Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2016) Volume 0, Issue 0
Fluorochrome banding pattern with CMA and DAPI showed variation in the type, amount and distribution of heterochromatin in all the Curcuma species studied. The study provided clear insight on the numbers and genomic components such as the repetitive (GC, AT and GC/AT-rich) and non-repetitive DNA fraction(s). The polymorphism in heterochromatic-rich repetitive fraction(s) reflects higher molecular organization of the genome and evolutionary status of the genus Curcuma. The present works entails the importance of heterochromatin rich-repetitive DNA fraction(s) which play significant role in genome organization and evolutionary pattern in the genus Curcuma.
Keywords: Curcuma; Heterochromatin; Repetitive DNA fraction(s); CMA; DAPI
CMA: Chromomycin A3; DAPI: 6-diaminido-2- phenyliodole
The genus Curcuma L. (Zingiberaceae) comprises of about 120 species [1], which are rhizomatous perennial herbs. Many of them are consumed/utilized for their utility as spices, medicines, dyes, cosmetics and starch yielding taxa [2-4]. It is widely distributed in the tropical and subtropical regions of Asia [5]. The genus Curcuma display great quantum of diversity in habitat, morphology and ethno botanical aspects [6]. From the systematic point of view, the genus Curcuma is highly challenging due to complete lack of descriptors for identification of the taxa, non-availibility of herbarium specimens and also due to ambiguity in morphological markers due to extensive interspecific hybridization in the genus Curcuma [7,8]. Polyploidy has further complicated our understanding about the genome and species relationship in the genus Curcuma resulting in the taxonomic conflict between many researchers. An increase in ploidy level is commonly associated with blurring of morphological boundaries between taxa [9], which is clearly visible in the genus Curcuma.
Cytogenetical information such as chromosome number, structure, morphology, behaviour of chromosomes during mitotic and meiotic divisions, distribution of heterochromatin content have been of considerable valuable for solving taxonomic riddles on one hand and issues related to systematics, evolution and delimitation of the taxa on the other [10,11]. These parameters are also of immense value for defining intrageneric relationships in the genus Curcuma [1,3,12-14]. Such studies in the genus Curcuma are limited to chromosome counts perhaps due to their small size and higher numbers. It is interesting to note that even molecular details such as composition and distribution pattern of heterochromatin, which are generally useful for such chromosomally problematic genera, are also lacking in the genus Curcuma. The information regarding heterochromatic regions are cytologically recognized due to its condensation and differential staining during cell division [15]. Quantitative and qualitative differentiation of the heterochromatin played significant role in understanding the molecular composition of the genome without the need of advanced molecular techniques [16].
Highly repetetive DNA sequences are the main component of the heterochromatin of higher eukaryotic genomes [17]. It is mostly enriched at the centromeric and telomeric regions of the chromosomes, depending on the species or group of species [15],which contains satellite DNA, retrotransposons and/or rDNA sequences. Heterochromatin also serves a number of other important cellular and biological roles during recombination and segregation of chromosomes in meiosis/mitosis, and global control of gene regulation and speciation [18]. Heterochromatic regions have been studied by either C banding techniques or direct staining with base-specific fluorochromes [19]. The most frequently used fluorochromes are chromomycin A3 (CMA) and 6-diaminido-2-phenyliodole (DAPI) which preferentially stains GC-rich and AT-rich regions respectively on the chromosomes [20,21], to distinguish heterochromatin from euchromatin and to permit a direct comparison of the relative GC or AT content in the chromatin fraction [22]. Thereforeit helps in identification and measurement of difference in the total amount of heterochromatin and wasextensively used for solving the taxonomic problem in several species of Crinum [23,24]and different forms of Lasia [25].
Although fluorochromes binding with CMA and DAPI have been reported in several plant genera, however, no such reports are available for the genus Curcuma.Thepresent study was conducted to determinethe distribution pattern of AT- and GC- rich regions (heterochromatin) in Curcuma species. Comparative analysis of the repetitive and non-repetitive DNA fractions may also provide useful information to understand the mechanisms underlying the chromosomal evolution in the genus. Such useful information will facilitate comparative cytogenomics and provide an insight into the patterns of chromosomal differentiation of the genus Curcuma.
Plant materials
The rhizomes of 15 Curcuma species used in the present investigation were obtained from Indian Institute of Spices Research, Kozhikode, Kerala (Table 1). The plants were grown and maintained in poly-house conditions at Department of Biotechnology and Bioinformatics, North- Eastern Hill University, Shillong, Meghalaya.
| S No | Species | ISSR voucher numbers | CMA+ | DAPI+ | CMA+/DAPI+ | % Repetitive | Total % Repetitive | % Non-repetitive | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Range | Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | CMA+ | DAPI+ | CMA+/DAPI+ | |||||
| GC % | AT % | GC+AT % | |||||||||||
| 1 | C. amadaRoxb. | 521 | 4 | 4 ± 0.0 | - | - | - | - | 16.00 | - | - | 16.00 | 84.0 |
| 2 | C. aromaticaSalisb. | 711 | 4–6 | 4.28 ± 0.6 | 4 | 4 ± 0.0 | - | - | 17.12 | 16.00 | - | 33.12 | 66.88 |
| 3 | C. comosaRoxb. | 644 | 2–7 | 4.35 ± 1.4 | - | - | 3 | 3 ± 0.0 | 17.40 | - | 12.00 | 29.4 | 70.6 |
| 4 | C. harithaMangaly& M. Sabu | 1136 | 4–5 | 4.2 ± 0.4 | - | - | - | - | 16.80 | - | - | 16.80 | 83.2 |
| 5 | C. manggaValeton&Zijp | 1049 | 4 | 4 ± 0.0 | 4–7 | 5.72 ± 1.1 | 5 | 5 ± 0.0 | 16.00 | 22.90 | 20.00 | 58.9 | 41.1 |
| 6 | C. montanaRoxb. | 649 | 4 | 4 ± 0.0 | 4–5 | 4.24 ± 0.4 | - | - | 16.00 | 16.97 | - | 32.97 | 67.03 |
| 7 | C. aeruginosaRoxb. | 1144 | 6–8 | 6.48 ± 0.6 | 7 | 7 ± 0.0 | 2 | 2 ± 0.0 | 25.90 | 28.00 | 8.00 | 61.9 | 38.1 |
| 8 | C. caesiaRoxb. | 700 | 6–8 | 6.48 ± 0.6 | - | - | 5 | 5 ± 0.0 | 25.90 | - | 20.00 | 45.9 | 54.1 |
| 9 | C. latifoliaRoscoe | 638 | 5–6 | 5.12 ±0.3 | 4–6 | 5.44 ±0.8 | - | - | 20.50 | 21.76 | - | 42.26 | 54.24 |
| 10 | C. leucorrhizaRoxb. | 1169 | 6 | 6 ± 0.0 | - | - | 4–6 | 5.44 ± 0.6 | 24.00 | - | 21.76 | 45.76 | 48.24 |
| 11 | C. longa L. | Pratibha | 4–6 | 4.24 ± 0.7 | - | - | 2 | 2 ± 0.0 | 16.96 | - | 8.00 | 24.96 | 75.04 |
| 12 | C. sylvaticaValeton | 526 | 5–6 | 5.08 ± 0.3 | 8 | 8 ± 0.0 | - | - | 20.32 | 32.00 | - | 52.32 | 47.68 |
| 13 | C. zanthorrhizaRoxb. | 1108 | 3–4 | 3.48 ± 0.5 | 3–5 | 3.6 ± 0.8 | 4 | 4 ± 0.0 | 13.92 | 14.40 | 16.00 | 44.32 | 55.68 |
| 14 | C. zedoaria (Christm.) Roscoe | 760 | 4–6 | 5.12 ± 1.0 | 0–8 | 3.28 ± 2.4 | 3 | 3 ± 0.0 | 20.48 | 13.12 | 12.00 | 45.6 | 54.4 |
| 15 | C. raktakantaMangalyand M. Sabu | 1137 | 4–8 | 4.96 ± 0.7 | 4–8 | 5.72 ± 1.5 | - | - | 19.84 | 22.90 | - | 42.74 | 57.26 |
Table 1: Specimen voucher and comparative heterochromatin banding pattern in Curcuma species.
Chromosome preparation
Actively growing root tips of 1-2 cm long were excised from field grown plants and were pretreated with saturated para-dichlorobenzene (HiMedia) for 3 hrs at room temperature and were subsequently fixed in freshly prepared 1:3 acetic acid/ethanol mixture for a minimum period of 24 hrs and thereafter stored in 70% ethanol.
CMA/DAPI staining
Fluorescent banding using CMA (excitation/emission wavelengths: 365-460/475-680 nm) and DAPI (excitation/emission wavelengths: 320-385/400-580 nm) which binds specifically to GC and AT - rich regions respectively, was performed according to method described by Schweizer and Ambros [26] with minor modifications. The fixed root tips digested in an enzymatic solution containing 2% cellulase (Onuzuka) and 2% pectolyase (Sigma) at 37°C for 30-60 mins. The hydrolyzed root tips were squashed in 45% acetic acid. Slides were frozen in liquid nitrogen and aged for 3 days at room temperature in the dark. The aged slides were stained with CMA (0.5 mg/ml) for 60 min and subsequently counterstained with DAPI (2 μg/ml) for 30 min. The slides were then mounted with an antifade solution containing in 1:1 (v/v) McIlvaine’s buffer (pH 7.0) supplemented with 2.5 mM MgCl2 and glycerol and kept for 2 days at 4°C for aging before observation under a fluorescence microscope with a different filter cassette for CMA and DAPI respectively. Photomicrographs for CMA/DAPI stained preparations were captured using Leica DFC310FX camera (Germany) attached to Leica Microsystems CMS GmbH fluorescent microscope at × 100 magnifications. An average of 25 cells was used in the present analysis.
The present investigations revealed differential heterochromatin banding pattern in 15 Curcuma species. Detailed analysis using CMA/DAPI staining showed constitutive heterochromatin as bright green fluorescence scored as CMA+ (CMA positive), while bright blue fluorescence as DAPI+ (DAPI positive) and CMA+/DAPI+ (CMA and DAPI positive) where both CMA and DAPI show intermingled fluorescence signals and the data are illustrated in Figures 1-4 and summarized in Table 1.
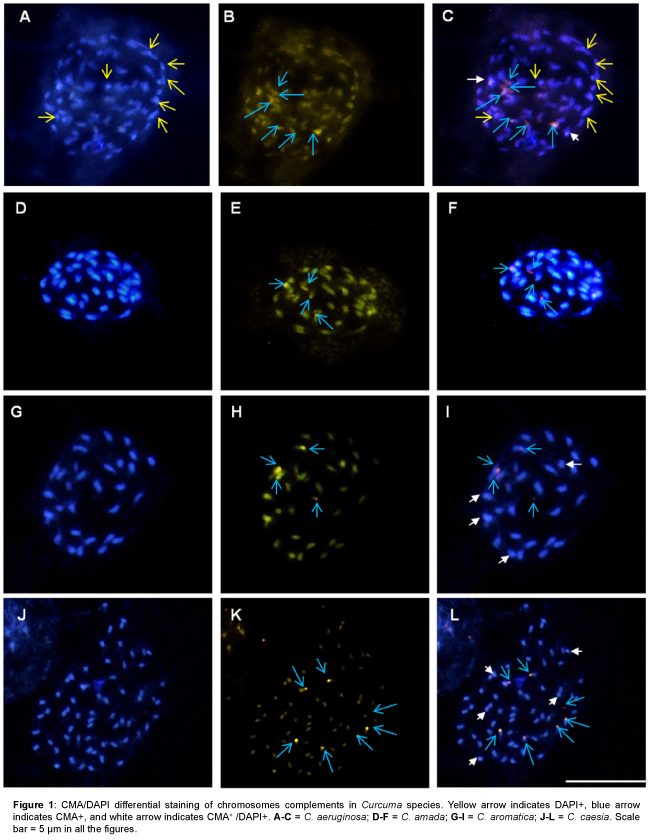
Figure 1: CMA/DAPI differential staining of chromosomes complements in Curcuma species. Yellow arrow indicates DAPI+, blue arrow indicates CMA+, and white arrow indicates CMA+ /DAPI+. A-C = C. aeruginosa; D-F = C. amada; G-I = C. aromatica; J-L = C. caesia. Scale bar = 5 μm in all the figures.
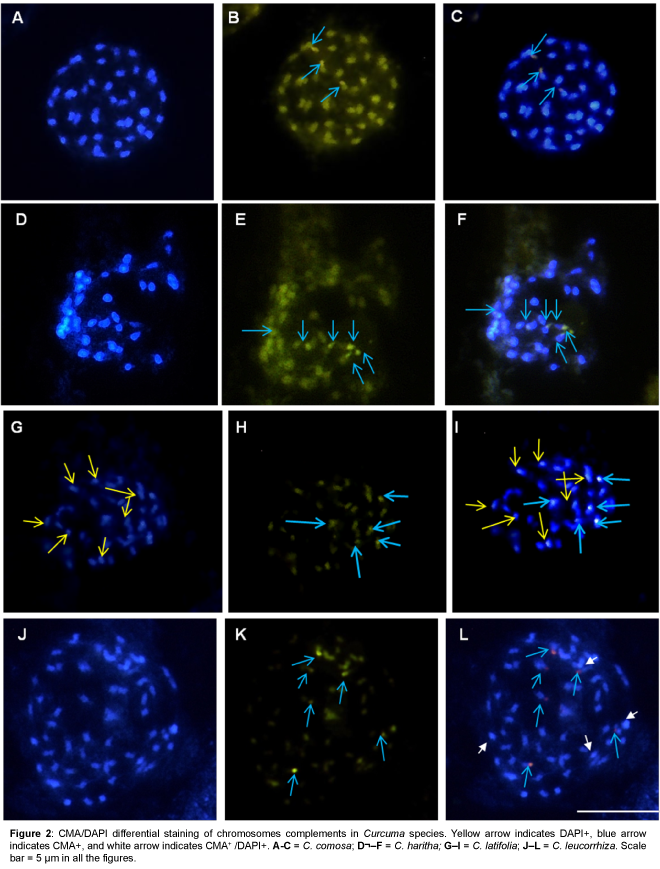
Figure 2: CMA/DAPI differential staining of chromosomes complements in Curcuma species.Yellow arrow indicates DAPI+, blue arrow indicates CMA+, and white arrow indicates CMA+ /DAPI+. A-C = C. comosa; D–F = C. haritha; G–I = C. latifolia; J–L = C. leucorrhiza. Scale bar = 5µm in all the figures.
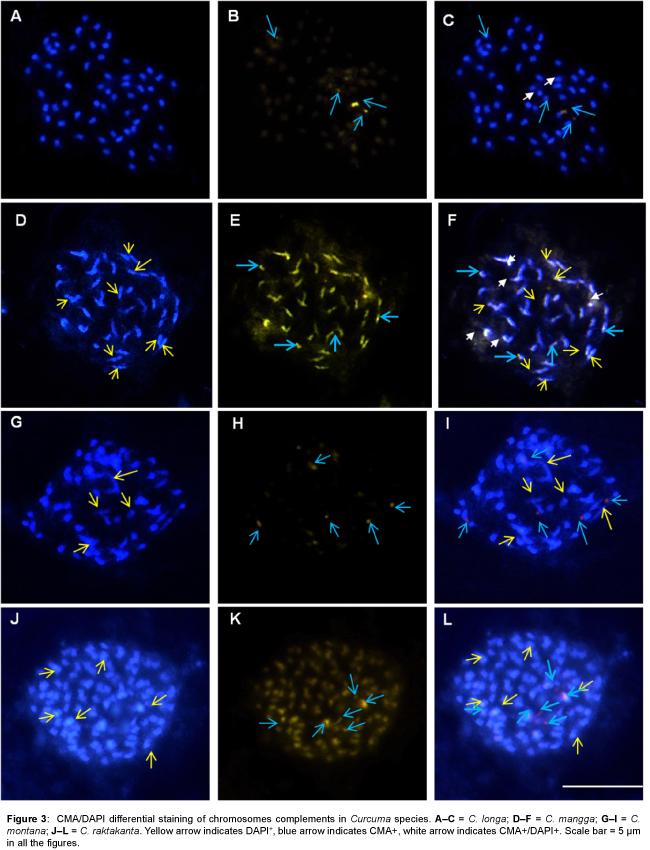
Figure 3: CMA/DAPI differential staining of chromosomes complements in Curcuma species. A–C = C. longa; D–F = C. mangga; G–I = C. montana; J–L = C. raktakanta. Yellow arrow indicates DAPI+, blue arrow indicates CMA+, white arrow indicates CMA+/DAPI+. Scale bar = 5 μm in all the figures.
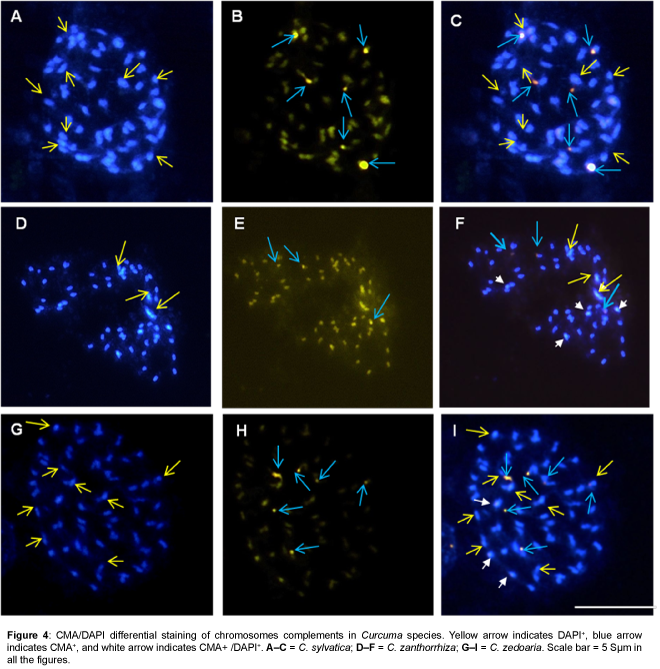
Figure 4: CMA/DAPI differential staining of chromosomes complements in Curcuma species. Yellow arrow indicates DAPI+, blue arrow indicates CMA+, and white arrow indicates CMA+ /DAPI+. A–C = C. sylvatica; D–F = C. zanthorrhiza; G–I = C. zedoaria. Scale bar = 5 μm in all the figures.
All species showed the presence of distinctive CMA+ banding pattern both at interphase and metaphase chromosomes. In most species CMA+ heterochromatin were located at the distal end of the chromosomes, however, one interstitial CMA+ heterochromatin region was also recorded in C. aromatica (Figure 1H), C. longa (Figure 3B), C. mangga (Figure 3E), C. caesia, C. comosa and C. leucorrhiza. In contrast, DAPI+ and CMA+/DAPI+ heterochromatin bands were mostly confined to the interstitial region (Figures 1C, 2G, 3G, 3F and 4G) of the chromosomes which varied in the number, size and location of the bands among the taxa.
A characteristic feature observed was the differential distribution of GC/AT- rich heterochromatin sites in the genome. The number of CMA+ binding sites ranged from 3.48 ± 0.5 (C. zanthorrhiza) to 6.48 ± 0.6 (C. aeruginosa and C. caesia) whereas that of DAPI+ sites ranged from 3.2 ± 2.5 (C. zedoaria) to 8 ± 0 (C. sylvatica) and that of CMA+/ DAPI+ ranged from a minimum of 2 ± 0 (C. aeruginosa and C. longa) to a high of 5 ± 0 (C. caesia and C. mangga).
Another distinctive feature of the heterochromatin distribution pattern is the amount of repetitive and non-repetitive fraction(s) of the DNA (Table 1). The total repetitive fraction(s) ranged from 16% (C. amada) to 61.9% (C. aeruginosa) which corresponds to GC-rich (16%, 25.90%), AT-rich (0%, 28%) and/or AT+GC rich (0%, 8%) respectively. It can be noted that the amount of AT-rich fraction(s) is slightly higher compared to the GC-rich fraction(s) in C. aeruginosa, C. aromatica, C. latifolia, C. mangga, C. raktakanta, C. sylvatica, and C. zanthorrhiza exception is the case of C. zedoaria where the GC-rich fraction(s) (20.48%) is more than as compared to AT-rich fraction(s) (13.12%). The non-repetitive DNA fraction(s) ranged from a minimum of 38.1% (C. aeruginosa) to 84% (C. amada).
Fifteen species of Curcuma currently investigated differed 3.9- fold in the amount of repetitive and 3.8 fold in non-repetitive DNA fraction(s). This variation in the amount and type of repetitive and non-repetitive DNA fraction(s) among species may be attributed to the rapidly evolving nature of the heterochromatin [18] and might have contributed to the genome evolution in the genus Curcuma. Several hypotheses have been proposed to understand the nature of the heterochromatin comprising of repetitive sequences. Among them, unequal crossing over [27], polymerase slippage or massive insertions/ removal of mobile elements [28,29] is well recognized. Alternatively, changes in the epigenetic state of the chromatin, by losses or alterations of chromatin insulators [30], or by de novo insertion of few mobile elements may determine changes in the size of heterochromatic bands [31].
Heterochromatin is one of the most important factors in evolution [32], since it can interfere with phenomena such as DNA replication, chromosomal structure, gene expression and the cell cycle [9,33]. DAPI+ or CMA+ bands observed in many plant genera reveal the presence of AT- or GC-rich repetitive DNA sequences, respectively [19,34]. Hence, CMA positive (CMA+) segment is high in GC-content and DAPI positive (DAPI+) segment is high AT-content. Comparative studies of plant repetitive sequences are useful for investigating evolutionary relationships between plant species [35]. Banding pattern with base-specific fluorochromes (CMA and DAPI) of metaphase as well as interphase cells showed the presence of GC- and/or AT- rich heterochromatin and some consisted of both GC- and AT-rich combine repetitive sequences in all the species analyzed (Table 1; Figures 1-4).
The amount of AT-rich fraction(s) is comparatively higher as compared to the number of GC-rich heterochromatin repetitive fraction(s) with a 1.1- fold mean difference in the amount of AT to GCrich repetitive fractions. Similar observations have also been reported in other species as well [16,19,36,37]. However, a contrary status has also been reported in some species [38], which is also the case of C. zedoaria presently studied. Though the polymorphism can be observed in the amount, type and distribution of heterochromatin but the general patterns were relatively unique for each species studied as reported in Vigna [39,40], Citrus [41], Cestrum [42] and Abelmoschus [16].
Heterochromatin studies in the genus Curcuma provide clear information on genome constitution such as the repetitive sequence (GC, AT and GC/AT-rich) families. The variation in the amount and distribution of heterochromatin are characteristics with highest taxonomic value at the species level [43], leading to greater genome size [1]. The polymorphism of the heterochromatic-rich repetitive fractions advocates their important role in genome structure and function of the genus Curcuma. This variation reflects higher molecular organization of the genome and polyploidy might have contributed significantly to the species diversification. The repeated DNA sequences of the heterochromatin are subjected to rapid evolutionary changes [17]. Therefore, it may be opined that the species within the genus Curcuma may have different evolutionary pattern which was also supported by the findings from genome size estimation by Škorničková et al. [1]. The present work entails the importance heterochromatin rich-repetitive DNA sequences which can give significant insights into the genome structure and should be considered a useful tool for understanding the genome organization and evolutionary drift in the genus Curcuma. Fluorescent in situ hybridization (FISH), a powerful tool for chromosome analysis, may authentically substantiate genome structure, organization and distribution of repetitive DNA families, and could resolve some of the unknown aspects of polyploidy in the genus Curcuma.
This work was financially supported by DBT-JRF Program, Department of Biotechnology, Government of India (Fellowship Ref. No. DBT-JRF/2010-11/582 dated 27th January, 2011). The present work was carried out in the Department of Biotechnology and Bioinformatics, North-Eastern Hill University, Shillong. The authors are thankful to the Head of the Department for providing necessary facilities. The authors are also grateful to the Director, Indian Institute of Spices Research (IISR), Kozhikode for providing the germplasm, VA Parthasarathy and Dr. Prasath Duraisamy for their help and encouragement. Sincere thanks are also due to the members of Plant Biotechnology Laboratory, Department of Biotechnology and Bioinformatics, North-Eastern Hill University, Shillong for their constant help and encouragement.