Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Hypothesis - (2014) Volume 2, Issue 3
Terrestrial plants lack inositol-1,4,5-trisphosphate (IP3) receptor regulating transient Ca2+ increase to activate cellular Ca2+-dependent physiological events. To understand an evolutional route of the loss of the IP3 receptor gene, conservation of the IP3 receptor gene in algae was examined in silico based on the accumulating information of genomes and expression sequence tags. Results clearly demonstrated that the lack of the gene was observed in Rhodophyta, Chlorophyta except for Volvocales and Streptophyta. It was therefore hypothesized that the plant IP3 receptor gene was eliminated from the genome at multiple occasions; after divergence of Chlorophyta and Rhodophyta and of Chlorophyta and Charophyta.
Keywords: Alga; Ca2+; Comparative genomics; Gene; Inositol-1,4,5-trisphosphate receptor
DAG: Diacylglycerol; IP3: Inositol-1,4,5- Trisphosphate; IP6: Inositol-1,2,3,4,5,6-Hexakisphosphate, PI-PLC: Phosphoinositide-Specific Phospholipase C, PKC: Protein Kinase C.
Inositol-1,4,5-trisphosphate [Ins(1,4,5)P3, IP3] is a second messenger involved in transient release of Ca2+ from the ER that activates cytosolic Ca2+ signalling cascades in response to extracellular and intracellular stimuli [1,2]. Phosphatidylinositol-4,5-bisphosphate is cleaved by phosphoinositide-specific phospholipase C (PI-PLC) into the second messengers diacylglycerol (DAG) and IP3 [3,4]. These second messengers then activate protein kinase C (PKC) and the ER-localised IP3 receptor, respectively, in animal cells [1,2]. However, although the PI-PLC signaling cascade is present in plants [5-7], genes encoding PKC and the IP3 receptor have not been found in terrestrial plant genomes, suggesting differences in second messenger systems between animals and plants. To date, the genomes of a variety of unicellular and multicellular algae have been sequenced [8-23] as shown in (Table 1). In addition, large-scale EST information for the red seaweeds Porphyra umbilicalis and Porphyra purpurea has been accumulated [24-26]. Such rich gene information enables us to identify the genes encoding IP3 receptor gene homologues in algae to hypothesize the evolutionary route of the loss of the IP3 gene in plant lineages.
| Phylum | Class | Order | Family | Species | Ref |
|---|---|---|---|---|---|
| Chlorophyta | Prasinophyceae | Mamiellales | Mamiellaceae | Ostreococcustauri | [8] |
| Ostreococcuslucimarinus | [9] | ||||
| Chlorophyceae | Volvocales | Chlamydomonadaceae | Chlamydomonasreinhardtii | [10] | |
| Volvocaceae | Volvoxcarteri | [11] | |||
| Chlorococcales | Coccomyxaceae | Coccomyxasubellipsoidea | [12] | ||
| Rhodophyta | Cyanidiophyceae | Cyanidiales | Cyanidiaceae | Cyanidioschyzonmerolae | [13] |
| Galdieriasulphuraria | [14] | ||||
| Porphyridiophyceae | Porphyridiales | Porphyridiaceae | Porphyridiumpurpureum | [15] | |
| Rhodophyceae | Bangiales | Bangiaceae | Pyropiayezoensis | [16] | |
| Florideophyceae | Gigartinales | Gigartinaceae | Chondruscrispus | [17] | |
| Glaucophyta | Glaucophyceae | Glaucocystales | Glaucocystaceae | Cyanophoraparadoxa | [18] |
| Heterokontophyta | Coscinodiscophyceae | Thalassiosirales | Thalassiosiraceae | Thalassiosirapseudonana | [19] |
| Bacillariophyceae | Naviculales | Phaeodactylaceae | Phaeodactylumtricornutum | [20] | |
| Pelagophyceae | Pelagomonadales | Pelagomonadaceae | Aureococcusanophagefferens | [21] | |
| Phaeophyciae | Ectocarpales | Ectocarpaceae | Ectocurpussiliclosus | [22] | |
| Charophyta | Klebsormidiophyceae | Klebsormidiales | Klebsormidiaceae | Klebsormidiumflaccidum | [23] |
Table 1: List of algal species whose genome sequences have been analyzed.
The origin of the IP3 receptor-dependent transient Ca2+ release system predates the divergence of animals and fungi [27,28]. Indeed, homologues of genes encoding the IP3 receptor have been identified in protozoa such as the choanoflagellate Monosiga brevicollis [29], the myxomycete Dictyostelium discoideum [30], the ciliate Paramecium tetraurelia [31], and the parasite Trypanosoma brucei [32]. Thus, it is plausible that an ancient eukaryotic cell containing an IP3 receptor gene was the target of endosymbiosis with an ancient cyanobacterium to produce plant cells, after which the IP3 gene was lost from plant lineages. At present, IP3 receptor homologues have been found in green algae, such as Chlamydomonas reinhardtii [10] and Volvox carteri [33,34], and in heterokont algae including Aureococcus anophagefferrens [21] and Ectocarpus siliculosus [22], but have not been identified in red algae or streptophytes (land plants and charophytic algae) (Figure 1). These findings have led to proposals that the IP3 receptor gene homologue was lost on multiple occasions during plant evolution. Because an ancestor of both green and red photosynthetic algal cells appeared after the primary endosymbiosis of a cyanobacterium into an ancient non-photosynthetic eukaryotic cell [35], the IP3 receptor homologue was probably lost from lineages of red algae and green algae except for Volvocales (Figure 1). In fact, the genomes of unicellular Aureococcus anophagefferrens and multicellular Ectocarpus siliculosus carry an IP3 receptor gene homologue (Figure 1). Because both photosynthetic algae arose from secondary endosymbiosis of a red algal cell into an ancient non-photosynthetic eukaryotic cell [35], it appears that red algae subsequently lost the IP3 receptor gene homologue during their evolution, although some of Heterokontophyta that evolved by secondary symbiosis retain an ancient progenitor of the IP3 receptor gene to this date. Moreover, in the green plant lineage, streptophytes have an impaired IP3 receptor that is structurally similar to that in animals, Volvocales of chlorophytes, and brown seaweed (Figure 1). Thus, the loss of the IP3 receptor may also occurred after the divergence of chlorophytes and streptophytes. Accordingly, there have been multiple occasions upon which the IP3 receptor was lost from plant lineages. In contrast to the above conclusions drawn from genomic sequence information, there is evidence of IP3-dependent Ca2+ release in terrestrial plants [36-42], which suggests the presence of a Ca2+ channel functionally resembling the IP3 receptor in streptophytes. However, IP3-dependent Ca2+ release has been reported only in green algae among plants [43,44]. Because the major intracellular store of Ca2+ in plant cells is the vacuole [45,46], IP3 receptor activity is thought to be localised to vacuolar membranes in green algae and streptophytes. Such is the case in the fungus Neurospora crassa, in which IP3-mediated Ca2+ release occurs from vacuoles [47], as it also does in protozoan ciliates and trypanosomes, in which the IP3 receptor has been visualized on vacuolar membranes [27,28]. Thus, the green plant lineage has maintained an ancient system for transient release of Ca2+ from vacuoles, which is distinct from ER-mediated Ca2+ release in animal cells that do not possess vacuoles.
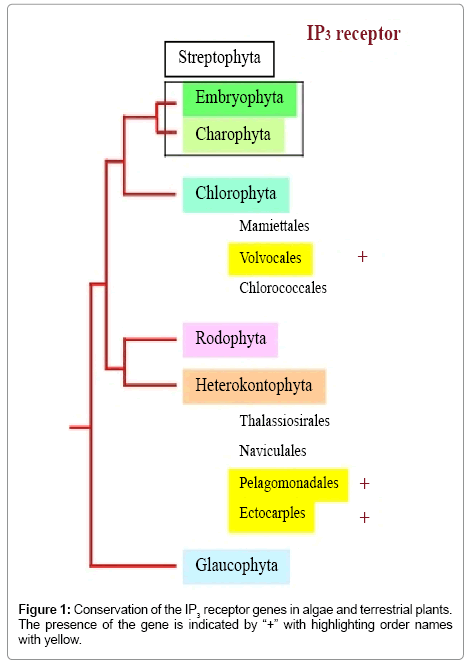
Figure 1: Conservation of the IP3 receptor genes in algae and terrestrial plants. The presence of the gene is indicated by “+” with highlighting order names with yellow.
IP3-mediated Ca2+ release has been observed at the ER membrane that predominates in the perinuclear and apex regions of cells of the brown seaweed Fucus serratus [48]. Because an IP3 receptor homologue was found in the Ectocarpus siliculosus genome [22], it is possible that the location of the IP3-sensitive Ca2+ store shifted from the vacuole to the ER in brown seaweeds, where it is currently found in animal cells. Thus, the brown seaweeds might possess a PI-PLC signaling system more similar to that in animals.
Although IP3-mediated Ca2+ release has not yet been shown in red algae, an inhibitor of the IP3-receptor, 2-APB, prevented establishment of cell polarity for the migration and germination of monospores in the red seaweed Pyropia yezoensis [49], which suggests the presence of an IP3 receptor-mediated Ca2+ release system in red seaweeds. However, an IP3-receptor homologue has not yet been identified in the Pyropia yezoensis genome. As there is currently no evidence indicating the presence of IP3 in Pyropia yezoensis, biochemical determinations of this inositol derivative will be necessary to elucidate Ca2+ release upon PIPLC action in red algae.
In plant cells, DAG is usually phosphorylated by DAG kinase [50,51] to produce phosphatidic acid, and IP3 is phosphorylated by inositol phosphate kinases, IPK1 and IPK2 [52,53] to produce inositol- 1,3,4,5,6-pentakisphate and inositol-1,2,3,4,5,6-hexakisphosphate [Ins(1,2,3,4,5,6)P6; phytate; IP6], a high-abundance molecule that is considered important for phosphorus storage in plant cells. To date, PA and IP6 are thought to act as major second messengers in plant cells [7,54], although the function of IP3 as a second messenger in plants has not been ruled out [42,47]. For instance, Munnik and Vermeer [54] have proposed that IP6, which is rapidly converted from IP3, is a major second messenger involved in abscisic acid-dependent inhibition of stomatal opening. They have also proposed a parallel between the IP3 and IP6 signalling systems because these two molecules are both produced by the action of PI-PLC. Although neither an IP3 nor an IP6 receptor have yet been identified in terrestrial plants, it is possible that an IP3 receptor or an IP6 receptor of unknown structure is present in streptophytes. Taken together, comparative genomic information clearly demonstrates the loss of the IP3 receptor gene in red algae, green algae except for Volvocales and streptophytes during plant evolution. However, IP3-dependent transient Ca2+ release from intracellular stores has been shown in these organisms by physiological experiments, although whether plants lacking the IP3 receptor might both possess a common system for such transient Ca2+ release is uncertain. Therefore, the identification and characterization of genes encoding putative IP3 or IP6 receptors of unknown structure is of the highest priority for elucidating and comparing the regulation of the PI-PLC signalling cascade between IP3 receptor-carrying and -lacking algae.