Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2014) Volume 2, Issue 4
Like plant leaves, suspension cultures of photoautotrophic Oxybasis rubra Fuentes-Bazan (syn. Chenopodium rubrum L.) cells pass through distinct developmental phases when grown under CO2 as the sole carbon source: an initial cell division phase of 4 weeks, a stationary phase of another 4 weeks and an aging phase (3-4 weeks) when the cell senesce and finally die. These phases are reflected by differential gene expression. A gene that was strongly expressed in the course of the stationary phase but much lesser during the exponential growth phase of the cell culture was isolated from a cDNA-library of stationary cells and completed by 5’-RACE. From homology analysis, the gene was tentatively identified as glycan exo-hydrolase (Oxybasis rubra glycan exo-hydrolase, OrGEH). Heterologous expression in E. coli yielded a protein with a preference to hydrolyze the ß-D-galactopyranoside, ß-D-fucopyranoside and ß-D-glucopyranoside of the corresponding artificial p-nitrophenyl substrates. Possible function of the protein in cell-wall metabolism was confirmed by extracellular localization of the GFP-fusion protein in transformed Nicotiana benthamiana leaves, by an N-terminal signal sequence for the secretory pathway and by the positive response of gene expression to auxin. Suspension cultured OrGEH RNAi-lines showed not only strongly reduced cell division activity but also lack of the stationary phase during which the cells usually double their biomass by extension growth. Although the natural substrate of the protein is not known we propose a function of OrGEH in the processing of cell wall components during cell division and cell extension growth.
Keywords: Cell culture; Cell wall metabolism; Oxybasis rubra Fuentes-Bazan (syn. Chenopodium rubrum L.); Development and gene expression; Glycan Exo-Hydrolase
OrGEH, GH3: Glycan Exo-Hydrolase From Oxybasisrubra 2,4-D, 2,4-Dichlorophenoxyacetic Acid; DD: Differential Display; GFP: Green Fluorescent Protein; GH3: Glycosyl Hydrolase Family 3 IAA, Indole-3-Acetic Acid; MDE: Mutation Detection Enhancement MOPS: 3-(N-Morpholino) Propanesulfonic acid; PCR: Polymerase Chain Reaction; pNP: Para-Nitrophenyl; PTGS: Post-Transcriptional Gene Silencing; RACE-PCR: Rapid Amplification of cDNA-ends with PCR
Suspension-cultured plant cells are powerful tools to dissect complex physiological processes at the cellular level which, in a plant tissue, may be partly superimposed and blurred by trans-cellular metabolic interactions. Another advantage of these systems is the direct accessibility of the cells through the medium. Most plant cell cultures are heterotrophic but from a few species, photoautotrophic cell cultures have been established which can serve as models of photosynthesizing cells of higher plants. One of the best characterized photoautotrophic cell cultures is that of Oxybasisrubra (formerly Chenopodium rubrum [1,2-13]). As long as favorable environmental conditions are maintained, these cells divide continuously without differentiation. Three growth phases can be described for a batch culture with CO2 as the only carbon source: a cell-division phase (for 3 to 4 weeks), a stationary phase of another 4 weeks and a phase of aging and senescence. The majority of the cells die between week 11 and 12. During the stationary phase, size and biomass of the individual cells increase significantly [2]. With regard to the life-span and the developmental phases, a batch culture of O. rubra resembles a leaf of a mesic plant. When examining gene expression in the course of a batch culture by mRNA differential display, we saw clear differences between the juvenile, dividing cells and those of the stationary phase (Appendix S1 A in Supporting Information). Focusing on stationary phase cells, an attempt was made to isolate differentially expressed genes and to investigate the functions of the corresponding proteins. One hypothesis was that some of the respective gene products might be involved in cell growth. Here, we report the isolation of the full-length cDNA of a glycan exo-hydrolase (Oxybasisrubra glycan exohydrolase, OrGEH), its comparison with similar genes of plant origin, expression in the course of the cell culture, localization of the gene product, heterologous expression in E. coli and the characterization of the enzyme activity. Although the natural substrate of the enzyme is not known, transformation of the cells with an OrGEH-RNAi-construct suggested the involvement of OrGEH in cell wall metabolism during both, cell division and growth. This conclusion was corroborated by the stimulation of gene expression when auxin was added to the medium of the cell culture.
Suspension culture of photoautotrophic Oxybasisrubra cells for suspension culture of photoautotrophic cells of O. rubra, the protocol of [1] was followed.
Extraction of RNA
O. rubra cells (1 g fresh weight) were washed three times with deionized sterile water. Liquid was removed by suction with an inlet filter and the cells were immediately frozen in liquid nitrogen and ground. Total RNA was extracted with Hybaid RecoveryTM-RiboLyser-Kit Green (Hybaid Ulm, Germany) as recommended in the instructions of the supplier. Total RNA was finally dissolved in 100 μl DEPC-treated water. mRNA was isolated with the Dynabeads® mRNA purification kit (Life Technologies, Darmstadt, Germany).
Amplification, SSCP and cloning of cDNA fragments, cDNA fragments selected from dd were re-amplified by PCR using the combination of 32P-labelled oligo dT-primer and the arbitrary primer A8 (5’-GTGACGTAGG-3’, Roth, Germany) at a final concentration of 2 μM and 15 U Taq polymerase (Boehringer Mannheim, Germany) per reaction. One μl aliquots of the PCR reaction volume were mixed with 10 μl loading MDE (Mutation Detection Enhancement)-dye, boiled for 2 min at 94 °C, rapidly cooled on ice and subjected to electrophoresis on a 0.5 x MDE [14] gel that was run at 60 W for 3 h. The gels were exposed on X-ray film for autoradiography. cDNA fragments producing strong signals were eluted and re-amplified by PCR with the same primer combination as above. Purified cDNA fragments were then cloned into the vector pT-Adv (Clontech, Heidelberg, Germany).
Construction and screening of a C. rubrum cDNA library
A cDNA library was constructed with mRNA isolated from 35-daysold, auxin-treated suspension-cultured C. rubrum cells. A ZAP-cDNA® Synthesis Kit (Stratagene, California) was employed for reverse transcription of the mRNA into cDNA. cDNAs were separated into 12 fractions representing different size classes and small cDNAs (< 400bp) were removed with SizeSepTM 400 columns (Pharmacia, Uppsala, Sweden). Fractions 6 to 8 and 9 to 12 respectively were combined and subjected to cloning and packaging according to the instructions. The two cDNA libraries were finally combined into a representative cDNA library of 2 x 106 clones. For screening of the library (according to the instructions by Stratagene), the radioactively labeled cDNA fragment no. 3 obtained from dd was used. Selected clones were subjected to in vivo excision and sequences were determined by Eurofins MWG Operon (Ebersberg, Germany).
RACE-PCR to complete the 5’ends of the cDNA
5’ends were produced with the SmartTM RACE cDNA Amplification kit (Clontech, Heidelberg, Germany) according to the instructions of the supplier.
Constructs for post-transcriptional gene silencing (PTGS)
PTGS (or RNAi)-constructs were made in two subsequent cloning steps using two different vectors. The pHannibal vector was used for the first step. Amplification of part of the coding region of the target gene in sense and antisense orientation was accomplished by PCR with the primer pair 5’-CTCTAGACTCGAG-3’ and 5’-CCATCGATGGTACC-3’. After restriction digest with XhoI and KpnI, cloning into pHannibal was carried out (1. Cloning step, sense part). Thereafter, the PCR product was digested with XbaI and ClaI and cloned into pHannibal (1. Cloning step, antisense part). The T-DNA was then cut from pHannibal and cloned into the vector pART27 (2. cloning step).
Construction of cDNAs encoding either the GFP- or a 6xHistarget- gene-fusion protein
In order to obtain a binary vector that allows expression of the target-GFP fusion protein, the coding region of the cDNA without stop-codon was amplified by PCR with the primer pair 5’-GGGGGA CAAGTTTGTACAAAAAAGCAGGCTTCATGTCATCTCGTAATA ACTTT-3’and 5’-GGGGACCACTTTGTACAAGAAAGCTGGGTCT TTCCCTATGTCAATGTGA-3’. The PCR product was cloned into the vector pDONR221 and thereafter introduced into the vector pK7FWG2 [15] with Gateway technology (Invitrogen). In order to obtain cDNA clones allowing the expression of the N-terminally His-tagged target enzyme, PCR was performed with the primer pair 5’-GGGGGACAA GTTTGTACAAAAAAGCAGGCTTCTCATCTCGTAATAACTTT TTT-3’ and5’-GGGGACCACTTTGTACAAGAAAGCTGGGTCCT ATTTCCCTATGTCAATGTGAAG-3’. The PCR-product was cloned into the vector pDONR221 and thereafter introduced into the vector pDEST17 with Gateway technology (Invitrogen).
Expression and purification of 6xHis-OrGEH-fusion protein
Plasmid DNA of OrGEH in pDEST17 was transferred into E. coli BL 21 Star Chemically Competent cells according to the instructions given by the supplier (Life Technologies). For expression of the 6xHis- OrGEH-fusion protein, transformed E. coli BL21* were incubated at 37 °C under shaking. When the bacterial suspension had reached an OD600 of 0.5, protein expression was induced with IPTG (1 mM final concentration). After induction, bacteria were further cultivated at 37 °C for 5 h and then sedimented by centrifugation (20 min at 5000 g, 4 °C). The pellets were washed two times with isotonic NaCl solution, re-suspended in 3 ml 1x MOPS-buffer (0.1M MOPS, 0.05M NaCl, 0.01M CaCl2, pH 6.0) and disrupted by sonification. After removing the insoluble material by centrifugation at 5000 g at 4 °C for 10 min, the supernatant was subjected to protein purification with a LKB FPLC system. The column contained Ni sepharose 6 Fast Flow-affinity medium. The fraction containing the purified protein was concentrated and taken up in 1 x MOPS buffer by ultrafiltration.
Determination of the protein concentration and substrate specificity of the bacterially expressed and purified OrGEH
Protein concentrations were determined with the Bradford assay [16]. An orienting substrate screening was carried out (see Appendix S2 in Supporting Information). Based on this test, the substrates number 4, 5, 6, 8, 9, 10 and 12 were selected for further analysis (Table 2). The assay mixtures contained 20 μl substrate solution (0.1 M), 50 μl MOPS buffer (1M MOPS, 0.5M NaCl, 0.1M CaCl2) (10x) and 20 μl enzyme solution (dilution 1:10 of the original solution obtained after purification) in a total volume of 500 μl (the pH was adjusted to 6.0 for all substrates except for pNP-ß-D-galactopyranoside where pH 5.5 was used) . The assays were incubated at 37°C until the color turned yellow and stopped with 1 ml NaHCO3 solution (1 M). The absorption (A) at 395 nm was read against blank containing water instead of enzyme solution. The activity was calculated with the following equation:
1 U [μmol x min-1] = OD395 x 0.091 μmol x incubation time [min]-1.
Determination of enzyme activity in O. rubra cells
Aliquots of 1 ml of O. rubra cell suspensions of different age were centrifuged at 14 000 rpm and 4°C for 10 min. The pellet was resuspended in 100 μl Na-acetate buffer (0.1 M, pH 5), ground with a blue pistil in a 1ml-Eppendorf microtube and centrifuged as before. Aliquots of 25 μl of the supernatant were used to measure ß-glucosidase activity (soluble fraction). For this, 10 μl of pNP-ß-D-glucopyranoside solution (100 μM) were added to the extract, the volume was adjusted with Naacetate buffer (0.1 M, pH 5) to 500 μl and incubated at 37 °C for 1 h. The reaction was stopped with 500 μl Na-carbonate solution (2%) and the absorption was measured at 405 nm. The pellets were re-suspended in 450 μl Na-acetate buffer (0.1 M, pH 5), and the enzyme activity was determined as described (insoluble fraction). Prior to reading of the absorbance, the suspension was centrifuged at 4 °C and 14 000 rpm for 10 min. One unit is defined as the amount of enzyme that cleaves 1 μmole of pNP-ß-glucopyranoside in 1 hour [17].
Transformation of O. rubra calli by particle bombardment
For particle bombardment, micro-calli of O. rubra were grown for 1 week on plates with MS-agar medium supplemented with 100 mM glucose. Gold particles (1 μm) were coated with 1 μg DNA (construct in pART27, prepared with HiSpeed™Plasmid Midi Kit, Qiagen, Hilde, Germany) according to the instructions by BioRad (Munic, Germany) and used for the transformation with the BioRad particle gun type PDS-100/He as recommended by the supplier. Transformed calli were selected on kanamycin-containing MS-agar medium (25 μg/ml kanamycin). Cell suspension cultures were established by transfer of an inoculum from the calli into liquid MS medium. 3 passages were required for a reasonably growing cell suspension culture.
Transformation of Nicotiana benthamiana leaves
Transformation of Nicotiana benthamiana (Col-0 ecotype) for transient expression was performed by infiltration of attached leaves with an Agrobacterium tumefaciens C58 pGV3121 suspension using a syringe. Two different types of Agrobacterium tumefaciens C58 pGV3121 suspension were used containing one of the following plasmids: i) coding region for green fluorescent protein (GFP) in vector pB7WG2, and ii) coding region for OrGEH in vector pK7FWG2 (for vectors see [15]). Two days after infiltration, green fluorescence was analyzed by confocal microscopy [18].
Confocal microscopy
Green fluorescence in leaves was analyzed with the confocal laser scanning microscope Leica TCS SP5 and documented with Leica software LAS. Collection of GFP signals was carried out after excitation with an argon laser (488 nm) in combination with a 510-525 nm emission filter. Chlorophyll fluorescence was simultaneously recorded (excitation as for GFP, emission filter 650-750 nm).
Isolation and identification of OrGEH as glycan-ßexohydrolase
Gene expression patterns of stationary phase cells (34-days-old) and exponential growth phase cells (12-day-old ) were compared by mRNA differential display (dd, see section of an autoradiogram in Appendix S1 in Supporting Information). Clear differences between the cDNA patterns of dividing and stationary-phase cells are visible. One of the bands representing transcripts that were pronounced in stationaryphase cells (number 3) was excised from the gel; the cDNA was eluted and re-amplified. Single strand conformation polymorphism (SSCP) revealed the presence of several cDNAs in that band (Appendix S1 part B in Supporting Information). The most prominent band was re-amplified and the cDNA fragment was cloned into the plasmid vector pCR-Script Amp SK(+). To obtain a full length clone, a cDNA library was produced from the RNA of 35-days-old, auxin-treated O rubra cells and screened with the cDNA-fragment no. 3. After in vivo excision, a 2451 bp clone was isolated and a full-length clone termed Alt6m42 was obtained by RACE-PCR and cloning. The nucleotide and derived amino acid sequences of the full length clone with a length of 2692 bp and an open reading frame between bp 59-2362 are deposited under the accession number AAp83934. The corresponding protein has a length of 767 amino acids and a predicted molecular weight of 83 kDa.
Table 1 shows plant amino acid sequences with homology to clone Alt6m42. The O. rubra protein exhibits a high degree of similarity to sequences from 19 species of Eu-dicots including 7 sequences from Arabidopsis thaliana, 7 sequences from monocots and one from a club moss. Based on the sequence relationship with the homologous sequences from A. thaliana and other plant species, Alt6m42 could be classified as putative glycan exo-hydrolase (therefore termed OrGEH for O. rubra glycan exo-hydrolase) belonging to the glycoside hydrolase family GH3 [19]. Multiple sequence alignment was performed for the conserved domain of GH3 enzymes. A high degree of conservation is obvious in the GH3-enzymes from Arabidopsis, rice, tomato, barley and O. rubra (Figure 1A). The active site of GH3 enzymes consists of two domains: the N-terminal catalytic domain A and the C-terminal non-catalytic but essential domain B [20], (Figure 1B). The most closely related proteins are A. thaliana putative ß-D-xylosidase 2 (BXL2, AF367266_1) and A. thaliana bifunctional ß-D-xylosidase/α-Larabinofuranosidase (BXL1, AED95802) (Figure 1C).
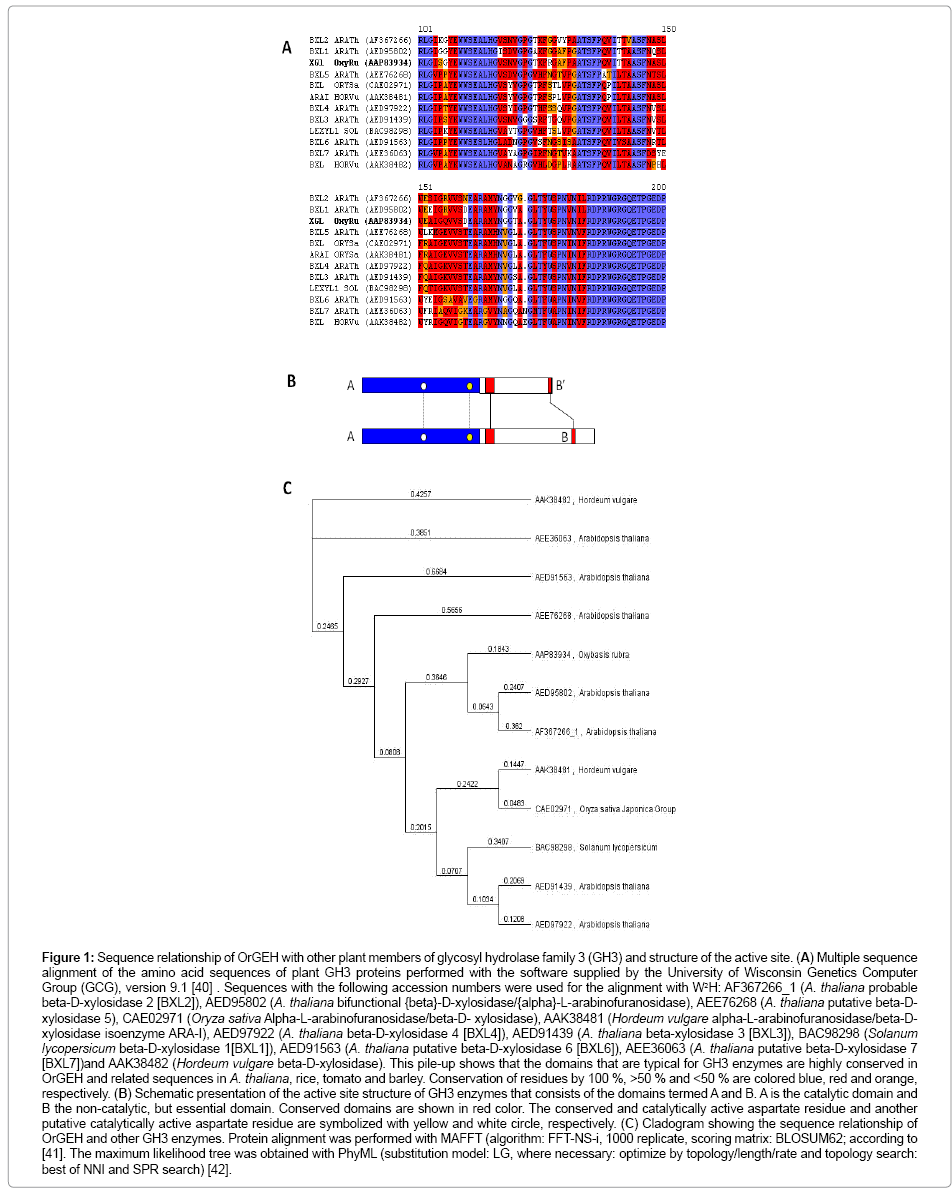
Figure 1: Representative photomicrograph of histopathological features in pulmonary necropsies.
| Organisms | Number of organisms | Organisms | Number of sequences per organism | Amino acid sequence similarity with Oxybasisrubra in % |
|---|---|---|---|---|
| Eudicots | 19 | Vitis vinifera | 9 | 86 - 66 |
| Ricinus communis | 5 | 85 - 68 | ||
| Populustrichocarpa | 6 | 85 - 66 | ||
| Prunus salicina | 1 | 85 | ||
| Cucumissativus | 8 | 84 - 68 | ||
| Glycine max | 8 | 84 - 67 | ||
| Citrus unshiu | 1 | 84 | ||
| Malus x domestica | 1 | 84 | ||
| Actinidia deliciosa var. deliciosa | 1 | 84 | ||
| Medicagotruncatula | 6 | 83 - 67 | ||
| Pyruspyrifolia | 1 | 83 | ||
| Fragaria x ananassa | 1 | 83 | ||
| Arabidopsis thaliana | 7 | 82 – 68 | ||
| Arabidopsis lyrata subsp. lyrata | 5 | 82 - 67 | ||
| Solanumlycopersicum | 3 | 81 - 70 | ||
| Camellia sinensis | 1 | 81 | ||
| Populus tremula x Populus alba | 2 | 76 - 67 | ||
| Medicago sativa subsp. x varia | 2 | 72 -71 | ||
| Raphanussativus | 1 | 71 | ||
| Monocots | 7 | Oryza sativa Indica | 2 | 76 - 71 |
| Brachypodium distachyon | 2 | 76 – 70 | ||
| Zea mays | 3 | 75 – 71 | ||
| Oryza sativa Japonica | 3 | 74 – 67 | ||
| Sorghum bicolor | 2 | 73 - 71 | ||
| Hordeum vulgare subsp. vulgare | 3 | 71 | ||
| Hordeumvulgare | 1 | 70 | ||
Club mosses |
1 | Selaginellamoellendorffii | 4 | 69 - 67 |
Table 1: Sequences from different groups of organisms with homology to the derived amino acid sequence of OrGEH from O. rubra.
Expression of OrGEH in the course of a batch culture of photoautotrophic O. rubra cells and in response to externally supplied auxin. For functional analysis of OrGEH in the course of a batch culture of photoautotrophic O. rubra cells, gene expression was examined between days 5 and 80. OrGEH transcripts were not detected until day 12 (Figure 2A and B) but two OrGEH-expression maxima were observed as the culture progressed (Figure 2A). The transcript level increased rapidly to reach a first peak at day 20 at the late cell division phase. Thereafter, the OrGEH-transcript level decreased rapidly but started a second increase at the beginning of the stationary phase reaching an even higher peak around day 50 in the late stationary phase. At the transition from the stationary to the aging phase, OrGEHtranscripts dropped rapidly and finally disappeared almost completely during aging and senescence (Figure 2A). The two peaks of expression of the OrGEH gene could suggest a role of the protein in cell extension growth. Phytohormones, in particular auxin, are known to be involved in this process. The photoautotrophic suspension culture of O. rubra showed pronounced fluctuations of the endogenous levels of auxin [1]. When the expression pattern of OrGEH in the batch culture (Figure 2A) was compared with the respective endogenous IAA contents [1], the increases in OrGEH transcript levels seemed to be preceded by peaks of IAA concentrations during the cell division and the stationary phase.
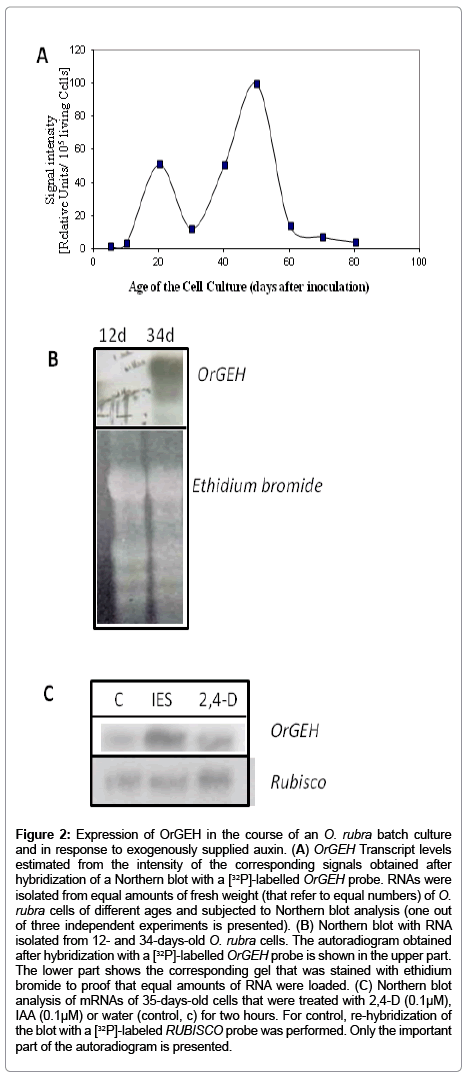
Figure 2: Representative photomicrograph of histopathological features in pulmonary necropsies.
To test how the expression of the OrGEH responds to the auxin level, IAA or 2,4-D was administered to a batch culture in the stationary phase for two hours. Controls were left untreated. Northern Blot analysis showed that both auxins, in particular IAA, triggered transcription of OrGEH (Figure 2C), but did not result in resumption of cell division.
Enzyme activity of heterologously expressed OrGEH and activity in O. rubra cells cDNA encoding OrGEH was cloned into the vector pDEST17 that allows the expression of the His-tagged protein. OrGEH was produced in E. coli BL 21* and purified via the N-terminal His-tag (see Appendix S2 in Supporting Information). Since the natural substrate is not known, substrate specificity was assayed with 12 different para-nitrophenyl-glycosides (see Appendix S3 in Supporting Information). The purified protein showed enzymatic activity with pNP-ß-D-derivatives of glucuronic acid, galactopyranose, fucopyranose, glucopyranose, xylopyranose and of cellobiose. The highest activities were obtained with pNP-ß-D-galactopyranoside, pNP-ß-D-fucopyranoside and pNP-ß-D-glucopyranoside, (Table 2).
| Substrate | Specific activity [mU mg-1] |
|---|---|
| pNP-β-D- glucuronide | 1.38 |
| pNP-β-D- galactopyranoside | 26.10 |
| pNP-β-D- fucopyranoside | 26.80 |
| pNP-β- Cellobioside | 4.70 |
| pNP-β-D- glucopyranoside | 17.26 |
| pNP-β- D-xyloside | 2.33 |
Table 2: Specific activity of purified heterologous OrGEH.
The ß-D-glucopyranoside-hydrolase activity was measured in the soluble and insoluble fraction of a homogenate of the O. rubra cells, using pNP-ß-D-glucopyranoside as substrate. Both fractions contained considerable enzyme activities, but that of the particulate fraction exceeded the soluble activity several-fold. The activity found in the particulate fraction of stationary-phase cells (50 days old, refers to the second expression maximum of OrGEH) was three times higher than that of young cells (25 days old, refers to the first OrGEH expression maximum) while no significant difference was found in the soluble fraction of cells of both age (Table 3).
| ß-glucosidic activity [mU cell-1 h-1] soluble fraction insoluble fraction |
||
|---|---|---|
| 25-days old cells 50-days old cells |
4.50 x 10-1 (0.66 x 10-1) 4.87 x 10-1 (0.58 x 10-1 ) |
10.52 x 10-1 (0.971 x 10-1) 30.37 x 10-1 (0.81 x 10-1) |
Table 3: Glucosidase activity in photoautotrophic O. rubra cells of the exponential growth phase (25-days-old) and stationary phase (50-days-old). O. rubra cells were disrupted and separated into a soluble and insoluble fraction, and the ß-glucosidase activity was determined in each fraction with pNP-ß-glucopyranoside. One unit is defined as the amount of enzyme that cleaves 1 μmole of pNP-ß-glucopyranoside in 1 hour (Keegstra and Albersheim, 1970).
Localization of the OrGEH gene product
Further sequence examination with TargetP (http://www.cbs. dtu.dk/services/TargetP), iPSORT (http://www.psort.org) and WoLFPSORT (http://wolfpsort.seq.cbrc.jp) indicated a signal peptide at the N-terminus and targeting to the secretory pathway and extraprotoplast localization which is in line with the enzyme activity found in the particulate fraction of the O. rubra cells. Thus, a function of the enzyme in cell wall metabolism seems likely.
Members of the GH3 family are known as hydrolases which cleave the ß-glycosidic bond releasing monosaccharides [21]. Two types of plant constituents could therefore be supposed as natural substrates: glycosylated secondary plant constituents or cell wall polysaccharides. The former compounds are commonly targeted to the vacuole or the plastids. Cellular localization of the enzyme could therefore be a clue to the function of the enzyme in planta. To that end, Nicotiana benthamiana leaves were transformed with a cDNA construct encoding OrGEH coupled to green fluorescent protein (GFP). For control, leaves were transformed with the same plasmid containing only the GFPgene. While GFP as a cytosolic protein was clearly located in that compartment (Figure 3A-C), the fusion-protein with OrGEH was only visible in the cell walls (Figure 3D-F) as expected from the enzyme activity of the cell wall fraction.
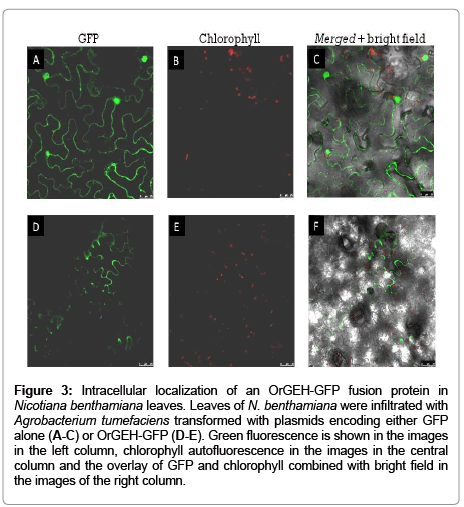
Figure 3: Representative photomicrograph of histopathological features in pulmonary necropsies.
Analysis of OrGEH by RNAi
RNAi-mediated knock-down of transcript levels was used to analyze the effect of reduced OrGEH transcript levels on cell division and growth. Stably transformed lines of photoautotrophic O. rubra calli were established by particle bombardment with either the pART27 vector containing the OrGEH RNAi construct or the empty pARTvector alone using kanamycin as a selection tool. The presence of the OrGEH sense plus antisense sequence in the OrGEH-RNAi-line was verified by PCR with genomic DNA (Figure 4A). Attenuated expression of OrGEH was demonstrated by semi-quantitative RT-PCR (Figure 4B).

Figure 4: Representative photomicrograph of histopathological features in pulmonary necropsies.
Three suspension cultures of the RNAi-line and the line with the empty vector each were started from transgenic calli using inocula of the same size. After two passages in liquid medium, growth of these three lines of both types of transformed cells was compared. By contrast to cells of the empty-vector-line, the growth of the OrGEH-RNAi-lines was slow and dramatically impaired as evidenced by the cell density and color (Figure 4D, E). Cells of the OrGEH-RNAi-lines showed a markedly reduced cell division rate and a shorter cell division phase (30 to 40 days) compared to the empty-vector-lines (50 days on the average) (Figure 4D).
Changes in gene expression during different growth phases of photoautotrophic O. rubra cultures In contrast to a continuous cell suspension culture, in a batch culture gene expression changes as the cells proceed from one developmental phase to the other [22]. Young, dividing O. rubra cells differ from older stationary-phase cells in their gene expression pattern (see Appendix S1 in Supporting Information. Similar changes have also been reported from A. thaliana suspension cultures. After termination of cell division, the phase of growth arrest was characterized by expression of genes that code for a member of the Fe (II)/ascorbate oxidase superfamily and for a protein with homology to ß-glucosidases [23]. Here, we describe and characterize a gene from Oxybasisrubra suspension-cultured cells that codes for a ß-glycan hydrolase which according to its DNA-sequence belongs to the multigene glycoside hydrolase family GH3 [19] (Table 1 and Figure 1).
Localization and function of the enzyme
Members of that family with a high degree of sequence similarity to the O. rubra enzyme are found in various mono- and dicotyledonous plant species (Table 1), e.g. in Arabidopsis thaliana (Figure 1A). Plant ß-glucosidases were described to be involved in cell wall metabolism, lignification, and the cleavage of O-glycosylated secondary plant constituents. The latter function plays an important role in defense, e.g. in the release of HCN from cyanogenic glycosides [24], or in the release of active forms of phytohormones from inactive conjugates [25,26]. In the sequence of OrGEH, the N-terminal signal peptide of 23 amino acids predicts targeting of the gene product to the secretory pathway (TargetP) or extracellular space (WoLFPSORT). Many proteins with corresponding N-terminal signal peptides that are targeted to the secretory pathway have been shown to be finally localized in the cell wall [27]. This concurs with the observation that the OrGEH-GFP fusion protein was detected in the cell walls of transformed Nicotiana benthamiana leaves (Figure 3) and the demonstration of the high ß-glycosidase activity in the particulate, i.e. cell wall fraction (Table 3).
The interpretation of OrGEH as cell wall enzyme fits also to the spectrum of potential substrates, although the natural substrates are still unknown. The O-ß-D-glycosyl residues that were preferred by the purified enzyme from transformed E. coli, namely (pNP)-ß- D-galactopyranoside (at pH 5.5), -ß-D-fucopyranoside and -ß-Dglucopyranoside (at pH 6.0) (Table 2) are known constituents of nonstructural plant cell wall polysaccharides subsumed under the term “hemicelluloses” [28]. Among the members of the GH3 family sharing a high sequence relationship with OrGEH (Figure 1A), several enzymes are involved in cell wall metabolism. The ß-D-xylosidase (LEXYL1; 70 % similarity to OrGEH) from tomato contributes to the modification of the cell wall during the ripening of the fruits [29]. Expression of the barley ß-D-xylosidase XYL gene (BXL HORVu) was demonstrated in most barley tissues [30]. For the Arabidopsis thaliana AtBXL1 protein (BXL5 ARATh ; 69 % similarity to OrGEH), targeting to the extracellular matrix and putative activity on cell wall xylans have been suggested [31].
Potential natural substrates of the OrGEH
p-Nitrophenyl-O-D-glycosides are convenient substrates for activity tests but not the natural substrates of the glycosidases. Nevertheless, they are useful for getting an idea on the substrate specificity with respect to the carbohydrates. Since the polysaccharide composition of the O. rubra cell wall is not known proposals for a potential natural substrate of the OrGEH remain hypothetic. The following findings were considered as clues for a first guess of potential substrate(s). A major part of the OrGEH is associated with the cell wall. As long as suspension cultured cells of O. rubra are dividing or still capable of cell division, they should not produce a secondary cell wall. Therefore, the substrates must be components of the primary cell wall. Members of the GH3 family are exo-polysaccharidases and therefore cannot dissect branched polysaccharide backbones. Rather, they modify side-chains or degrade the backbone from the non-reducing end until they are stopped by the side-chains. Since OrGEH does not show activity with galacturonides, acid polysaccharides like pectins may be tentatively excluded as substrates. Rather, the natural substrates of the OrGEH may belong to the hemicelluloses. Polysaccharides of this group consist of ß-1→4 linked glucan or xylan backbones which are decorated with longer or shorter side-chains of e.g. galactose and fucose residues [32]. In Arabidopsis, 11 members of the GH3 family have been recognized by bioinformatics analyses which are ß-xylosidases or ß-glucosidases, and one is bifunctional, namely an α-arabinofuranosidase/ß-xylosidase [33]. These exoglycanases are less specific with respect to the glycosidic linkages of the monomers and thus could participate in various stages of a plant cell’s life.
Function of OrGEH in the course of the life of suspensioncultured O. rubra cells
In suspension cultures of sycamore and bamboo, ß-glycosidase activities are involved in cell wall metabolism and cell division, respectively [17,34]. The dynamics of the OrGEH transcript during a batch culture of O. rubra cells suggests involvement in cell wall metabolism upon cell division and in particular during the stationary phase when cell extension takes place which is evidenced by the doubling of fresh weight of cells (Table 4 and [2]). In line with this interpretation is the observation that treatment of stationary phase cells with auxin (2,4-D or IAA) reinforced the expression of OrGEH (Figure 3B) while not inducing cell division. The levels of (endogenous) IAA and cytokinins might be decisive whether cell division or cell expansion takes place. For cells of the tobacco cell line cv Virginia Bright Italia-0, the concentrations of NAA necessary to stimulate cell extension are much lower than to stimulate cell division [35]. The same might hold for the induction of cell division or growth in O. rubra cells by the phytohormones. In the cell division phase, higher endogenous IAA and cytokinin concentrations were found than in the stationary phase [1]. Limitations in cell wall metabolism should, however, impact both cell division and expansion. Consistent with the anticipated function of OrGEH in cell wall metabolism is the observation that cell division was dramatically impaired in the OrGEH-RNAi-lines as well as cell growth, indicated by the absence of the stationary phase (Figure 4D and C).
| 20 days | 50 days | |
|---|---|---|
| Cell number per ml | 20 x 105 x ml-1 | 29 x 105 x ml-1 |
| Fresh weight per ml | 67 mg x ml-1 | 176 mg x 105 x ml-1 |
| Fresh weight per cell | 34 ng | 61 ng |
Table 4: Cell number, fresh weight and fresh weight per cell of photoautotrophic O. rubra cells in the cell division phase (day 20) and the stationary phase (day 50).
Since we only suppose the natural substrate of the OrGEH, its function in the metabolism of the cell wall is likewise hypothetical. Xyloglucans were shown to play an important role for cell expansion and cell division [36]. Consistent with the proposed function of OrGEH in xyloglucan metabolism is the fact that a strong effect of auxin on the xyloglucans of the cell walls, i.e. promotion of solubilization and degradation, was reported [37]. Auxin induced the expression of OrGEH (Figure 2C) and is known to stimulate H+-extrusion, thus conferring a favourable acidic pH for enzymatic activity. Given that a certain form of a branched xyloglucan is the target of OrGEH, cleavage of the side-chains might decrease the strength of the non-covalent linkages between the cellulose microfibrills and the hemicellulose matrix [38,39] and thus would facilitate extension growth of the cell wall.[40-42]
Experiments/analyses were performed by O.V.D. (Figures 1, 2 and 4A, D and E, Table 1), W.P. (Appendix S1 Supporting Information), C.R. (Figure 3, 4 B and C, Table 3, Appendix S2 Supporting Information) and W.S. plus V.Z. (Table 2 and Appendix S3 Supporting Information). C.R. and E.B. wrote the manuscript, E.B. was the leader of research and is senior author.
This work was supported by the German Research Foundation (DFG) with a grant to E.B. (Be 473/37-1). We are grateful to Dr. Alfons Weig (University of Bayreuth) for performing sequence analysis with MAFFT and PhyML. Technical assistance by C. Meinen is gratefully acknowledged.