Fungal Genomics & Biology
Open Access
ISSN: 2165-8056
ISSN: 2165-8056
Research Article - (2011) Volume 1, Issue 1
Keywords: Shiga toxin – producing E. coli; Jejunal Haemorrhage Syndrome; Haemorrhagic Enteritis; Colonization; Prebiotic; Probiotic.
Mouldy feeds are less nutritious, impair health and decrease production in cattle [1]. JHS is a cattle disease that develops as a result of consuming mouldy feed with the disease complex consisting of Aspergillus fumigatus and Clostridium perfringens type A [2]. Recent studies have amended this complex to include only mycotoxins in feed components and STEC infections [3,4]. The latter studies did not identify Clostridium species in haemorrhaged tissues supporting earlier claims that this group of bacteria may overgrow rapidly after ruminant death masking the presence of causative agents. It is unclear whether other haemorrhagic diseases attributed to Clostridium infections are also associated with consuming mouldy feeds. For example, HE in adult goats is attributed to C. perfringens type C infections [5] with the clinical symptoms and pathology similar to JHS. Significantly, this disease occurs mainly in the winter months [6] indicating a potential link with the seasonal development of moulds in feeds and their associated deposition of mycotoxins [7]. A common report from producers experiencing JHS outbreaks in cattle is the ineffectiveness of antibiotic treatments [3,4] providing the impetus to develop novel approaches to managing mycotoxicoses. In monogastric livestock, probiotic and prebiotic feed additives mitigate pathogen issues in a number of ways including establishing beneficial bacteria in the intestinal tract [8,9], improving the ability of the mucosa to withstand infection [10,11,12], improving the defense system [8,9], binding toxins [3,4,13] and interfering with pathogen colonization sites [3,4,14]. Probiotics can improve milk production and rumen function in goats [15-21] with the greater benefits derived when there are environmental or pathogen challenges present. The aim of this study was to characterize the disease complex associated with a natural outbreak of HE in dairy goats during the winter months and to assess the effectiveness of probiotic and prebiotic applications to alleviate morbidity and production losses associated with the consumption of mouldy feeds.
The protocols were reviewed by the Agriculture and Agri-Food Canada Animal Care Committee and approved under ACC protocol 1131.
Animal and diets
Alpine, Nubian, La Mancha and Alpine-Nubian cross adults in a 280-head commercial dairy herd were used in the study. The goats were housed in an open stall barn with ventilation and had continuous access to water.
For 3 months, the herd received the production site diet consisting of corn silage, alfalfa silage and hay supplemented with a vitamin and mineral package. For two weeks, the herd received a reformulated diet with the alfalfa silage removed. Finally, the diet was replaced with a commercial complete feed ration containing barley, wheat mill run, corn, canola meal, dehydrated alfalfa meal, corn distillers dried grains, soybean meal, canola oil, limestone, salt, and a full profile of vitamin/trace mineral package (Table 1; New-Life Feeds, A Division of Parrish and Heimbecker Ltd., Lethbridge, Alberta) for the rest of the experimental period. This diet required supplementation with hay and water.
| Ingredient | Analysis |
|---|---|
| Crude Protein (min.) | 16.0% |
| Crude Fat (min) | 1.5% |
| Crude Fibre (max) | 12.0% |
| Calcium (actual) | 0.85% |
| Phosphorus (actual) | 0.45% |
| Sodium (actual) | 0.41% |
| Vitamin A (min.) | 20,000 IU/kg |
| Vitamin D3 (min.) | 2,000 IU/kg |
| Vitamin E (min.) | 35 IU/kg |
Table 1: Nutritional content of a complete commercial diet pellet supplied to the dairy goats. A 60 kg lactating goat will consume 3.5% of their body weight on a dry matter basis of total mixed ration (grain and forage) combined (2.1 kg), and received 0.9 kg of the 16% Lactating Goat Ration per head per day.
After 1 1/2 months with the commercial diet in place, half the herd received a prebiotic (Celmanax™ dry, 7 g/head/day ) or a probiotic (Dairyman’s Choice™ Dry, 7g/head/day plus 10 cc of Dairyman’s Choice™ paste on Day 0) for 28 days administered directly as a top dress in the individual trough. The Celmanax™ consists of a non-living formulation of yeast cell walls or mannan oligosaccharide (MOS) and yeast metabolites. The Dairyman’s Choice™ dry (Animal Pro- Products, Arthur, ON, Canada) consists of protected live yeast, yeast metabolites, vitamins and minerals. Dairyman’s Choice™ paste consists of two Bacillus strains, yeast metabolites, vitamins and minerals. All symptomatic goats received a 20 to 40 cc prebiotic: probiotic mixture [Celmanax™ liquid (20-40 cc) + Dairyman’s Choice™ paste (10 g)] to interfere with pathogen colonization [3,4], provide essential nutrients [22] and to absorb potential mycotoxins [3,4].
Experimental procedures
For the experimental period, the adult goats were monitored for the type and progression of clinical symptoms. All animals developing severe neurological symptoms were euthanized by the producer according to the codes of practice for the care and handling of goats in Canada (https://www.nfacc.ca/codes-of-practice/goats). Fourteen goats underwent a post mortem examination to assess the pathology and to characterize pathogens. Feed components were examined for mycotoxigenic fungi and mycotoxins. Two approaches were taken for determining the impact of the prebiotic and probiotic on the potential mycotoxicosis: 1) a spot treatment where a prebiotic: probiotic combination was orally administered to 28 symptomatic goats until the symptoms stopped; and 2) a prophylactic treatment where either a prebiotic or probiotic was applied to 140 goats for 28 days. For the prophylactic treatment, a randomized crossover design was chosen to minimize the influence of confounding covariates, such as parity, as each goat served as its own control.
Experimental observations and measurements
The goats were monitored for the “struck” symptom suggestive of the development of C. perfringens type C – based HE and other symptoms suggestive of a mycotoxicosis.
Prior to adding the commercial diet the herd was evaluated for body condition using standard criteria (https://www.smallstock.info/ tools/condscor/cs-goat.htm). A sub-sample of 40 goats per treatment group was evaluated for body condition after the goats consumed the commercial feed for 28 d and at the end of the prophylactic treatment.
Six dairy goats died from reproduction issues or Caseous Lymphadenitis during the same time period and served as negative controls. The tissue from 14 dairy goats confirmed the HE status with a focus in the jejunum based upon the presence of acute haemorrhaging in the jejunum, inflammation in the ileum and cecum, small blood clots in the small intestine and the lack of similar pathology in the control animals. A 30 to 60 cm piece of tissue was removed from the acute haemorrhaged region of the jejunum. The digesta and tissue were evaluated for bacterial pathogens. The method used has been described previously [3]. Released bacteria from tissue or digesta samples were stored at -80°C or serial dilutions were direct plated. Samples were applied to CHROMagar™ Salmonella, CHROMagar™ E. coli, CHROMagar™ O157, CHROMagar™ Salmonella Plus, and CHROMagar™ Listeria plates (Dalynn Biologicals, Calgary, Alberta). To confirm identity, the presumptive isolates were subjected to a GN-ID A + B biochemical test (Alere™ Canada, Ottawa, Ontario). Presumptive Salmonella were also subjected to a Salmonella Latex Agglutination test (Alere™ Canada, Ottawa, Ontario). Presumptive Listeria was identified using a Listeria ID kit (Alere™ Canada, Ottawa, Ontario). To detect C. perfringens, fresh tissue smears or digesta samples were examined for the presence of large, rectangular bacilli (rod) with or without spores (ovoid, sub-terminal) using a compound Nikon microscope set at 1000 x magnification. To distinguish potential STECs, serial dilutions of digesta or released bacteria from tissues were plated onto Sorbitol MacConkey agar (SMAC; Dalynn Biologicals, Calgary, Alberta, Canada) to identify non-sorbitol fermenting bacterial colonies and on Potato Dextrose agar (PDA; Dalynn Biologicals, Calgary, Alberta, Canada) to isolate fungi. Any suspect colonies were tested as O157 and H7 using the RIM™ E. coli O157 latex test (Fisher Scientific, Ottawa, Ontario, Canada). To further characterize the composition of the infections associated with HE cases, sorbitol-fermenting bacteria representing about 50% of the sample were re-examined in a DNA microarray (MaxiVir1.0) which carries 514 oligonucleotides of 70 bases in length targeting 348 virulence or virulence-related genes and 96 antimicrobial resistance or antimicrobial resistance-related genes found in gram-negative bacteria [3,4]. To select isolates, the original samples were transferred to SMAC, CT-SMAC and CHROMagar™ O157 and four STEC colonies were characterized based upon differences in morphology and Stx expression using a ImmunoCard STAT!® EHEC test (Somagen, Edmonton, Alberta, Canada) [3]. Because the adults had neurological symptoms suggestive of Listeriosis, a 200 g sample of alfalfa silage known to harbour such pathogens [23] was mixed with 50 ml of water and shaken for 1 hr at 200 rpm. A 1 ml aliquot was serially diluted, added to CHROMagar™ Listeria plates and incubated for 24 to 48 hr under standard conditions. This was repeated 4 times.
For the isolation of mycotoxigenic fungi, a 10 g sub-sample of corn silage, alfalfa silage or hay was finely ground and a 5 ml volume added to a PDA plate. The plate was incubated for 1 to 7 days and the individual fungal isolates transferred to new PDA plates. Fusarium isolates were identified by examination of micro-morphological characters [24] and by PCR amplification and sequencing of a fragment of the EF1-a gene and comparing the sequence with the FUSARIUM-ID database [25]. Penicillium isolates were identified by microscopic examination of morphology [26] and by PCR using partial beta-tubulin sequences, Bt2a and Bt2b [27]. Aspergillus species were identified by microscopic examination of morphology [28]. For extraction of mycotoxins, a 25 ml aliquot of 50% methanol was added to 3 g of ground sub-sample of corn silage, alfalfa silage or hay and placed on a shaker at 200 rpm for 3 h. The supernatant was collected in another tube, and stored at 4°C until use.
Milk production was monitored for two weeks after the animals had been consuming the commercial diet for 28 days. After the addition of the treatments, milk production was monitored for 28 days. Within each treatment group (140 goats), 40 goats were randomly selected and monitored for milk production for five days a week using a Waikato™ DHI milk meter. A milk sample was also collected for quality assessments once a week and these samples were analysed at a commercial laboratory (CanWest DHI, Edmonton, Alberta, Canada).
For comparing prebiotic and probiotic efficacy to antagonize STEC colonization, an in vitro cell culture (IVCC) adherence assay was performed as described previously [3]. The STEC isolates used in the assay were: E. coli O157:H7 E318N (A. Borezyk, Enteric Reference Laboratory, Ministry of Health, Toronto, Ontario) for comparison with earlier studies [3] and the non-O157 isolate from the current study. Briefly, the isolates were maintained at -20°C in 20% glycerol: 75% Nutrient broth (Sigma-Aldrich, Oakville, Ontario, Canada) and were grown statically overnight at 37°C in nutrient broth (Fisher Scientific, Ottawa, Ontario, Canada) as required. Each strain was serially diluted and bacterial cell counts were determined by plating on SMAC agar. Non-sorbitol fermenting colonies (O157 STEC) appeared as colourless colonies while sorbitol fermenting colonies (non-O157 STEC) appeared as pink colonies. The diluted bacterial suspension was added to a confluent monolayer of a bovine colonic cell line consisting of 105cells. The prebiotic was previously shown to significantly decrease E. coli O157:H7 E318N colonization while the Dairyman’s Choice paste had no effect on E. coli O157:H7 E318N colonization [3]. To determine the impact of the probiotic, Dairyman’s Choice™ dry, on E. coli O157:H7 E318N colonization, each well of confluent cells was washed once with PBS and then, 3 ml of DMEM was added with varying concentrations of the probiotic including 0%, 0.01%, 0.05%, 0.1% and 0.5%. The O157 STEC was added to the cell monolayer to deliver a final exposure dose of 106 CFU/105 cells. The cell cultures were incubated for 4 h under standard culture conditions. At the end of the experiment, cell-monolayers were washed with PBS and 2 ml of PBS supplemented with 0.25% Triton X-100 was added and incubated overnight at 4°C. Serial dilutions of the released bacteria were performed and quantified by plating onto SMAC plates. Plates were incubated at 37°C overnight and quantified by counting the non-sorbitol fermenting colonies that appeared as colourless colonies. To determine the impact of the prebiotic and probiotic on non-O157 STEC colonization, the threshold exposure dose was determined by adding 106, 105 and 104 CFU to the prepared cell monolayer. The cells were incubated for 4 hr and the wells processed as described earlier for the O157 STEC strain. An exposure dose that provided similar adherence to the O157 STEC strain was chosen and then used in the additional assays. Briefly, 3 ml of DMEM with 0% or 0.1% prebiotic or probiotic and 104 CFU of the non-O157 STEC were added to the prepared cell monolayers. At the end of the experiment, cell monolayers were washed with PBS to remove any unattached bacteria and 2 ml of PBS supplemented with 0.25% Triton X-100 was added. The plates were then incubated overnight at 4°C. Serial dilutions of the released bacteria were performed and quantified by plating onto SMAC plates. Plates were incubated at 37°C overnight and quantified by counting the sorbitol fermenting colonies that appeared as pink coloured colonies.
The lawn assay was previously used to examine the cytotoxicity of feed extracts containing mycotoxins in the absence and presence of 0.1% Celmanax™ and Dairyman’s Choice™ paste [3,4]. Enterocytes isolated from the jejunum served as the cell lawn and the extracts were applied to assess for cytotoxicity. Briefly, a 1% SeaKem Agarose (Mandel Scientific, Guelph, Ontario, Canada) support gel was poured into a petri dish. Next, the lawn agarose [3 ml of 3.7% SeaPlaque agarose (Mandel Scientific, Guelph, ON, Canada)] was mixed with 3 ml of enterocyte suspension and poured over the support agarose. Each extract (5 μl) was applied with or without 0.1 % prebiotic or probiotic and the plate incubated for 4 h under standard culture conditions. The lawn was stained with 0.1% trypan blue (Sigma-Aldrich) and destained using PBS. Plates were scored the same day and the amount of extract cytotoxicity was scored as low (1), moderate (2) or high (3) which was visualized as a faint blue spot, a blue spot or a dark blue spot respectively. These activities were compared to two standards, ground corn containing 0.1 ppm aflatoxin that had a cytotoxicity score of 1 or 1 ppm aflatoxin that had a cytotoxicity score of 3. The assay was repeated a minimum of three times.
Statistical analysis
Statistical analyses were conducted utilizing a repeated measures design (SYSTAT 10.2.01). The statistical model used for the analysis was
yijt = yo+μ+di+γj(i)+τt+(dτ)it+eijt,
where yijt is the measurement taken at time t on the jth goat assigned to the ith diet, yo is a covariate (pre-treatment measurement), μ is the overall mean effect, di is the ith fixed diet effect, γj(i) is the random effect of the jth goat within ith diet, τt is the fixed tth time effect when the measurement was taken, (dτ) it is the fixed interaction effect between diet and time, eijt is the random error associated with the jth goat assigned to the ith diet at time t. The body condition scores were analysed using a non-parametric Kruskal-Wallis one-way ANOVA. IVCC adherence assay data were log transformed to normalize the data prior to analysis. All other data were analysed using ANOVA followed by a posthoc Tukey’s test for comparison of the means. For the IVCC and lawn assay, the model consisted of two factors, passage number of the cell line or animal and dose. Results were considered significant if P < 0.05 and non-significant if P > 0.05.
The clinical symptoms in the dairy goats started with a “struck” appearance described for C. perfringens type C infections [5]. The symptoms progressed to grinding of teeth, cold extremities, rumen stasis, profuse drooling, hind-limb paralysis and no faecal pellets. The removal of the alfalfa silage from the diet did not curtail clinical symptoms or development of disease. In contrast, removal of the corn silage stopped all clinical symptoms and development of disease in 24 hr. This supported the diagnosis of a mycotoxicosis associated with the consumption of mouldy corn silage.
Celmanax™:Dairyman’s Choice™ spot treatment was associated with 50% recovery of symptomatic goats or 14 of 28 animals. Prior to adding the commercial diet, the average body condition score for the adult goats was a 2 or “skinny”. After the commercial diet was consumed for 31 days, the body score remained the same. The average body condition score after adding the prebiotic or probiotic significantly increased relative to the control period (Figure 1, P=0.001) with the biggest body condition score change associated with increased fat cover.

Figure 1: The impact of Celmanax™ (CEL) and Dairyman’s Choice™ (DC) on body condition scores for dairy goats (n=40, ***, P= 0.001).
The control animals had no pathology in the intestinal tract. All 14 symptomatic goats that succumbed to disease had the following common pathology: raised Peyer’s Patches in the jejunum, severe jejunal haemorrhages visible through the serosa, bloody digesta visible through the serosa, complete loss of mucosal structure in the haemorrhaged regions, dark-red erythema, small blood clots and oedema. Thus, the disease could be defined as HE with a focus in the jejunum, but the gross pathology was also the same as described for JHS in dairy and beef cattle [3,4].
Non-sorbitol fermenting colonies were identified at significant levels colonizing the haemorrhaged regions of the jejunum relative to the control animals (P = 0.001, Table 2). The bacterial isolates were confirmed as O157 and H7 using the RIM™ E. coli O157 Latex test, however there were about 50% of the colonies that were sorbitol fermenting suggestive of the presence of other pathogens. Presumptive O157 STEC appeared as mauve colonies with a small to large white halo on CHROMagar™ O157 plates. Presumptive O157 STECs were confirmed as pathogenic E. coli in the GN-ID A + B test. Other presumptive pathogenic E. coli appeared as blue colonies on CHROMagar™ E. coli plates. All presumptive pathogenic E. coli were confirmed as non-pathogenic E. coli in the GN-ID A + B test. White colonies were also found on this medium and were mauve with a white halo on CHROMagar™ E. coli O157. This provided a crosscheck for the results from the CHROMagar™ E. coli O157. Presumptive Salmonella were identified and appeared as mauve colonies on CHROMagar™ Salmonella plus plates, but were subsequently determined as false positives (pathogenic E. coli using the GN-ID A+ B test and the Salmonella latex agglutination test). Presumptive Listeria species were identified and appeared as pinpoint blue colonies with a halo on CHROMagar™ Listeria plates and these isolates were later determined to be Listeria grayi, a non-pathogenic strain. No C. perfringens was detected in tissue smears or digesta. Thus, the only pathogen identified in the haemorrhaged tissues or bloody digesta for goats with HE, were STECs. Listeria strains were also not identified in the alfalfa silage. There were four potential types of non-O157 STEC isolated from the tissue based on distinct morphologies on the CHROMagar™ O157 plates: mauve with a small to large white halo that had a circular form, raised elevation and undulate margin; mauve with no distinct halo that had an irregular to regular form, raised elevation and undulate margin. All bacterial colonies were sorbitol-fermenting and tellurite-resistant. The DNA microarray detected one genotype having the locus of enterocyte effacement (LEE) pathogenicity island, stx1A, stx1B, ehxA, hlyE, fimA, fimH, efa1, NleA (EHEC), NleA(EPEC), NleB (O157), IpfA, OmpA and OmpT. The isolate was tested for Stx expression and it expressed both Stx 1 and Stx 2. Interestingly, the absence of the stx2 gene in the DNA microarray analysis suggests that the detected Stx 2 from the original isolations may represent a strain that was ultimately not isolated or potentially the loss of the stx2 gene during sub-culturing [29].
| Sample type | Goats with HE (n=14) Mean Log CFU + SE | Goats without HE (n=6) Mean Log CFU + SE |
|---|---|---|
| Jejunum (per 2.5 cm2 mucosa)1 | 5.14 + 0.10a | 0.000 + 0.000b |
| Digesta (per ml) | 4.87 + 0.15a | 0.000 + 0.000b |
1 row numbers followed by different letters are significantly different P = 0.001.
Table 2: The average CFU O157 Shiga toxin – producing E. coli isolated from tissues of dairy goats with or without HE.
There was no Aspergillus fumigatus present in the samples (Table 3). There were several types of mycotoxigenic fungi present in the corn silage including F. culmorum, F. verticillioides, F. sporotrichioides, A. flavus, P. roqueforti, P. crustosum, P. aurantiogriseum, P. cyclopium and P. paneum. There were no mycotoxigenic fungi isolated from the alfalfa silage or hay samples. We also identified several fungi in the digesta and haemorrhaged regions of the HE cases including P. roqueforti, Trichothecium roseum, A. flavus and P. crustosum. There were no systemic fungal infections. The extracts from the corn silage produced cytotoxicity equivalent to a 1 ppm aflatoxin (in corn) standard or cytotoxicity score 3 (Table 4). Equivalent extracts from the alfalfa silage or hay had no effect on cell viability suggesting that the corn silage was the source of the mycotoxins. A 0.1% concentration of Celmanax™ eliminated the corn silage extract cytotoxicity in vitro (Table 4, P = 0.001). A 0.1% concentration of Dairyman’s Choice™ dry had no effect on silage extract cytotoxicity (Table 4, P > 0.05).
| Mycotoxigenic Fungi | Corn Silage | Alfalfa Silage | Hay |
|---|---|---|---|
| Fusarium culmorum | 100 | 0 | 0 |
| F. poae | 100 | 0 | 0 |
| F. verticillioides | 100 | 0 | 0 |
| F. sporotrichioides | 100 | 0 | 0 |
| Aspergillus flavus | 100 | 0 | 0 |
| Penicillium roqueforti | 100 | 0 | 0 |
| P. crustosum | 100 | 0 | 0 |
| P. paneum | 100 | 0 | 0 |
| P. flavigenum | 100 | 0 | 0 |
| P. aurantiogriseum | 100 | 0 | 0 |
Table 3: Percent of samples positive for mycotoxigenic fungi for corn silage, alfalfa silage and hay samples (n=4).
| Treatment | Cytotoxicity Index 1,2 (Mean + S.E.) |
|---|---|
| Control (50% methanol) | 0 + 0 a |
| CS | 3 + 0 b |
| AS | 0 + 0 a |
| H | 0 + 0 a |
| C + CS | 0 + 0 a |
| DC + CS | 3 + 0 b |
1 0 = no cytotoxicity or no blue spot; 1 = about 25% cell death or faint blue spot; 2
= about 50% cell death or blue spot; 3 = about 100% cell death or dark blue spot.
2 column means followed by different letters are significantly different from each
other, P < 0.001.
Table 4: Impact of 0.1% Celmanax™ (C) and Dairyman’s Choice™(DC) dry on the cytotoxicity of corn silage (CS), alfalfa silage (AS) and hay (H) extracts, n=3.
The feed additives significantly increased milk production with changes detected within the first 24 hr (P=0.01; Figure 2). At the end of the study period, the amount of milk produced in the morning and evening equilibrated further supporting the conclusion that there was a recovery in rumen function. The inclusion of the prebiotic or probiotic also significantly reduced somatic cell counts, but was particularly true for animals with somatic cell counts greater than 400 000 which represented sub-clinical or clinical mastitis cases (P=0.001, Figure 3). There was no change in the average percentage milk fat or protein, but there was a trend for these components to slightly decline during the study period, perhaps related to the increased volume in milk production (Table 5).
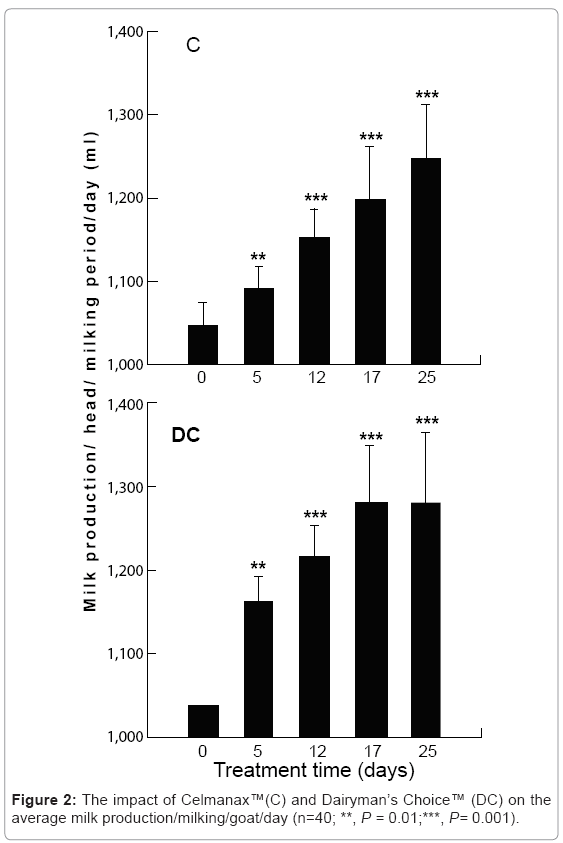
Figure 2: The impact of Celmanax™(C) and Dairyman’s Choice™ (DC) on the average milk production/milking/goat/day (n=40; **, P = 0.01;***, P= 0.001).
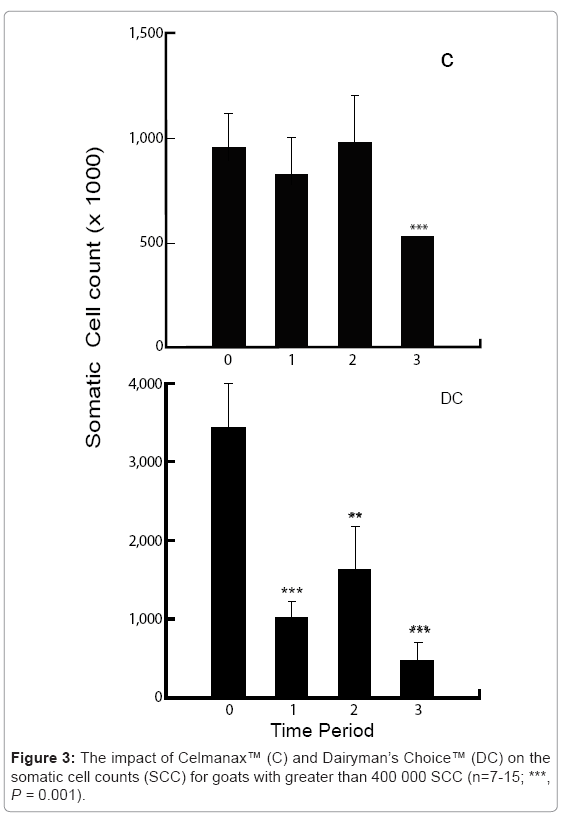
Figure 3: The impact of Celmanax™ (C) and Dairyman’s Choice™ (DC) on the somatic cell counts (SCC) for goats with greater than 400 000 SCC (n=7-15; ***, P = 0.001).
| Time (week) | Treatment | Average % Fat ±S.E.M | Average % Protein ± S.E.M. |
|---|---|---|---|
| 0 | Dairyman’s ChoiceÔ | 3.64 ± 0.08a | 3.27 ± 0.04a |
| 1 | Dairyman’s ChoiceÔ | 3.54 ±0.07a | 3.26 ±0.03a |
| 2 | Dairyman’s ChoiceÔ | 3.41 ±0.11a | 3.29 ±0.03a |
| 3 | Dairyman’s ChoiceÔ | 3.35 ±0.07a | 3.26 ±0.03a |
| 0 | CelmanaxÔ | 3.57 ± 0.08a | 3.35 ± 0.04a |
| 1 | CelmanaxÔ | 3.55 ±0.07a | 3.38 ±0.04a |
| 2 | CelmanaxÔ | 3.36 ±0.05a | 3.31 ±0.05a |
| 3 | CelmanaxÔ | 3.41 + 0.08a | 3.31 + 0.05a |
2column means followed by the same letter are not significantly different, P > 0.05.
Table 5: The impact of Celmanax™ dry and Dairyman’s Choice™ dry on the percentage fat and protein present in milk samples (n=40).
There was a significant dose-dependent reduction in E. coli O157:H7 E318N colonization of the bovine colonic cell line in response to varying concentrations of Dairyman’s Choice™ dry (P = 0.001, Figure 4). There was a significant passage effect (P=0.001) with no passage/treatment interaction suggesting that Dairyman’s Choice™ dry affected pathogen colonization regardless of passage number. The threshold dose for the reduction in E. coli O157:H7 colonization of the colonic cell line was 0.1% Dairyman’s Choice™ when compared with the control (P = 0.05, Figure 5). This resulted in about a 3-fold reduction in colonization per 105cells. Thereafter, there was a decline in E. coli O157:H7 colonization with a maximum effect at 0.5% Dairyman’s Choice™ when compared to the control (P = 0.001, Figure 4). This resulted in about an 8-fold reduction in colonization per 105 cells. We had previously reported similar reductions in E. coli O157:H7 E318N colonization by the prebiotic [3]. The non-O157 STEC strain was more efficient at colonization than the O157 STEC as evidenced by a 100 x lower exposure dose to provide the same adhesion as the E. coli O157:H7 E318N strain. Inclusion of 0.1% Celmanax™ liquid or Dairyman’s Choice™ dry significantly reduced the non-O157 STEC colonization relative to controls (P=0.002; Figure 5).
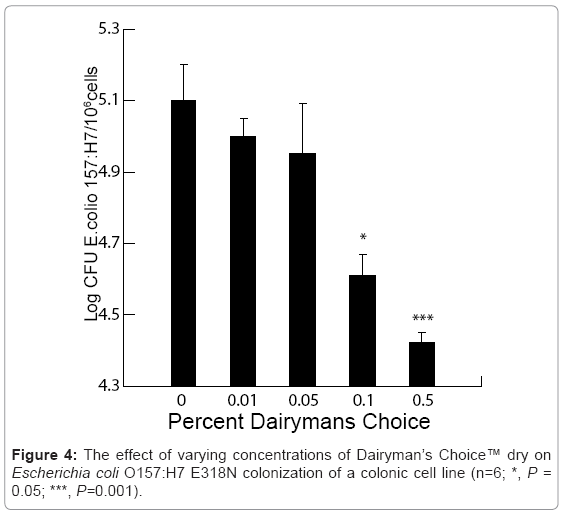
Figure 4: The effect of varying concentrations of Dairyman’s Choice™ dry on Escherichia coli O157:H7 E318N colonization of a colonic cell line (n=6; *, P = 0.05; ***, P=0.001).
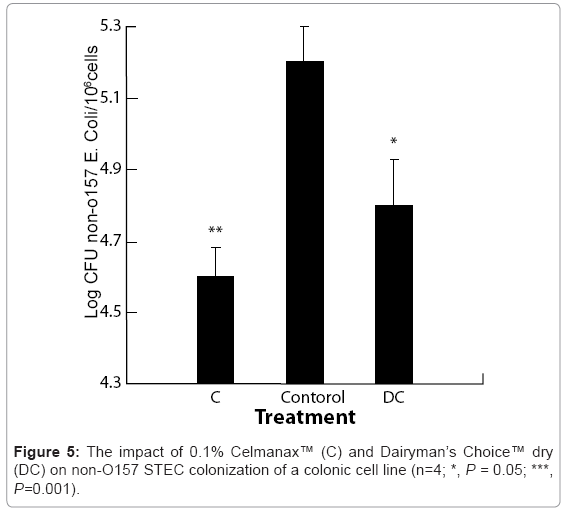
Figure 5: The impact of 0.1% Celmanax™ (C) and Dairyman’s Choice™ dry (DC) on non-O157 STEC colonization of a colonic cell line (n=4; *, P = 0.05; ***, P=0.001).
Mature ruminants are considered a reservoir for O157 and non-O157 STECs, with the amount and duration of shedding related to exposure dose [30,31], age [32] and mucosal factors [33]. Despite this information, it is unclear why one animal becomes infected while another does not. Here we describe an interaction between mycotoxins and STECs that resulted in colonization and the development of a specific disease in goats, HE. We recorded this same interaction in cattle with JHS supporting STEC as a pathogen affecting ruminant health with the driving force for colonization and carriage, mycotoxin exposure. This has substantial implications for improving food safety by simply applying mycotoxin binders to feeds to interfere with mycotoxin: STEC interactions.
HE is a common disease for adult goats during the winter months [5]. In the current study, the “struck” or stary eyed appearance of the goats during the early stages of the mycotoxicosis together with the similar gross pathology to C. perfringens type C associated HE suggests that this type of HE has gone undetected in adult goats. There is one case report documenting a chronic STEC infection in a weaning kid [34] with the adherent pathogen identified as E. coli O103:H2, a non-O157 STEC that produced Stx. In the current study, we describe O157 and non-O157 STEC infections in adult goats with the non-O157 STEC requiring a 100 times lower exposure dose when compared with the O157 STEC to achieve the same colonization [33,35]. This is a remarkable virulence factor that may have contributed to the rapid onset of symptoms observed in the goats. A possible explanation for the recorded differences in colonization may be related to structural differences in the FimH [36]. In support of this role, the application of the prebiotic: probiotic mixture stopped symptoms in 50% of the symptomatic goats. The prebiotic, Celmanax™, contains significant mannose that is known to block FimH-mediated non-O157 STEC adherence to target cells [37] and in vitro studies indicated that it caused a higher magnitude of reduction in colonization for the non-O157 STEC compared with the O157 STEC [3]. The probiotic also produced a decrease in O157 and non-O157 STEC colonization, but the mechanism is not likely related to the presence of mannose. Another virulence factor that differed between the STECs was the NleA (EPEC) gene which was found only in the non-O157 STEC. This is a non-LEEencoded effector protein that is not required for A/E lesion formation but contributes to bacterial colonization and increases the leakiness of the mucosa [38,39]. Here we observed a progression in the neurological symptoms of the goats that corresponded with high colonization rates by the STEC in the jejunum suggestive of mucosal leakiness as the mechanism for mycotoxin and Stx entry. Interestingly, the progression and similarity of neurological symptoms with mycotoxin-sensitive animals also support goats as a sensitive species [40].
There is conflicting evidence within the literature as to whether probiotics should be of host origin or not for livestock applications. For example, a host Lactobacillus strain reduces STEC shedding in beef cattle, but does not affect performance [41], while a host Lactobacillus strain reduced pig performance after an E. coli F4 challenge which resulted in greater E. coli F4 shedding [42]. Adding to this confusion, non-host probiotics and prebiotics increase milk production [43], alleviate disease [3,4] and decrease pathogen shedding [44], but the responses are quite variable. In an effort to determine why products differ in their effectiveness, we examined the ability of a host probiotic and a non-host prebiotic to interfere with O157 STEC colonization and mycotoxin activity in vitro and in vivo [3,4]. The non-host prebiotic reduced pathogen colonization and eliminated mycotoxin activity while the host probiotic had no effect in vitro. Application of the nonhost prebiotic or the non-host prebiotic: host probiotic combination to symptomatic dairy or beef cattle stopped the development of JHS supporting the possibility that these apparent inconsistencies with responses are related to efficacy. In the current study, a non-host probiotic reduced non-O157 and O157 STEC colonization but had no effect on mycotoxin activity in vitro. Adding to previous results [3], the non-host prebiotic reduced non-O157 and O157 STEC colonization and eliminated mycotoxin activity in vitro. Application of the nonhost prebiotic: host probiotic to symptomatic goats challenged by STEC and mycotoxins stopped the symptoms and development of HE. Together, these results suggest that it is the targeting of stressor events such as pathogen and mycotoxin challenges that determine efficacy not prebiotic or probiotic origin.
Goats supplemented with yeast probiotics show improvements in milk production but the results are quite variable [15-20]. These studies indicated that stronger responses to prebiotic or probiotic treatments were achieved with animals that were challenged by multiple factors such as heat [45] and feed quality [17]. In the current study, impaired rumen function was detected as a lack of improvement in body condition or milk production after replacement of the mycotoxin contaminated diet with the commercial diet. As evidenced in earlier in vitro studies [17,46], the current study suggests that the application of an efficacious probiotic or prebiotic reversed the impaired rumen function in the goats leading to weight gain and higher milk production.
Mastitis is a common symptom for dairy cattle exposed to mycotoxins [47] which results from mycotoxin-induced immune suppression [48]. To date, most reports for prebiotics or probiotics improving specific ruminant diseases are indirect observational data that were not a part of the experimental design. The high somatic cell counts documented in the dairy goats at the beginning of this study indicated that the herd had a high incidence of clinical mastitis (25%). Application of the prebiotic or probiotic caused a significant decline in somatic cell counts but also eliminated clinical mastitis cases over a short time period supporting an indirect benefit for the immune system. Future studies should examine the underlying mechanisms responsible for the prebiotic and probiotic decline in mastitis.
The study conducted on goats indicates that mycotoxins and STEC infections are part of the disease complex for HE during the winter months when animals are consuming mouldy feeds. The disease complex is the same as described for JHS in cattle. Application of a prebiotic: probiotic treatment alleviated morbidity while application of a prebiotic or probiotic alleviated production losses associated with the mycotoxicosis in dairy goats.
Special thanks to Ray Roberts DVM for the many discussions regarding pathology and haemorrhagic diseases in cattle. We gratefully acknowledge Keith A. Seifert and Dr. Tharcisse Barasubiye from the National Fungal Identification Service (Agriculture and Agri-Food Canada, 960 Carling Avenue, Ottawa, Ontario) for continued confirmation of fungal isolates from field samples. This research was supported by a grant from The Growing Forward Fund (Risk Mitigation Strategies Initiative RBPI # 1366) from Agriculture and Agri-Food Canada.