Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2011) Volume 2, Issue 1
In this study, changes of ionic ratios (Na+ to K+, Na+ to Ca+2, Na+ to Mg+2, K+ to Ca+2, K+ to Mg+2 and Ca+2 to Mg+2) during spawning migration of roach and its effect on sperm motility traits (sperm movement duration, percentage of motile spermatozoa) and sperm production characteristics (spermatocrit, sperm density and sperm volume) were investigated. Sperm of 30 males was collected during the spawning migration (February, March and April). Ionic ratios of Na+ to K+, Na+ to Ca+2 and Ca+2 to Mg+2 significantly changed (P<0.05) between three terms, but Na+ to Mg+2, K+ to Ca+2 and K+ to Mg+2 ratios did not show significant differences (P>0.05) between February, March and April. The results suggested that sperm movement duration is slightly influenced by (P<0.05) Na+ to K+ ratio. A similar tendency was observed in values of spermatocrit and percentage of motile spermatozoa due to Na+ to Ca2+ ratio. Also, Ca+2 to Mg+2 ratio influenced percentage of motile spermatozoa. The present study describes that biochemical composition (Ionic ratios) and semen characteristics will help in development of the basic knowledge and the strategies during artificial spawning programmes in this species.
Keywords: Spermatological properties; seminal plasma; ionic ratio; Roach.
The Roach, Rutilus rutilus caspicus (Teleostei: Cyprinidae) is a migratory fish that reproduces in hatcheries and the fingerlings are released into rivers flowing into the Caspian Sea [1]. This species is found in the southeastern part of the Caspian Sea, i.e. in the coastal waters of Iran and Turkmenia. The roach is a migratory fish which enters the Iranian border river Atrak and other Iranian rivers such as Gharesoo and Gorganrood for spawning. In Gorganrood river (southern Caspian Sea), reproduction migration starts in January-February, and continues until the spring (April). During this period, the fish travels 70-80 km up-stream [1]. Mostly the migration takes place at water temperature of 10-12 °C. Determination of sperm quality parameters such as seminal plasma composition, sperm production characteristics and sperm motility could provide us with knowledge to develop and improve artificial reproduction in fish farms [2]. Seminal plasma produced by the sperm duct provides an ionic environment that maintains the viability of spermatozoa after their release from the testes [3]. Various factors can affect sperm motility such as pH, temperature, ions and osmolality [2]. Some other parameters have been used to estimate the viability of sperm, such as mineral content of the seminal plasma, especially Na+ to K+ ratio [4]. The biochemical composition of teleost semen has been studied by many researchers over the years [5-7]. Some parameters such as spermatocrit, sperm density, fertilization capacity, pH, osmolality and seminal plasma composition are used in the evaluation of sperm quality [8]. The seminal plasma analysis includes inorganic constituents (Na+, K+, Ca+2, and Mg+2) involved in the process of inhibition or activation of sperm motility [9,10]. Correlations between seminal plasma composition and sperm motility have been documented in some species like Atlantic salmon, Salmo salar [4], common carp, Cyprinus carpio [11], bleak, Alburnus alburnus [12], rainbow trout, Onchorhynchus mykiss [13], Persian sturgeon, Acipenser persicus [14] and Chinook salmon, Onchorhynchus tshawytscha [15]. Studies suggest that Na+, K+ and Cl- ions are predominant in seminal plasma and that the concentration of these ions are lower in sturgeon than in teleost fishes. However, Na+ to K+ ratio is higher in sturgeon compared to teleost [16]. To have controlled and successful production in fish farming, it is important to have adequate knowledge of semen characteristics and composition of seminal plasma. In this study, we determined changes of ionic ratios and its effect on sperm characteristics during spawning migration in the Caspian roach.
Fish and sperm collection
Brood fish were caught from Gorganrood River three times during the spawning season, February 10, March 3, and April 5 (beginning, middle and last spawning season, respectively). Sperm of 10 mature males was collected by pressing gently on the abdomen at each sampling time. Sperm samples were collected with a syringe fitted with a plastic needle. We attempted to collect all the sperm at each stripping and avoided its contamination with urine, mucus or blood; to provide enough oxygenation to the sperm by maintaining enough headspace in syringe. A syringe of sperm from each male was placed on ice (4 °C) and immediately transported to the laboratory for analyses.
Experimental design
The fish were divided into 3 treatment groups in terms of ionic ratios obtained during sampling (February, March and April) for each period of sampling. Ionic ratios were divided to 3 treatments: Na+ to K+ (1: 5 - 6.4, 2: 6.5 –7.9 and 3: 8 - 9.5 mmol -1); Na+ to Ca+2(1: 15 –16.9, 2: 17 – 18.9 and 3: 19-21 mmol -1); Na+ to Mg+2 (1: 45 – 54.9, 2: 55 – 64.9 and 3: 65 - 75 mmol -1); K+ to Ca+2 (1: 5 - 6.4, 2: 6.5 –7.9 and 3: 8 - 9.5 mmol -1); K+ to Mg+2(1: 1–1.9, 2: 2 –2.9 and 3: 3 - 4 mmol -1); Ca+2 to Mg+2 (1: 1.5 - 2.4, 2.5-3.4 and 3.5- 4.5 mmol-1).
Sperm analysis
Sperm motility was triggered directly in activation medium 0.3% NaCl at ratio1: 1000 and immediately recorded with a 3 CCD video camera (Panasonic wv.cp240, Japan) mounted on a dark field microscope (Leica, USA). The duration of sperm motility was measured immediately after initiation of sperm activation until 100 % spermatozoa were immotile and expressed as sperm movement duration. The percentage of motile spermatozoa was defined as the percentage of progressively motile spermatozoa within each activated sample. Progressively motile spermatozoa were defined as those actively swimming in a forward motion. Only forward moving sperm was judged as motile and sperm cells that vibrated in place were not considered motile. Observations were made within two hours of semen collection. The spermatocrit was defined as the ratio of volume of white packed material to the total volume of semen ×100 [17]. Semen was drawn into glass Microhaematocrit capillary tubes (75mm length, 1-1–1-2mm internal diameter) until 60–80% of the tube volume was occupied by semen. One end of the tube was then sealed with clay and the tubes were centrifuged for 8 min at 3000 rpm (Sigma, 13 USA). Spermatozoa concentration was calculated with haemocytometer by placing a droplet of diluted semen with 0.3% NaCl solution on a Thomas haemocytometer slide (depth 0.1mm) with a cover slip and counted using light microscopy. After a few minutes (to allow sperm sedimentation), the number of spermatozoa was counted at100X magnification and expressed as spermatozoa ×109 per ml. All experiments were performed in triplicate at room temperature (20-22°C). Semen volume was measured in graduated tubes and expressed in ml.
Seminal plasma characteristics
Sperm samples were centrifuged at 3000 rpm for 8 min (Eppendorf AG, Hamburg, Germany) and then seminal plasma (supernatant) was collected. Plasma was centrifuged twice to avoid possible contamination with spermatozoa. The samples were frozen at -20 °C until analysis. Ca+2 and Mg+2 of the seminal plasma were measured spectrophotometrically (S2000-UV/VIS, England). The concentration of Na+ and K+ were determined with flame photometer (Jenway PFP, England; Standard kits from Parsazmoon, Tehran, Iran).
Statistical analysis
A one-way analysis of variance (ANOVA) was carried out to determine variation in ionic ratios (Na+ to K+, Na+ to Ca+2, Na+ to Mg+2, K+ to Ca+2, K+ to Mg+2and Ca+2 to Mg+2). Data from individual fish on all ionic ratios and its effect on sperm quality parameters during spawning migration were analyzed using ANOVA. Before analysis, data were tested for normality and homogeneity of variance. Means were tested for significant differences by one-way ANOVA followed by Duncan’s multiple range test at 0.05 significance level. Statistical analysis was performed with the SPSS software. All mean values represent mean ± standard error of mean (SE).
The ionic ratios of the seminal plasma are presented in Table 1. Similar to other cyprinids, Na+ and K+ are predominant ions in the seminal plasma of roach. However, their concentrations are higher in sturgeon than teleost [16,18]. Depending on ionic concentrations, most of these ions are involved in regulating sperm motility either by contributing to the intracellular ionic composition or by regulating osmolality [19,20]. In this study, significant differences (P<0.05) were observed in ionic ratios of Na+ to K+ and Na+ to Ca+2 during spawning season (Table 1).Sperm movement duration changed slightly (P< 0.05) due to Na+ to K+ ratio (Fig. 1). A similar tendency was detected in the spermatocrit and percentage of motile spermatozoa due to the Na+ to Ca+2 ratio. Spermatocrit and percentage of motile spermatozoa changed with proportion of Na+ to Ca+2 ratio fluctuation (Fig. 2 a,b). Ionic ratios of Na+ to K+ and Na+ to Ca+2 decreased from the beginning to the middle part of the spawning period and increased towards the end of the reproductive period (Table 1). Ionic contents differ among studies, which could be due to stripping time (early, middle or late spawning season), environmental conditions (which stimulate regulatory functions of the endocrine system involved in spermiation), frequency of stripping and hormonal stimulation of spermiation [3,7,21]. In addition, Na+ to K+ ratio has been used to estimate the viability of sperm in fish [4]. The roles of ions in sperm motility in fish have been reviewed several times [16,21,22]. In Atlantic salmon, Salmo salar, there is a positive correlation between Na+ and K+ of seminal plasma [4]. Some studies have found a relationship between the composition of the seminal plasma and sperm motility [12]. On the other hand, no significant changes were observed (P>0.05) in Na+ to Mg+2, K+ to Ca+2and K+ to Mg+2 ionic ratios of seminal plasma (Table 1). Season can significantly affect the composition of seminal plasma [3]. No correlation has been observed between seminal plasma ionic parameters and sperm motility in Persian sturgeon, Acipenser persicus and rainbow trout, Onchorhynchus mykiss respectively [18,23]. Significant changes were observed in the Ca+2 to Mg+2 ratio during spawning season (Table 1, P>0.05), and a higher Ca+2 to Mg+2 ratio was observed during the middle part of the spawning period compared to beginning or the end (Table 1). It has been shown that there is a significant variation in seminal plasma composition at different times of the spawning season in rainbow trout [24], Persian sturgeon [18] and common carp [12]. In the present study, we found that percentage of motile spermatozoa was changed (P<0.05) due to Ca+2 to Mg+2 ratio (Fig. 3). Research has documented the effect of ions on the initiation of motility and the duration of sperm in an effort to determine the biosensitivity of sperm. Thus, the longer period of sperm motility in sturgeon compared to teleost fish may be due to higher Na+ to K+ and Ca+2to K+ ratios in seminal plasma [14,16]. However, this may also be attributed to the ATP content in sperm, the number of mitochondria, intracellular ionic osmolality or pH. It has also been demonstrated that longer duration of motility in carps compared to salmonids is due to existence of higher Ca+2 to K+ ratio in seminal plasma [16]. A different relationship was found between motility and K+ in Esox masquinong, both in terms of the percentage of motile spermatozoa and total period of sperm motility [25].
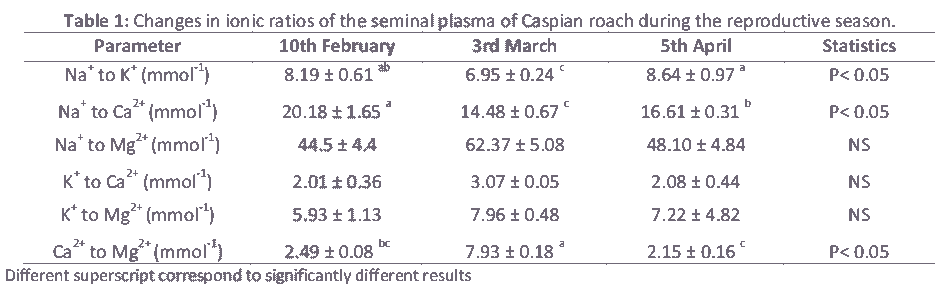
Table 1: Changes in ionic ratios of the seminal plasma of Caspian roach during the reproductive season.
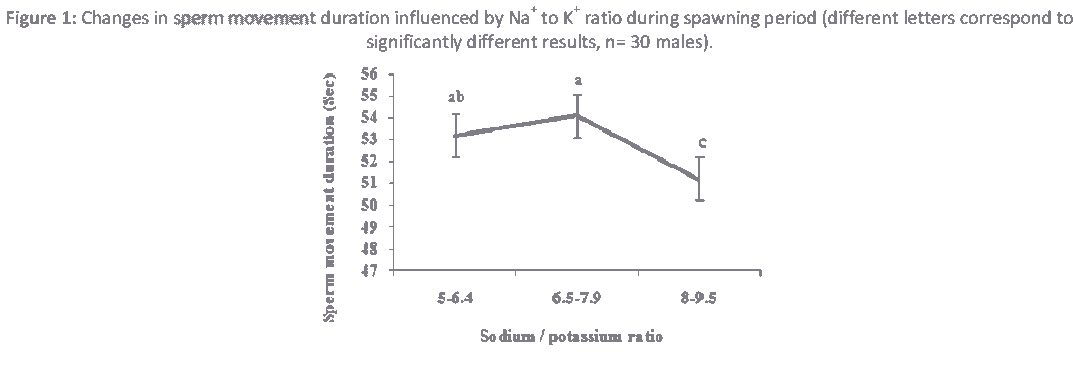
Figure 1: Changes in sperm movement duration influenced by Na+ to K+ ratio during spawning period (different letters correspond to significantly different results, n= 30 males).
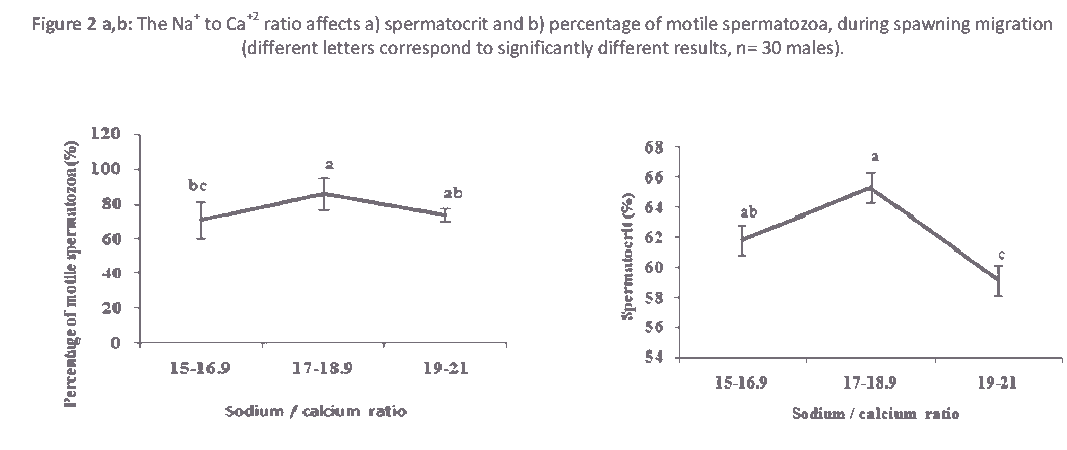
Figure 2: The Na+ to Ca+2 ratio affects a) spermatocrit and b) percentage of motile spermatozoa, during spawning migration (different letters correspond to significantly different results, n= 30 males).
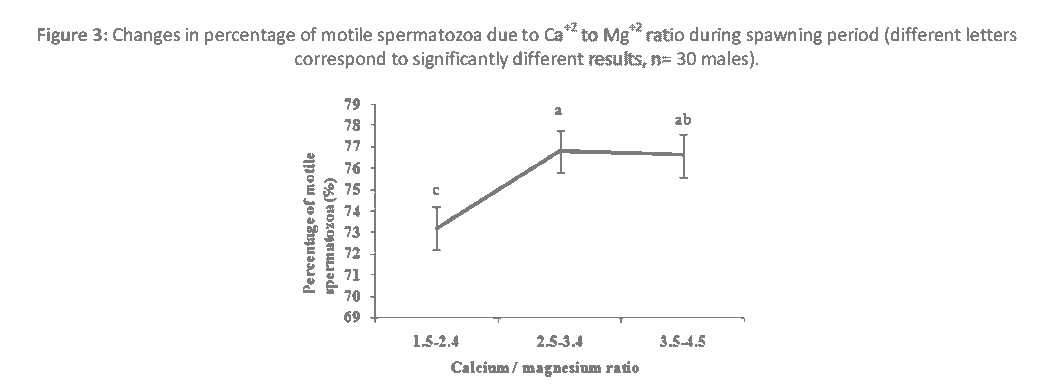
Figure 3: Changes in percentage of motile spermatozoa due to Ca+2 to Mg+2 ratio during spawning period (different letters correspond to significantly different results, n= 30 males).
The present study provides the first report about changes in ionic ratios of seminal plasma in Roach and its sperm characteristics response to ionic ratios of seminal plasma. Studies on seminal parameters are important to optimize the procedure of artificial fertilization in fish farming by improving sperm handling techniques such as fertilization technique (wet or dry) and storage method (short–term) of fish semen. The correlation between ionic ratios and sperm characteristics is an important aspect for future experimentation.
This work was financially supported by Gorgan University of Agricultural Science and Natural Resource, Islamic Republic of Iran. Otherwise, the authors declare that they do not have any competing interests.
All authors contributed more or less equally to this research work.
We are grateful to staff of the central lab in Gorgan University. The authors wish to thank M.Sc. students, Mahdi Taati and Milad Kabir, for their valuable contributions.