Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2013) Volume 3, Issue 1
Keywords: Antheridiogen factor (AF), Vesicle like sac formation factor (VLS-F), Bacillus amyloliquefaciens, Appressorium, Venturia inaequalis, RPC, Tandem electrospray MS
Fungi depend on specific intercellular messaging for managing sexual interactions yet the chemical components required for recognition of sexual partners reveals no common communication strategy. All Zygomycota analysed seem to depend on trisporic acid derivatives as mediators of sexual recognition [1] whereas the Dikaryomycota depend on a communication system based on small peptides [2]. The non-mycotan group Oomycota makes use of steroid compounds, e.g. the well characterized compounds oogoniol and antheridiol in Achlya spp [2]. In the plant pathogen, Phytophthora, a lipid-based compatibility system provides an additional regulatory level [2].
Infection by Venturia inaequalis is initiated in spring by ascospores. The germ tubes arising from ascospores penetrate through the cuticle, via an appressorium [3] and develop into multilayered pseudoparenchymatous structures, termed stromata that are fertilized by a special body termed antheridium. The result of this fertilization is the development of a mature pseudothecium containing asci and ascospores (Figure 1), we have partially purified antheridiogen factor from culture supernatants of Bacillus amyloliquefaciens strain PFR-1. The factor inhibited appressorium formation and cause development of terminal antheridia of germinating tubes of Venturia inaequalis conidiospores. The mechanisms by which this factor initiates antheridium formation are unknown. There is a possibility that these factors would turn down the expression of the genes involved in appressorium formation and instead, turn on genes that lead to antheridium development. Also, this fungus, entersits their host by means of the appressorium [4,5]. The role of AF in controlling the infection needs to be investigated.
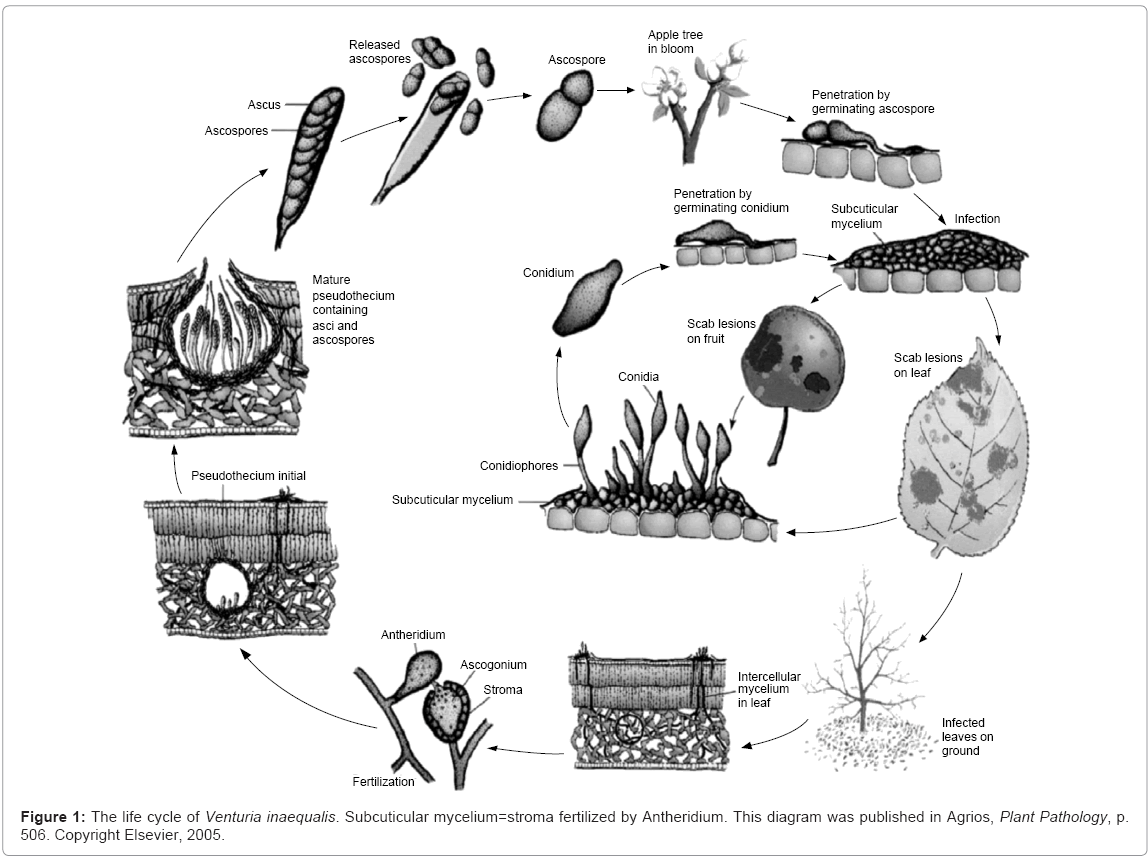
Figure 1: The life cycle of Venturia inaequalis. Subcuticular mycelium=stroma fertilized by Antheridium. This diagram was published in Agrios, Plant Pathology, p. 506. Copyright Elsevier, 2005.
Fungal strain
Venturia inaequalis strain MNH120 New Zealand was maintained on potato dextrose agar. Conidia were stored at -80°C in sterile MQ water containing 10% glycerol.
Bacterial strains Bacillus amyloliquefaciens PFR-1, Bacillus subtilis subsp. Subtilis (Ehrenberg) Cohn ATCC 605l, Bacillus subtilis subsp. spizizenii ATCC 6633, and Bacillus subtilis ESR 151, were maintained on Nutrient agar (Difco).
Strain identification
B. amyloliquefaciens Strain PFR 1 was identified according to the characteristics of morphology, physiology and biochemistry tests as described by Claus and Berkeley [6] and Gordon et al. [7] and by the comparison of the16S rDNA sequence.
Antheridium assay and growth conditions
Following inoculation of potato dextrose agar overlaid with asterile cellophane membrane production of Venturia inaequalis conidia was induced as described previously [8]. V. inaequalis conidia were harvested by transferring the growth on cellophane membrane into sterile tube contains sterile MQ water. The conidia were detached by inverting the tube gently several times. The spore suspension was filtered through cotton cheese cloth into a sterile tube. The spores were washed three times using sterile MQ water. The Conidial concentration was determined using a haemocytometer.
Isolation of apple wax
Surface wax from 200 Granny Smith apples was extracted by submerging the fruit for 30s in 2 liters of chloroform containing 0.5% (v/v) methanol. The wax solution was concentrated by rotary evaporation to approximately 0.13 g ml−1, and the preparation stored at -20°C.
Bioassay for appressorium and antheridium formation
Apple wax was smeared onto a glass microscope slide as described previously [8]. Suspensions of fresh, washed conidia were mixed with samples to be tested for activity. Droplets (10 μl containing approximately 100 conidia) were applied to a wax-coated microscope slide and the slides were incubated in a humid atmosphere at 24°C for 18-38 h.
Preparation of AF
Stock cultures of B. Amyloliquefaciens (0.5 ml) were inoculated onto each of 10 plates of Nutrient agar. The plates were left open in the Laminar flow hood for 1-2 h to dry, and incubated at 26°C for 36 h. For inoculation of the cultural medium, 0.5% glucose medium (2 ml) was added into each plate wascollected by scrubbing with a sterile cotton swab and transferred to a Scott bottle containing eitherto 1 litre of 0.5% glucose medium and or onto nitrogen base medium (Difco, Detroit, MI, USA) supplemented with 0.5% glucose in 1 l Scott bottle. The cultures were incubated at 26°C with shaking at 100 rpm. Cultures were sampled (1 ml) at different time intervals, centrifuged and the supernatant checked for the presence of AF and VLS-F. After 2-3 days of incubation, supernatants were collected by centrifugation at 26000×g for 15 min and the resulting supernatant was boiled for 20 min and recentrifuged at 40,000×g for 60 min to remove any precipitates. The final supernatants were aspirated and concentrated onto 20 ml under vacuum at 70°C, to 20 ml.
Fractionation of the culture supernatant Fluid (CSF)
Fractionation of the concentrated supernatant (Figure 4) was carried out by dialysis in 100-500 Da cut off dialysis membrane (Spectrum Laboratories Inc., CA, USA) against 600 ml of 20% ethanol at room temperature. The dialysate termed AF was concentrated to 10 ml under vacuum and kept on ice until use. The non dialysate was fractionated further by dialysis in 5 kDa cut-off membranes against 600 ml of 20% ethanol at room temperature. The dialysates from 5 kDa cut-off membranes were dried under vacuum. Then the solid residues were dissolved in 20 ml of methanol. The methanol soluble materials were dried under vacuum and dissolved in 10 ml MQ water. Fractionation of this portion was carried out by dialysis in 3 kDa MW cut-off membranes against 600 ml of 20% ethanol for three days at ambient temperature. The non dialysates causedformationof sac like vesicles of the germ tube termed vesicles like sac formation factor (VLS-F). Both AF and VLS-F were kept on ice bath until use.
Extraction of CSF and AF and VLS-F with organic solvents
Samples from CSF, AF and VLS-F were appropriately diluted with sterile MQ water to contain 1-2 units of factors (1 unit causes the development of antheridum of approximately 70% of germinating spores or the development of vesicles like sac formation of approximately 98% of germinating spores). Each portion was divided into four parts (10 ml). The pH of the first part (untreated) was approximately was 3.8; the second part was adjusted to pH 2 with hydrochloric acid, the third part to pH 6and the fourth part to pH 10 with concentrated ammonium hydroxide. Each sample was aliquotted into four fractions one of which was kept as acontrol and the remaining fractions were extracted with 10 ml of ethyl acetate, diethyl ether, or dichloromethane. The phases were separated by centrifugation. The solvent phases and the aqueousphases were separated and evaporated to dryness under vacuum. Each residue was re-dissolved in 10 ml of MQ water and bioassayed along with the unfractionated sample as control.
Stability studies
The temperature stability of the AF and VLS-F were examined by exposure of 1 ml of AF and VLS-F to 80°C, 100°C and 120°C for 10 min followed by cooling to room temperature and bioassay. AF and VLS-F (200 μl) were a treated (2 h, and 18 h at 37°C) with two proteases, Trypsin and Proteinase K (1 mg ml−1 in 20 mmol l−1 phosphate at a pH of 6). The enzymes were inactivated by exposure of the mixture to 100°C for 10 min followed by cooling and bioassay. Samples of heat inactivated enzymes were used as a negative control.
Purification of AF and VLS-F activity
A) Fractionation of AF and VLS-F by ion exchange chromatography: The pH of AF and VLS-Fwere adjusted to pH 5 and 8 using KOH respectively and fractionated on HiTrap SP FF or HiTrap DEAE FF respectively (GE Health Auckland, New Zealand) and were eluted with a linear gradient of 1M NaCl at appropriate pH.
B) Fractionation of AF by reversed-phase liquid chromatography (RPLC): Samples of AF in 0.1% trifluoroacetic acid (TFA)were subjected to RPLC by using a 1-ml HiTrap RPCcolumnon an AKTA Explorer high-performance liquid chromatography system (Amersham Pharmacia, Auckland, New Zealand). Separation was carried out according to the manufacturer’s operating specifications using solvent A(0.1% TFA) and B {(100% acetonitrile (ACN) in 0.01% TFA)} using a stepwise gradient from 0-95 % (v/v) ACN containing 0.1% (v/v) TFA. Fractionswere driedunder vacuum and dissolved in MQ water. AF activity appeared in the flow through thereforea mixed-mode anion-cation exchange and hydrophilicinteraction chromatography (ACE-HILIC) methods wereemployed as described previously [9]. Determination of the biological activity was carried out as described above.
C) Fractionation of VLS-F by RPLC: Fractionation of the VLS-Fusinga 1ml HiTrap RPC column and detection of the biological activitywas carried out as described above. The active fractions were pooled, and evaporatedunder vacuum. The solid material was dissolved in 1 ml 30% ACN containing 0.1% TFA and diluted with 2 ml of 0.1% TFA, and reapplied onto the column. Samples (1 ml) were injected onto the column at a flow rate of 0.1 ml/min and washed with the same solvent at 0.2 ml/min for a further 10 min.The solvent composition of the mobile phase was change as follows: The program consisted of 450 min with a constant mixing ratio (B=5%) from t=0 to 60 min, a linear gradient (B=5% to 40%) from t=60 to 140 min, a linear gradient (B=40 to 42 %) from t=60 to 200 min, a linear gradient (B=42to 44%) from t=250 to 300 min, a linear gradient (B=44 to 46%) from t=300 to 350 min a linear gradient (B=46 to 48%) from t=350to 410 min, a linear gradient (B=48 to 55%) from t=410 to 450 min,a constant ratio (B=95%) from t=450 to 480 min and a downward gradient (B=95%to 0%) from t=480 to 490 min. Portions of the respective HPLC fractions were saved for Tandem electrospray MSMS measurements.
B. amyloliquefaciens Strain PFR 1 was isolated from the root of a leguminous plant. Identification was carried out according to the characteristics of morphology, physiology, biochemistry tests and the comparison of a 16S rDNA sequence.
B. amyloliquefaciens Strain PFR 1 showed: 100% sequence identities to B.amyloliquefaciens strain FZB 42, 99% sequence identity to B.subtilis subsp. subtilis str. SMY SMY_ctg6 and 93% sequence identity to B.subtilis subsp. subtilis str. SMY SMY_ctg6.
Kinetics of appearance of AF
The CSF of B.amyloliquefaciens strain PFR1 from both a 0.5% glucose medium and nitrogen base medium was collected at various times and its absorbance measured (Figure 2). The activity appeared after 18 h of incubation. Maximal levels were reached after 7 days of incubation at 26 °C. The results indicated that the factorswere produced and released early in the growth cycle. CSF from three strains of B. subtilis, anduninoculated medium showed no AF or VLS-Factivity.
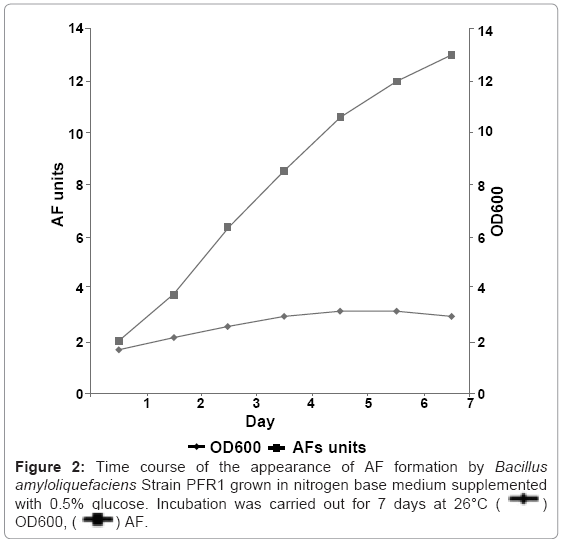
Figure 2: Time course of the appearance of AF formation by Bacillus amyloliquefaciens Strain PFR1 grown in nitrogen base medium supplemented with 0.5% glucose. Incubation was carried out for 7 days at 26°C ( ) OD600, (
) AF.
Preliminary characterization of AF
V. inaequalis conidia germinated on apple wax-coated glass slides produced appressoria (Figure 3a and 6a). Addition of boiled undiluted CSF (1.6 mg/ml protein)of B. amyloliquefaciens strain PFR1 to the conidia inhibited the germination,while dilution of 1/4of the CSF inhibited appressorium formation and resulted in formation of thick septated germinating tubes after 48 hr incubation at 24°C (Figure 3b). Adilution of 1/8 - 1/16 CSF caused the formation of sac like catenulate vesicles of germinated tubes of conidia (Figure 3c). Ethyl acetate, diethyl ether, and dichloromethane extracts of the CSF, at three different pHs, showed no activity. The aqueous phase from these extractions contained AF and VLS-F activities comparable to the original culture supernatant.

Figure 3: Inhibition of appressorium development and formation of vesicles like sac of V. inaequalis germinated conidiospores. Conidia were incubated without added CSF (A), or with diluted CSF (1/4) (B), diluted CSF (1/16) (C) on waxcoated glass slides for 18 h at 24ºC Bars represent 10 μm.
Partial purification of antheridiogen factors from CSF
Fractionation of the CSF was summarized in (Figure 4). AF (63 μg/ml protein) at a dilution of 1/64-1/128 caused the development of antheridia at the ends of germinated tube after overnight incubation at 24°C (Figure 5). VLS-F (4.8 mg/ml protein) at a dilution of 1/32 caused the development of vesicles like sac at the ends of germinated tubes after overnight incubation at 24°C (Figure 6c). Dilution of 1/64 caused the development of catenulate vesicles like sac after 48 h incubation at 24°C.
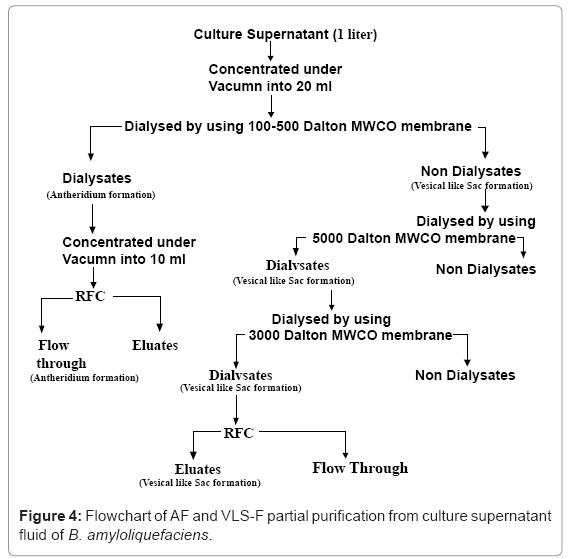
Figure 4: Flowchart of AF and VLS-F partial purification from culture supernatant fluid of B. amyloliquefaciens.
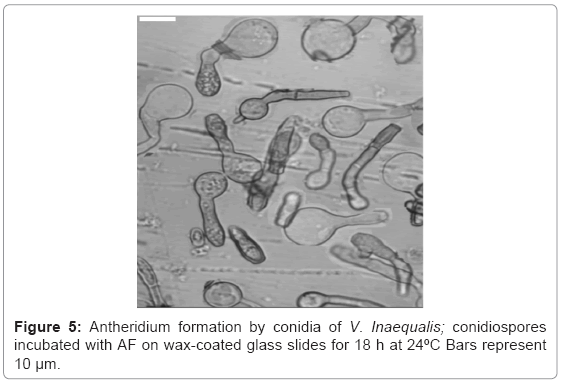
Figure 5: Antheridium formation by conidia of V. Inaequalis; conidiospores incubated with AF on wax-coated glass slides for 18 h at 24ºC Bars represent 10 μm.
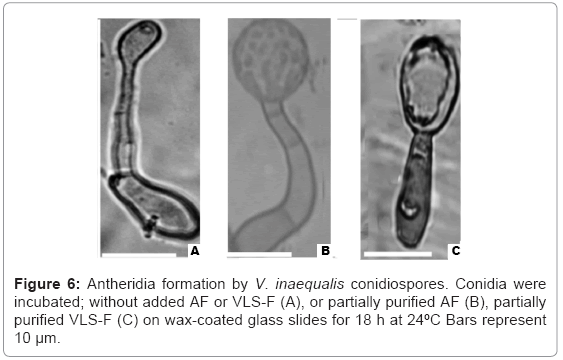
Figure 6: Antheridia formation by V. inaequalis conidiospores. Conidia were incubated; without added AF or VLS-F (A), or partially purified AF (B), partially purified VLS-F (C) on wax-coated glass slides for 18 h at 24ºC Bars represent 10 μm.
The first attempt to purify AF was carried out by employing SP and DEAE columns. The failure of the adsorption of the AF on cation and anion exchange resins is presumptive evidence that the AF molecules carry no positive and negative charges at appropriate pH.
The second attempt was to bind AF onto HiTrap RPC column (see materials and methods). The AF did not bind onto the column and appeared at the flow through (similar as in Figure 6b). The failure of the adsorption of the AF onto the HiTrap RPC column is presumptive evidence that the AF molecules are small and very hydrophilic molecule. Since the separation selectivity provided by RP-HPLC has been limited to the hydrophobicity based resolution of relatively nonpolar sample components (polar compounds present a significant challenge for RP-HPLC), a hydrophilic interaction chromatography (HILIC) was adapted. The AF did not bind onto the column and appeared in the flow through. Tandem electrospray MS MSMS showed a comprehensive list of hits from which the METLIN database was searched to come up with hits based on accurate mass data. The METLIN database search showed for any ion m/z mass, a number of hits, for example the ions at 198.0972 and 203.0526 are ammoniated and sodiated monosaccharide with the possible identities of one or mix of sugars. Also, the analysis showed the presence of phenolic compound derivatives. Thirteen short peptides (2-3 amino acid length) were present, but are likely to be anomalies. Synthesis of these peptides was carried out but none of the synthetic peptides had the activity AF properties. No charge envelope representing larger peptides and proteins were detected at all.
On other hand, VLS-F was bound into the column and eluted at approximately 44%-50% ACN, (similar result as in Figure 6c) (see materials and methods). The activity was detected in 8 fractions and each fraction with variable strength of activity. Since the profile of the elution from reverse-phase chromatography was complex, the fractions were pooled and further fractionation was carried out by gradual changing the solvent composition of the mobile phase (see materials and methods). The VLS-F activity was eluted at approximately 41-43% CAN (Figure 6c). These fractions inhibited the germination of the conidiospores (190 μg/ml protein). A dilution of 1/32-1/64 caused development of terminal vesicles like sac after 18 hr of incubation. Digestion with Trypsin or Proteinase K destroyed the inhibitory effect on germination of conidiospores but not the formation of vesicles like sac activity. Treatment with Trypsin or Proteinase K of the fractions resulted in the formation of catenulate vesicles like sac. Based on that, the most pure preparations of VLS-F have a complex elution profile from reverse-phase chromatography (Figure 7). Tandem electrospray MS revealed no results. Chymotrypsin digestion of the most active fraction followed by Tandem MSMS yielded several peptides but none of the synthetic peptides had the activity of vesicles like sac formation properties. Two peptides (amino acid sequences; MKKGFFLMAM and KLRGAYNKMRQL) tested showed an inhibitory effect on germination of conidiospores of V. inaequalis at 25 μg/ml and 50 μg/ml respectively.
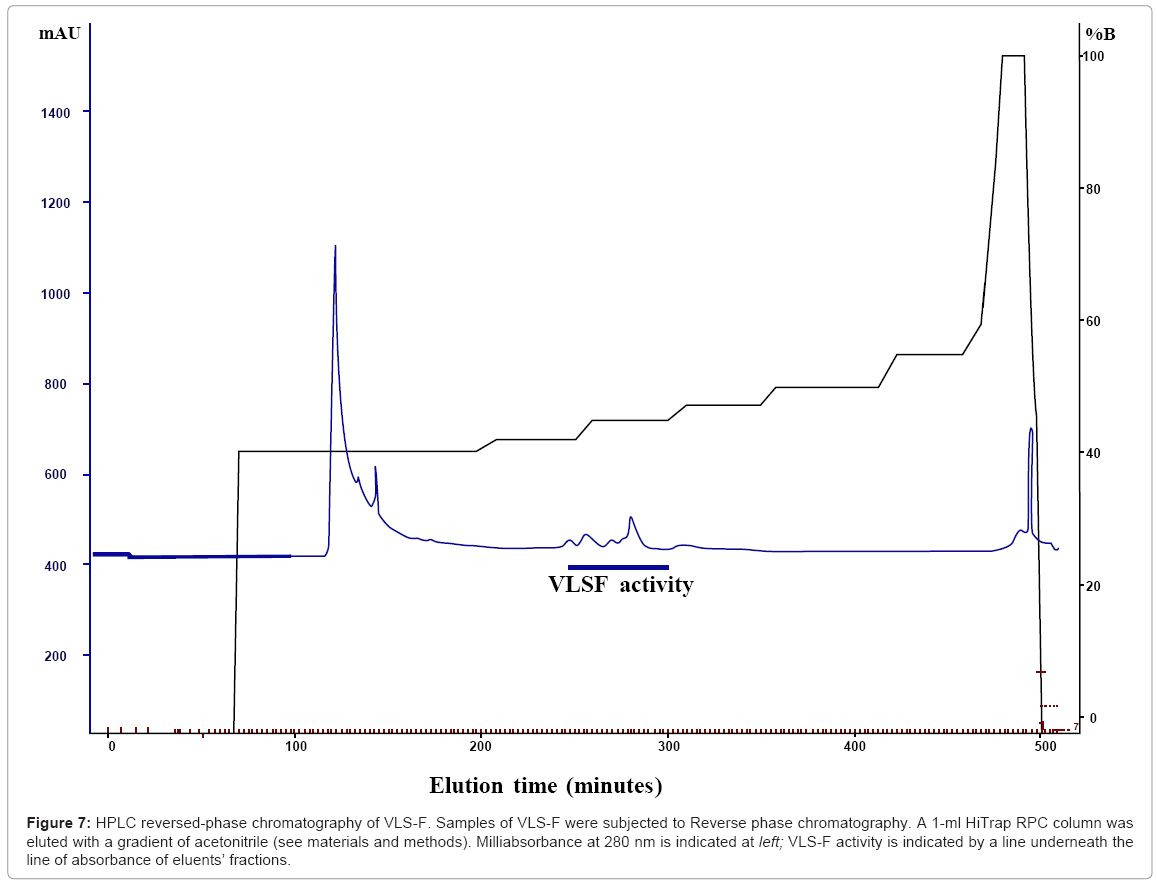
Figure 7: HPLC reversed-phase chromatography of VLS-F. Samples of VLS-F were subjected to Reverse phase chromatography. A 1-ml HiTrap RPC column was eluted with a gradient of acetonitrile (see materials and methods). Milliabsorbance at 280 nm is indicated at left; VLS-F activity is indicated by a line underneath the line of absorbance of eluents’ fractions.
Stability studies on AF and VLS-F
The sensitivity of AF and VLS-F to extremes of pH, proteases and elevated temperature was determined. Treatment at pH 2 or 10 for 2 h and 18 h failed to inactivate the AF and VLS-F (Table 1). A temperature of 80 and 100°C did not inactivate the activity of the both AF and VLS-F. Conditions routinely employed during autoclaving (121°C, 10 min) did not destroy the activity. Ethanol and methanol (final concentration 95%) also did not affect the activity. The proteases tested were effective in destroying the AF activity, suggesting that the AF activity could be due to one or more peptides containing the appropriate cleavage sit residues. On other hand proteases tested on VLS-F destroyed the inhibitory effect on germinating of conidia but not the formation of vesicles like sac activity, suggesting that the VLS-F activity was not due to a peptide.
| Treatment | AF (% Antherdium) | VLS-F (% Vesicles like sac formation) |
|---|---|---|
| Sterile medium | 0 | 0 |
| pH2·0 | 91±6·0 | 96 ± 3.0 |
| pH10·0 | 82·0±5·0 | 97 ± 1.0 |
| Trypsin | 4±0.5 | 88 ± 6.0 |
| Proteinase-K | 0 | 91 ± 4.0 |
| 121°C | 93 ± 4 | 94 ± 5.0 |
Table 1: Stability of antheridiogen and vesicles like sac formation factors to various treatments.
Estimation of the molecular weight of AF and VLS-F
To obtain an estimate of the molecular weight (MW) of the AF and VLS-F, a 10 ml sample of concentrated CSF was dialyzed in 100-500 MW cut-off membrane (Spectra/Por; Spectrum Medical Industries, Houston, TX, USA) against sterile MQ water (200 ml of 20% ethanol) for overnight at 4°C. Antheridia-stimulating activity (AF) was observed in dialysates suggesting that the MW is below 100-500 Da. To obtain an estimate of the molecular weight of the VLS-F, a 10 ml retentate from 100-500 Da MW cut-off membrane, was dialysed in 5 KDa MW cut-off membrane against sterile MQ water (200 ml of 20% ethanol) for three days at ambient temperature. The VLS-F stimulating activity was observed in dialysates. The dialysates were concentrated into 10 ml under vacuum and dialysed in 3 KDa MWCO membranes against sterile MQ water (200 ml of 20% ethanol) for three days at ambient temperature. VLS-F stimulating activity was observed in retentate suggesting that the MW is below is below 5 kDa and above 3 kDa.
This work was supported by a grant from the New Zealand Foundation for Research, Science and Technology C06X0812.