Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2014) Volume 3, Issue 4
Calbindin-D28K is a calcium sensor protein responsible for maintaining cellular calcium homeostasis and is known to have anti-apoptotic properties. The goals of this study were to propose calbindin-D28K peptide inhibitors based on known protein:protein interactions, in particular calbindin-D28K’s interaction with caspase-3. A total of 160,000 potential cyclic peptide inhibitors were computationally screened against calbindin-D28K. Several scoring mechanisms were used to validate each cyclic peptide inhibitor:calbindin-D28K interaction. A general consensus sequence of the cyclic peptides was shown to contain positive electrostatic characteristics with at least one aromatic-containing amino acid. Binding for one of the best scoring cyclic peptides is subsequently shown to alter the caspase-3 binding site on calbindin-D28K. Overall, these results suggest that calbindin-D28K is a druggable target via cyclic peptide inhibitors ultimately leading to the reactivation of caspase-3 and natural cell death.
<Chemotherapy and radiotherapy induce tumor cell death by activating apoptosis (programmed cell death), a cellular pathway inactive in tumor cells. Any mechanism that inhibits apoptosis makes tumor cells resistant to these treatments. Therefore, strategies that sensitize tumor cells by targeting/enhancing the apoptotic pathway are of enormous interest. Effective therapeutic options are ones that also must affect tumor cells without compromising normal cell function as much as possible. Consequently, it is important to define a target that (i) has significant, well-established, anti-apoptotic function in the cell and (ii) is known to be expressed at higher levels in cancerous cells than normal cells. Considering all of these points, the anti-apoptotic calcium-binding protein calbindin-D28K is an especially appealing target.
It is well known that small cell lung cancer and non-small cell lung cancer cells have significantly higher concentration of calbindin- D28K compared to normal lung extracts. In addition, the non-small cell lung cancer cells in these studies are also seen to appear at an advanced cancer stage [1,2]. High expression levels of calbindin-D28K have also been observed in medulloblastomas, a highly malignant brain tumor predominantly affecting children under 10 years of age [3,4]. Overexpression of calbindin-D28K protects osteocytics and osteoblastic (bone) cells from induced apoptosis [5]. Furthermore, a human endometrial adenocarcinoma cell line that overexpresses calbindin-D28K was also shown to inhibit cell death and decrease the amount of Bax, p53 and caspase-3 expression during H2O2-induced apoptosis [6]. Several other proteins, including members of the Bax, p53, Bcl-2 and caspase-3 protein families regulate the apoptosis mechanism. These studies indicate that calbindin-D28K is present at higher levels in tumor cells than normal cells and also has an important role in blocking the up-regulation of apoptosis in many cancer cell lines.
Calbindin-D28K achieves its anti-apoptotic effects through direct inhibition of the pro-apoptotic ‘cell executioner’ protein, caspase-3 [7-9]. The interaction between the calcium-loaded form of calbindin-D28K and caspase-3 has been previously characterized using isothermal titration calorimetry (ITC) and molecular docking. These studies provided the first insights into the mechanism of caspase-3 inhibition [8]. Calbindin-D28K’s interaction with linear peptides has also been characterized using circular dichroism, nuclear magnetic resonance (NMR) and molecular modeling [10]. Affecting the ability of calbindin-D28K to bind caspase-3 will induce apoptosis – helping overcome resistance to chemo- and radiation treatments.
Druggability assessment of a target protein - the likelihood of being able to regulate protein:protein interactions with a therapeutic - is an important stage in the development of new therapeutics [11]. Here we show the druggablity of calbindin-D28K using a library of 160,000 proposed cyclic peptide inhibitors (CX1X2SIKNX3X4C). We have chosen to target calbindin-D28K with cyclic peptides. (1) Cyclic peptides are well-established therapeutics, e.g. actinomycin D (cancer chemotherapeutic), cyclosporin A (immunosuppressant), oxytocin (labor induction), polymyxin E (antibiotic) and octreotide (growth hormone antagonist). (2) Cyclic peptides overcome many cell permeability issues compared to linear peptides. It has been shown that cyclization of linear peptides can increase their lipophilicities, thereby increasing their permeation characteristics. (3) Since the cyclic peptide is somewhat constrained, there is little entropic penalty to the binding compared to a linear peptide, which has to 'freeze out' superfluous conformations in order to obtain optimal binding - a pre-organized cyclic conformation frequently exhibits notably better affinity for its target. (4) Cyclic peptides are more metabolically stable, being more resistant to digestion/degradation (particularly to hydrolysis by exoproteases since they lack both amino and carboxyl termini). This significantly increases bioavailability. (5) The cyclic peptide motif is much more amenable to computational modeling and structure-based analog design [12-23].
Here in our present study, we show that of the potential druggable sites within calbindin-D28K, the highest affinity location for the cyclic peptides corresponds to the area where linear peptides have previously been shown to bind (region of EF-hand 2) [8]. This location is immediately adjacent to the site where caspase-3 interacts with calbindin-D28K [10]. We also identify a general consensus sequence of potential cyclic peptide inhibitors of calbindin-D28K. This consensus contains positive electrostatic characteristics with at least one aromaticcontaining amino acid on opposite sides of the SIKN motif. Further computational analyses also confirm the ability of these cyclic peptides to alter the surface and dynamics characteristics of the calbindin- D28K:caspase-3 binding site. Such alterations would theoretically impede caspase-3 interactions and re-potentiate the apoptotic process as required.
Druggability assessment was performed through the ProDY/ DruGUI plugin for VMD [11,24]. Briefly, calbindin-D28K PDB was solvated in a system containing water, counter ions, as well as probe molecules isopropanol, acetamide and acetate and isopropylamine in a ratio of 70:10:20. A simulation box of water with padding at 6 Å was constructed. Molecular dynamic simulations were then performed through NAMD [25]. The small molecules were allowed to interact with calbindin-D28K. Through this method the surfaces of calbindin- D28K that bind a wide range of fragments usually coincide with known druggable sites. The analysis steps involved selecting high-affinity probe binding spots, clustering them, and then merging the data into a sum of their binding free energies.
Docking experiments were performed using automated docking program AUTODOCK (v.4.2.5.1) [26]. Before the actual docking process, the peptide ligand library containing 160,000 different peptide sequences was constructed using PyMOL and Crystallography and NMR System (CNS) with the basic sequence of CX1X2SIKNX3X4C, where X denotes the variable amino acid positions [27-29]. PyMOL was used to construct the linear ligand PDBs [29]. CNS was used to cyclize the linear PDBs through the N- and C-terminal cysteine residues, i.e. formation of a disulfide bond [27,28]. Ligand and receptor preparation was performed involving addition of hydrogen atoms and computing charges. The rotatable torsion bond information for each cyclic peptide was acquired defining the bond flexibility parameters. The receptor preparation was performed through Auto-Grid and was used for grid map calculations that identified atom types of the ligand, which acted as probes. The grid was a volume 60×60×54 Å with a spacing of 0.375 Å centered on the linear peptide receptor-binding site of calbindin-D28K. Then the docking calculations were performed using the genetic algorithm (GA) procedure with default parameters: ga_num_evals=1000000 - maximum number of energy evaluations, ga_pop_size=100 - number of individuals in population, and ga_ run=30 (number of GA runs). The following scoring methods were used; Ki (AUTODOCK), binding energy (BE – AUTODOCK), ligand efficiency (LE - AUTODOCK), liphophilic efficiency (LiPE), and ligand efficiency dependent lipophilicity (LELP). Results were visualized with PyMOL and if the cysteine residues of the cyclic peptide were primarily responsible for the interaction with calbindin-D28K, the result was discarded.
HADDOCK docking webserver (default parameters) was used to investigate the binding surface change of calbindin-D28K when bound to caspase-3 vs. a cyclic peptide [30]. To avoid repeating the full docking procedure and bias results away from those of the AUTODOCK procedure, the cyclic peptide was targeted to the binding surface as determined with AUTODOCK. The highest populated cluster based on Cα root mean square deviation (rmsd) was joined into a single ensemble for PCA analysis. 200 out of 200 modeled complexes clustered into a single cluster based on an rmsd of 7.5 Å.
PCA was performed on the structure ensembles using THESEUS [31]. PCA is a standard tool in the field of multivariate analysis for extracting from a set of interrelated variables a much smaller set that retains most of the variation contained in the full set. By setting up a correlation matrix whose elements are the ensemble average of the pairwise products of displacements from their average position of landmarks (such as the Cα positions in a protein), PCA can be very helpful in identifying, from an NMR structural ensemble, correlations in conformational rearrangements within a protein [32].
The druggablity analysis performed in this study confirms the relevance of calbindin-D28K as a target for peptide therapeutics. Calbindin-D28K was examined through the druggability plugin ProDY/DruGUI [11] using the molecular visualization program VMD. This study resulted in two high-affinity sites and several lower affinity sites (Figure 1 and Table 1). The highest affinity site corresponds to the site on calbindin-D28K where a series of linear peptides were previously shown to bind [10]. These linear peptides were derived from the following known calbindin-D28K binding proteins: Ran-binding protein M (RanBPM), myo-inositol monophosphatase (IMPase), and caspase-3/procaspase-3 [7,28]. From these linear peptides, a small pseudo-consensus sequence became apparent, SIKN. A cyclic vpeptide library was then computationally constructed based on this sequence, CX1X2SIKNX3X4C, where X denotes any of the 20 naturally occurring amino acids and targeted to the highest affinity binding site (Figure 1). The cysteine residues at the termini of each peptide are present to cyclize the peptide in the computational studies and are not meant to be significantly involved in the interaction with calbindin-D28K. Disulfide cyclized peptides are not ideal for therapeutic intervention as the disulfide bond can be reduced resulting in a linearized peptide. The disulfide linkage shown here is merely used to provide a high throughput computational screening method utilizing established computational protocols. Any derived therapeutic must consequently have this disulfide bond removed and replaced with another stable bond, i.e. a carbon-carbon bond. The library represented all sequence combinations possible, hence a total of 160,000 possibilities.
| Site | Binding Free Energy (kCal/mol) | Affinity |
|---|---|---|
| #1 | -11.65 | 3.24 nM |
| #2 | -11.05 | 8.84 nM |
| #3 | -10.04 | 47.80 nM |
| #4 | -8.29 | 900 nM |
| #5 | -7.76 | 2.22 mM |
Table 1: Druggability results
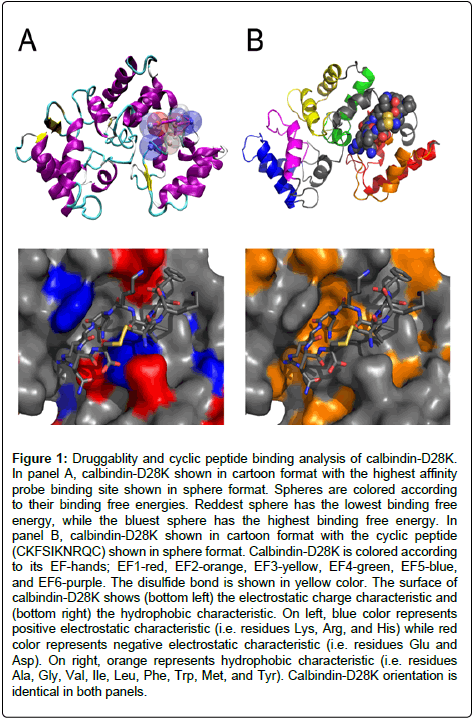
Figure 1: Druggablity and cyclic peptide binding analysis of calbindin-D28K. In panel A, calbindin-D28K shown in cartoon format with the highest affinity probe binding site shown in sphere format. Spheres are colored according to their binding free energies. Reddest sphere has the lowest binding free energy, while the bluest sphere has the highest binding free energy. In panel B, calbindin-D28K shown in cartoon format with the cyclic peptide (CKFSIKNRQC) shown in sphere format. Calbindin-D28K is colored according to its EF-hands; EF1-red, EF2-orange, EF3-yellow, EF4-green, EF5-blue, and EF6-purple. The disulfide bond is shown in yellow color. The surface of calbindin-D28K shows (bottom left) the electrostatic charge characteristic and (bottom right) the hydrophobic characteristic. On left, blue color represents positive electrostatic characteristic (i.e. residues Lys, Arg, and His) while red color represents negative electrostatic characteristic (i.e. residues Glu and Asp). On right, orange represents hydrophobic characteristic (i.e. residues Ala, Gly, Val, Ile, Leu, Phe, Trp, Met, and Tyr). Calbindin-D28K orientation is identical in both panels.
Next, each cyclic peptide was docked to calbindin-D28K at the high-affinity site identified through the druggability screen through AUTODOCK. The resulting docked solutions were analyzed through AUTODOCK python scripts revealing a hierarchy of binding based on both energy and the inhibition constant, Ki (Table 2). The top 50 solutions (out of 160,000 - 0.03%) based on the following scoring methods were visually analyzed with PyMOL for binding orientation: inhibition constant Ki (AUTODOCK); binding energy BE (AUTODOCK); ligand efficiency LE (AUTODOCK); liphophilic efficiency (LiPE = pKi-logP) and ligand efficiency dependent lipophilicity (LELP = (logP/LE)) [29]. In the resulting docked solution structures, if the disulfide bond was as a major part of the interaction with calbindin-D28K, the cyclic peptide was then subsequently removed from the analysis. Only those peptides where the disulfide bond played little to no role in the interaction were kept for further analysis example in Figure 1.
Using the employed scoring methods, there is a clear indication of the amino acid types/characteristics preferred in the cyclic peptides bound to calbindin-D28K. The scoring methods LELP and LiPE show preferences for aromatic amino acids at all four variable positions of the CX1X2SIKNX3X4C motif (Trp, Tyr, and Phe). These scoring methods also resulted in the least number of docked cyclic peptides with the disulfide bond playing no major role in the interaction with calbindin-D28K (13 out 50 and 21 out of 50, respectively). On average, the disulfide bond was 3.6 Å (± 1.2) and 4.4 Å (± 1.3) away from any heavy atom in calbindin-D28K, respectively. For the LE scoring method there is a preference for small hydrophobic amino acids (Ala, Gly, and Val) in positions X1 and X2 and polar uncharged amino acids (Asn, Thr, and Ser) in positions X3 and X4 on the CX1X2SIKNX3X4C motif. However, the BE and Ki scoring methods (AUTODOCKderived) reveal a preference for positive electrostatic characteristic and aromatic hydrophobic amino acids. Generally the positive electrostatic amino acid is preferred in one or both of the first two variable positions (positions X1 and X2) while aromatic hydrophobic amino acids were favored in the last two variable positions (in positions X3 and X4) of the CX1X2SIKNX3X4C motif. These scoring methods resulted in 30, 27, and 33 out of the top 50 docked cyclic peptides with the disulfide bond playing no significant role in the interaction with calbindin-D28K, respectively. On average the disulfide bond was 4.5 Å (± 1.3), 4.0 Å (± 1.0), and 4.8 Å (± 1.4), respectively, away from any heavy atom in calbindin-D28K. In the BE scoring method, there is a preference for Lys, Arg, and/or His residues (in positions X1 and X2) as well as Phe and Trp residues (in positions X3 and X4). Finally, for the Ki scoring method there is a preference for charged amino acids such as Glu, Asp, His, Lys, or Arg (in positions X1 and X2) as well as Phe and Trp residues (in positions X3 and X4) (Table 2).
| BEa | LEb | Kic | LELPd | LIPEe |
|---|---|---|---|---|
| CCKSIKNRNC | CASSIKNNGC | CCESIKNIDC | CFFSIKNFFC | CFFSIKNLLC |
| CCWSIKNLRC | CGPSIKNTGC | CGVSIKNPKC | CFWSIKNFLC | CIFSIKNFWC |
| CHWSIKNWVC | CGSSIKNKPC | CHWSIKNYAC | CIWSIKNFFC | CIFSIKNLLC |
| CKFSIKNRAC | CGTSIKNTSC | CKFSIKNRQC | CWWSIKNWFC | CLLSIKNFWC |
| CWYSIKNHRC | CVASIKNPAC | CPDSIKNFPC | CWFSIKNWFC | CWWSIKNLFC |
Table 2: Scoring results of top 5 cyclic peptides (CX1X2SIKNX3X4C) bound to Calbindin-D28K
a Binding energy – determined through AUTODOCK
b Ligand efficiency – determined through AUTODOCK
c Estimated Inhibition Constant – determined through AUTODOCK
d Ligand efficiency dependent lipophilicity – (logP/LE)
e Liphophilic efficiency – (pKi-logP)
The binding surface of calbindin-D28K, identified through the druggability assay above, provides insight into the best potential cyclic peptide binders. The binding surface presented both an electrostatic and a hydrophobic cavity (Figure 2). Taking all of the above into consideration, the BE and Ki scoring methods provides the best characterization method for scoring this peptide library. These scoring methods identified peptides with charged and aromatic residues with a SIKN core as the best possibility for disruption of the calbindin-D28K:caspase-3 interaction. Disruption of the calbindin- D28K:caspase-3 interface would result in a release of caspase-3 and consequently lead to reactivation of the apoptotic cascade that is inactivated in cancer cells and cell death. HADDOCK (High Ambiguity Docking) was used to compare the conformational changes in the caspase-3 binding surface of calbindin-D28K when bound to caspase-3 versus one of the best-scoring cyclic peptides (CKFSIKNRQC -Table 2) [30].
HADDOCK allows for the movement of backbone and sidechain atoms to characterize conformational changes in the surface, whereas AUTODOCK generally does not. In particular, when calbindin-D28K is bound to CKFSIKNRQC, the hydrophobic caspase-3 binding surface on calbindin-D28K is significantly altered (Figure 2a-c). This suggests that binding of the cyclic peptide can allosterically alter the calbindin- D28K binding site, thereby interfering with caspase-3. To further verify our analysis, principal component analysis (PCA) (see Material and Methods) was used to analyze the: (i) calbindin-D28K:CKFSIKNRQC, (ii) calbindin-D28K:caspase-3 [8,10], and (iii) the 4-Ca2+ loaded NMR structure of calbindin-D28K [31] (Figure 2d-f). PCA assists in defining correlations in conformational rearrangements from a family of structures [32]. Shown in Figure 2, regions colored blue have a propensity to move together in a correlated fashion; regions colored red also have a propensity to move together in a correlated fashion but in an opposite (anti-correlated) fashion to those in the blue region. Manual visual inspection shows distinct differences in the motional propensities of the three systems. The PCA results suggest that upon binding a cyclic peptide, the conformational traits of calbindin-D28K are modified such that caspase-3 may no longer be recognized as a binding partner. Overall the molecular modeling data indicate that a cyclic peptide, such as CKFSIKNRQC, will bind calbindin-D28K in such a way that alters the conformational and dynamic characteristics of the protein where any interaction with caspase-3 would be impeded and thereby reactivating apoptosis.
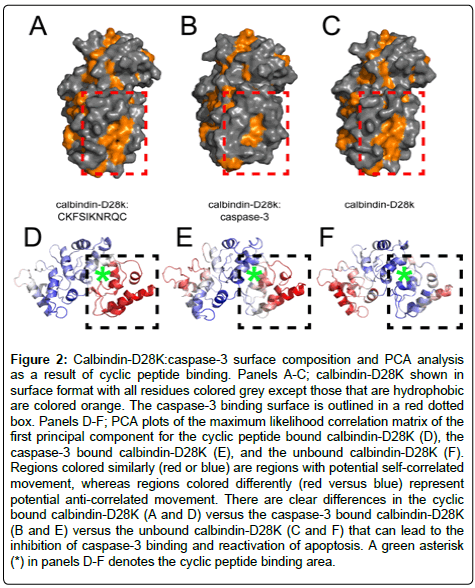
Figure 2: Calbindin-D28K:caspase-3 surface composition and PCA analysis as a result of cyclic peptide binding. Panels A-C; calbindin-D28K shown in surface format with all residues colored grey except those that are hydrophobic are colored orange. The caspase-3 binding surface is outlined in a red dotted box. Panels D-F; PCA plots of the maximum likelihood correlation matrix of the first principal component for the cyclic peptide bound calbindin-D28K (D), the caspase-3 bound calbindin-D28K (E), and the unbound calbindin-D28K (F). Regions colored similarly (red or blue) are regions with potential self-correlated movement, whereas regions colored differently (red versus blue) represent potential anti-correlated movement. There are clear differences in the cyclic bound calbindin-D28K (A and D) versus the caspase-3 bound calbindin-D28K (B and E) versus the unbound calbindin-D28K (C and F) that can lead to the inhibition of caspase-3 binding and reactivation of apoptosis. A green asterisk (*) in panels D-F denotes the cyclic peptide binding area.
Circumvention of apoptosis in cancer cells makes the apoptotic pathway ideal for targeted cancer therapy. Most cancer cells have become resistant to therapeutics that target the initiator caspases (caspases–2, –8, –9, and –10) proteins upstream in the apoptosis pathway. Therefore therapeutics directly targeting the effector proteins, such as caspase-3, should lead to reduced resistance and more effective therapies, since effector caspases ultimately are responsible for cellular suicide. Current therapeutics target the dimerization necessary for proper active site formation of all caspases. These unfortunately have limited results but the targeting of caspase-3 natural inhibitors such as calbindin-D28K offer new viable avenues of therapeutic intervention.
We would like to thank Dr. Erik A. Feldmann for critical reading of this manuscript. This research was funded by grant support from the V Foundation for Cancer Research and Swim Across America.