro-environmental stimuli from the surrounding stem cell-niche. Cells differentiate in a multi-step process in which the progression from one stage to the following is regulated by a complex network of growth factors, signaling molecules and hormones, and it is characterized by the activation and subsequent inactivation of transcription factors and the expression of bone specific marker genes [82].
In order to induce osteogenic differentiation in primary MSCs and derived cell lines it is necessary to reproduce the environmental stimuli that drive the process in vivo. The most common supplements used to induce osteoblastogenesis in cultured cells are represented by a combination of dexamethasone, L-ascorbic acid-2-phosphate and sodium β-glycerophosphate, but also additional or alternative supplements, such as 1α,25-dihydroxyvitamin D(3) and rhBMPs, have been successfully used [83]. BMP-4, for example, is able to stimulate osteogenic differentiation in cell culture systems and also in vivo, as shown in Figure 2 (panels B and D). Furthermore, additional methods to stimulate osteogenic differentiation have been investigated, including the use of various bio-polymeric scaffolds, mechanical stress or hydrostatic pressure, alone or in combination with osteogenic supplements [82].
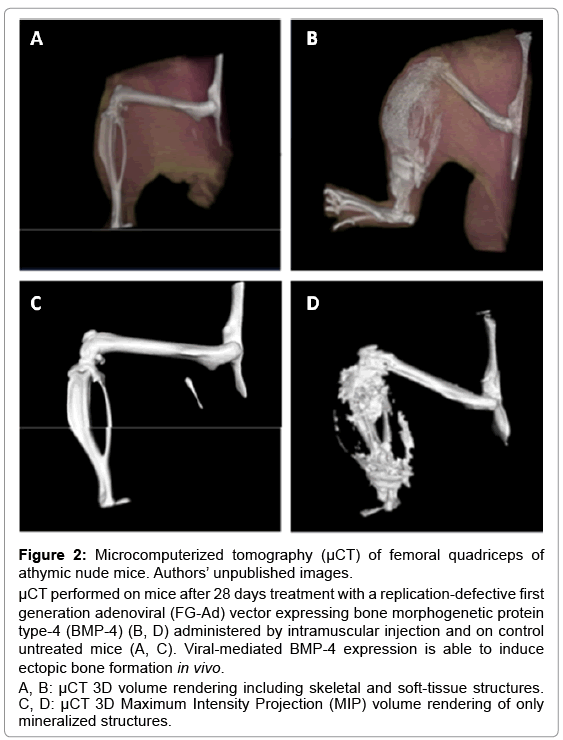
Figure 2: Microcomputerized tomography (μCT) of femoral quadriceps of athymic nude mice. Authors’ unpublished images.
μCT performed on mice after 28 days treatment with a replication-defective first generation adenoviral (FG-Ad) vector expressing bone morphogenetic protein type-4 (BMP-4) (B, D) administered by intramuscular injection and on control untreated mice (A, C). Viral-mediated BMP-4 expression is able to induce ectopic bone formation in vivo.
A, B: μCT 3D volume rendering including skeletal and soft-tissue structures. C, D: μCT 3D Maximum Intensity Projection (MIP) volume rendering of only mineralized structures.
More recently a number of automated procedures for cell culture and manipulation have been developed (high-throughput screenings, HTS) making it possible to conduct a large number of tests with reduced manpower and more affordable costs. These screenings allow the identification of genes involved in a number of biological processes or compounds able to alter them. Recently, RNA interference-based approaches have become particularly popular for their ability to silence specific genes and, therefore, study the effect of the absence of a single gene product in definite biological phenomena, such as embryonic stem cells self-renewal or differentiation [84,85]. These studies can be performed using chemically synthesized RNA molecules or with DNAbased approaches, using plasmids, lentiviruses or retroviruses encoding short hairpin RNAs (shRNAs), that allow to achieve the sustained gene silencing necessary for long-term studies. A typical strategy that we have recently used for a HTS in W20-17 cell lines is outlined in Figure 3 [45]. In addition, HTS have been used to screen compounds libraries in order to directly identify chemical compounds able to induce or block a biological process in order to discover potential drugs [86-88]. Genes or molecules identified with HTS represent only the first step towards the identification of a therapeutic compound. In fact, HTS results need a thorough in vitro validation in order to characterize their actual mechanism of action and, subsequently, a validation in animal models.
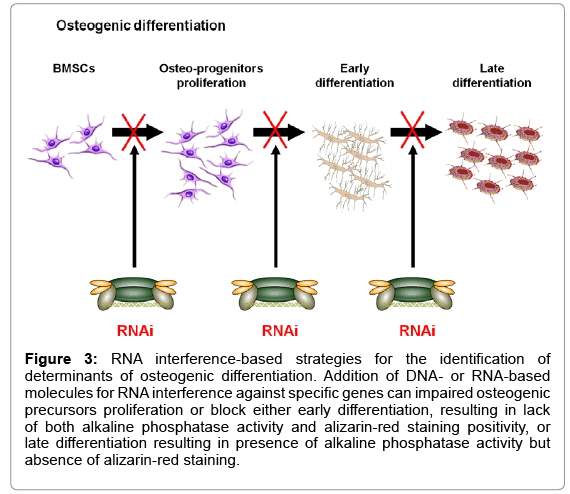
Figure 3: RNA interference-based strategies for the identification of determinants of osteogenic differentiation. Addition of DNA- or RNA-based molecules for RNA interference against specific genes can impaired osteogenic precursors proliferation or block either early differentiation, resulting in lack of both alkaline phosphatase activity and alizarin-red staining positivity, or late differentiation resulting in presence of alkaline phosphatase activity but absence of alizarin-red staining.
An appropriate animal model for research should be based on the following considerations: appropriateness, possible genetic similarity, transferability of the information obtained to human conditions, genetic uniformity when possible, deep background knowledge of biology and genetics, and possibly low cost and wide availability [89].
The most commonly used animal model for osteoporosis studies is represented by rodents [90], in particular rats and mice. Their use in research is advantageous since they are inexpensive to maintain, grow rapidly, have relatively short lifespan and are widely available. The ovariectomized (OVX) rat is the most frequently used animal model because of its ability to successfully mimic cancellous and endocortical bone changes observed in women after menopause [91]; however, the molecular mechanisms of bone disorders, such as osteoporosis, have been investigate predominantly in mice because of their large use in gene manipulation technology and their numerous inbred strains varying in peak bone mass.
Despite their unquestionable advantages, rodent animal models present also some limitations; for instance they are quadrupeds and therefore do not allow an accurate investigation of spinal osteoporosis and the risk of vertebral fractures, as spinal lesions are less evident in absence of bipedal posture. Nevertheless, these models are considered extremely reliable for the experimental evaluation of the effect of potential drugs.
Animal models of osteoporosis can be classified in two categories: models induced by a number of procedures, such as ovariectomy, immobilization and drugs administration, and genetically determined models, represented by transgenic and knockout animals.
The OVX rat is the animal model commonly used to evaluate skeletal response to estrogen depletion and postmenopausal osteoporosis. In this model, after an initial rapid loss of cancellous bone mass and strength, a steady state phase of bone mass is reached, featured by a generally increase of bone turnover [92]. Since this model has no naturally occurring fragility fractures associated with the osteopenia, various bones such as proximal femur, vertebral body and femoral shaft are evaluated by mechanical testing. The OVX mice, similarly to rats, exhibit bone changes that mimic those occurring in postmenopausal women, although the magnitude of response of both cancellous and cortical bone to OVX is dependent on mouse strain and skeletal site [93]. Despite the fact that OVX rat represents the gold standard of animal models for postmenopausal osteoporosis, mice can be particularly useful as an initial in vivo evaluation of new drugs for osteoporosis, since much less drug is needed and experimental procedures have been developed more extensively.
Another method used to induce osteoporosis is represented by immobilization, which can be either surgical such as nerve, tendon and spinal cord resection, or conservative such as casting, bandaging of one limb or suspension of both hind limbs [92]. In the immobilization model, the bulk of bone loss occurs in the hind limbs, sites of the greatest mechanical loading, but in general, the rate of bone loss is faster in cancellous than in cortical bone, probably due to the surface to volume ratio, which is increased in cancellous bone [94]. In this model, in contrast to the ovariectomized model, periosteal bone formation ceases in cortical bone, but endosteal resorption, leading to slow bone loss, continues. Moreover, one of advantages of this model is that bone changes also occur in the distal tibial metaphysic whose architecture is similar to that of human adults [95].
Even administration of several drugs is known to induce osteoporosis; indeed, glucocorticoid administration to mice is able to alter bone metabolism, as showed by histomorphometric parameters and biochemical markers, and to induce osteopenia/osteoporosis differing from postmenopausal or senile osteoporosis [96]. The glucocorticoid-induced osteoporosis mouse model simulates closely the human bone disease due to glucocorticoid excess, that represents the third most common cause of osteoporosis, and it is characterized by decreased bone formation and turnover, impaired osteoblastogenesis and osteoclastogenesis and a significant reduction in bone density and cancellous bone mass. The decrease of (BMD) and biomechanical competence is even more pronounced when steroid medication is administered to ovarectomized animals [97].
In order to study osteoporosis, bone metabolism and the activity of factors involved, various transgenic and knock-out rodent models have been proposed. Senescence-accelerated mouse-prone 6 (SAMP6) described by Matsushita et al. [98] was the first mouse model of senile osteoporosis with spontaneous fractures at old age and represents the most used mouse model to study senescence-related pathologies, including bone disease and fractures, and their genetic background [99]. The SAMP6 model reproduces many characteristics of human senile osteoporosis and, interestingly, presents spinal alterations making it a suitable model to study vertebral fragility associated with the pathology [100].
Numerous endogenous agents were identified for their involvement in bone metabolism such as the calcium-binding protein regucalcin. Indeed, regucalcin transgenic rats manifest hyperlipidemia and osteoporosis due to increased osteoclastic activity and impaired osteoblast development and bone formation [101].
Gamma-Glutamyltransferase (GGT), an enzyme present primarily in liver, is also involved in bone metabolism. Transgenic mice overexpressing GGT present accelerated osteoclast development and bone resorption; the ability of GGT to stimulate osteoclastogenesis is independent from its enzymatic activity, suggesting a new potential function of GGT as a cytokine [102].
Also factors of immune system were identified for their potential involvement in osteoporosis pathogenesis. Duque et al. investigated the association between bone metabolism and Interferon-γ (IFN-γ) signaling: IFN-γ receptor knockout mice show reduced bone mass and bone turnover and the administration of IFN-γ is effective to improve bone mass in ovariectomized mice [103].
Macrophage migration inhibitory factor (MIF) is a proinflammatory and immunomodulatory cytokine likely involved also in bone metabolism with a not yet elucidated mechanism. In fact, transgenic mice overexpressing MIF develop high-turnover osteoporosis characterized by an increase in both bone resorption and bone formation. The enhanced bone resorption is not due to an increase in osteoclast number but it is probably caused, at least in part, by MIF ability to upregulate the osteoblast secretion of metalloproteinase (MMPs) that favor the process of bone resorption [104].
Furthermore, mouse models of a number of diseases were investigated for their involvement in bone metabolism; for instance diabetes, particularly type 1 Diabetes (T1D), causes frequently skeletal complications including decreased bone mass, osteoporosis and increased fracture risk. Indeed, the T1D mouse model that over-express the inducible nitric oxide synthase under rat insulin promoter control (RIP-iNOS) spontaneously develop osteopenia and could represent an useful model to study and prevent osteopenia/ osteoporosis associated to diabetes [105]. Another cause of secondary osteoporosis is hyperthyroidism [106]; several mouse models with mutations in triiodothyronine (T3) receptor gene have been generated and demonstrated that T3 has an anabolic effect during growth whereas it stimulates bone resorption in adult skeleton [107]. It has been shown that thyroid-stimulating hormone (TSH) also plays a role in bone metabolism; indeed, TSH administration is able to restore bone mass in OVX rodent models [108,109] while TSH receptor (TSHR) knockout mice develop severe osteoporosis [110,111].
The methods used to evaluate bone mass, the architecture and the metabolism in the skeleton in animal models are quite similar to those used for the same purposes in humans; therefore, it is possible to determine biochemical markers in blood and urine, such as calcium, phosphorus, magnesium and specific proteins and compounds released by osteoblasts and osteoclasts during the synthesis and resorption of bone matrix. Markers of bone formation, such as alkaline phosphatase and osteocalcin, and markers of bone resorption, as tartrate-resistant acid phosphatase and urinary type I collagen cross-linked N-telopeptides, can be measured by using several techniques, including luminescence immunoassay, liquid chromatography, radioimmunoassay, enzymelinked immunosorbent assay and immunoradiometric assay [112,113]. Even though these markers are useful to monitor the biological processes underlying osteoporosis, they allow detecting only changes in bone metabolism in the whole skeleton and do not provide information regarding bone mass and strength. In addition, the availability of reagents to detect biochemical markers in animal models is currently limited. Because of these limitations, direct observation of bone is always preferred.
Dual-energy X-ray absorptiometry (DEXA) is often used to evaluate BMD and bone mineral content (BMC) and it allow the evaluation of both total and regional bone density, also in small animals such as rats and mice. Although DEXA provides important information about fractures risk, BMD is not the only determinant of bone strength and other parameters, including connectivity, bone architecture and trabecular volume and spacing, need to be evaluated.
Another method useful for osteoporosis evaluation in small animal models is represented by peripheral quantitative computerized tomography (pQCT) [114]. pQCT is a powerful imaging technique that presents some advantages over DEXA, such as the ability to analyze cancellous and cortical bone separately and to detect earlier changes in bone mass, even though it does not allow to detect structural and morphological changes at the level of an individual trabecula in small rodents.
Micro computerized tomography (μCT), instead, allows to evaluate a bone area as small as an individual trabecula and provides very high resolution images; this technique can evaluate parameters linked to connectivity and elasticity of the bone and allows a tridimensional analysis of bone architecture (Figure 2, panels A-D).
All these noninvasive methods combined allow a complete assessment of bone structure and mass and a continuous follow-up in rodent models avoiding the sacrifice of animals required by histological assessments. Indeed, invasive methods such as histomorphometry, provide higher resolution 2-dimensional images compared to noninvasive methods, but they present also some limitations. In fact, they require the sacrifice of animals and, therefore, the use of a large number of subjects is needed, also in order to overcome inter-subject variability and to obtain statistically significant data. In addition, they allow the evaluation of a restricted bone area that could not be representative of the changes occurring in the whole skeleton.
Nevertheless, histomorphometry represents a valuable technique that allows an accurate study of bone mass and architecture and the evaluation of several parameters, including the number of osteoclasts, osteoclasts and osteocytes, and the trabecular thickness and number.
The above-described animal models of osteoporosis are of paramount relevance for the evaluation of newly developed compounds identified after in vitro HTS or to extend the available knowledge about currently used medications, alone or in combination with other drugs, or the consequences of long-term therapy [115,116]. The development of research centers where it is possible to combine expertise in the analysis of cellular models for HTS and the presence of state of the art equipment and expertise for the evaluation of animal models constitute the basis for the identification of molecular mechanisms of osteoporosis and the consequent identification of novel therapeutic strategies capable of reducing bone loss and fracture risk in human.
Ruosi C, Querques F, Colella G, Liccardo S, Lombardo B and Pastore L declare that they have no conflict of interest.