Journal of Physical Chemistry & Biophysics
Open Access
ISSN: 2161-0398
ISSN: 2161-0398
Research Article - (2016) Volume 6, Issue 5
The exact mechanism of binding of (-)-trans-Δ9-tetrahydrocannabinol (the main psychoactive component of marijuana) to the cannabinoid receptor, CB1, is unknown. Conformational information of the cannabinoids may give insight to this mechanism and the elicited effects of consumption. Herein, we report on the theoretical conformational study of Δ9-tetrahydrocannabinol (Δ9-THC) and cannabidiol (CBD), the psychoactive and a non-psychoactive compound found in marijuana, respectively, using electronic circular dichroism (ECD) and two-photon circular dichroism (TPCD). The compounds were optimized in vacuo and in the receptor site using DFT and B3LYP with the 6-311G** basis set and spectra were calculated using the same level of theory, but with the 6-311++G** basis set. First, we present and discuss the comparison of experimental and theoretical ECD spectra of (-)-trans-Δ9-THC and CBD in methanol solution in order to corroborate our theoretical approach. Second, we show, theoretically, the enhanced sensitivity of TPCD compared with ECD to conformational changes of cannabinoids upon docking, giving rise to a potential application in vivo. Finally, the comparative analysis of the theoretical TPCD spectra of Δ9-THC and CBD show distinct fingerprints in the far-UV region for two conformers of each molecule, which may help to understand the specific binding mechanisms of these species to the cannabinoid receptors and to describe the difference in psychological effects upon consumption. Our results show the complementarity of these two spectroscopic techniques and the potential of TPCD to determine the conformational changes of cannabinoids upon docking to the CB1 receptor.
<Keywords: Nonlinear circular dichroism; Cannabinoids; Protein shaving method; Ligand docking
For centuries, marijuana has been used for medical purposes due to its properties as a pain reliever, appetite stimulant and antiemetic [1]. More recently medical marijuana has been used in the treatment and prevention of several medical conditions including chemotherapyinduced nausea [2] and chronic pain and spasticity associated with multiple sclerosis [3]. With the onset of the legalization of medical and recreational marijuana in the United States, interest has increased in possible therapeutic uses of the drug. However, medical marijuana remains federally illegal in the U.S. since it has yet to go through the comprehensive studies required for Food and Drug Administration (FDA) approval.
Cannabinoids, a class of compounds unique to the cannabis sativa plant, are known to exhibit various physical and psychological effects upon consumption of marijuana. Remarkably, of the more than 80 cannabinoids present in cannabis sativa, (-)-trans-Δ9- tetrahydrocannabinol (THC) and cannabidiol (CBD) are considered the two most pharmacologically interesting. THC, the main psychoactive component of marijuana attributed with the euphoric effect [4,5], is also reportedly responsible for the stimulation of appetite [6,7] and reduction of pain [1] as well as increased anxiety, paranoia and impairment of memory, [8] among many other effects. While CBD is likewise credited with appetite stimulation and pain relief [9], it has been shown to have opposing psychological effects [10]. In fact, when administered together, CBD has been shown to reduce the psychotropic effects of THC [11-13], making it a potentially exceptionally useful therapeutic agent for psychological disorders.
The psychological effects of marijuana are specifically associated with the activation and blocking of cannabinoid 1 (CB1) receptors in the brain [14]. While THC is a known weak agonist, CBD acts as an antagonist to CB1, partially explaining the opposing effects. In the pursuit of therapeutic drugs without the euphoric effect, it would be helpful to understand why these different cannabinoids interact differently with CB1. Certainly, the molecular mechanism of cannabinoid docking to CB1 can provide clues to the varying effects of cannabinoids, but this mechanism is yet unknown. In an attempt to answer this question, we propose the following two possible scenarios:
• Scenario 1: The receptor site of CB1 has a slightly different conformation in each area of the brain which is responsible for controlling different functions. This may allow only certain ligands to bind in certain areas of the brain. In this case, it would be possible that CBD does not bind to CB1 in the area of the brain that produces the so-called “high”, but does bind in the area that controls, for example, appetite.
• Scenario 2: CB1 has the same conformation in all areas of the brain, which allows more than one type of ligand to bind. However, when the cannabinoids do bind, they induce different structural changes on the protein which sends a different signal to the cell, causing different effects.
Of course, we are aware that interest in this mechanism is not novel. In 2001, a generic mechanism for G-protein coupled receptors (GPCRs), a larger class of proteins that encompasses CB1, was theoretically predicted in which transmembrane helices of a GPCR rotate when the receptor is activated [15]. Other theoretical studies focused on changes in the receptor site when a ligand was bound. In 2004, a toggle-switch mechanism was predicted in which a tryptophan residue undergoes a χ1 rotamer change upon activation of CB1 [16]. However, the changes that cannabinoids undergo upon binding to the receptor remain unexplored theoretically and experimentally. Finding new ways to study these changes will contribute to the understanding of the proposed mechanisms.
Taking advantage of the chirality present in THC and CBD one could consider using electronic circular dichroism (ECD) for investigating the changes that cannabinoids undergo upon binding to the receptor. It should be highlighted that circular dichroism has been the standard technique of excellence for the study of optically active biomolecules during the last two decades [17]. While ECD has proven to be a reliable technique for studying large systems such as proteins, there are some limitations when working with small molecules, i.e., in the same spectral region where the signature of a small molecule is expected to appear, linear absorption of common solvents and buffers, and scattering at short wavelengths is observed. In order to overcome the limitations of ECD, Hernandez and co-workers recently proposed to use two-photon circular dichroism (TPCD) [18], the nonlinear counterpart of ECD. TPCD, first proposed theoretically in the ‘70’s by Tinoco [19] and Power [20], is defined as the difference between two-photon absorption (TPA) cross-sections using circularly polarized light of opposite handedness. This phenomenon was not able to be measured reliably until the development of the double L-scan technique in 2008 [21]. Since two-photon absorption occurs at lower energies than one-photon absorption (OPA), there is negligible linear absorption in the excitation region and scattering is minimized. Additionally, depth penetration and spatial resolution are increased [22] and photodamage to living cells is diminished [23]. Recently, TPCD has been used to investigate the structure of natural amino acids [24]. Vesga and co-workers showed theoretically that TPCD is more sensitive than ECD to the differences between α-helix, β-strand and random-coil conformations of a tryptophan residue. Based on this, we consider TPCD as a potentially effective method of detection of minor conformational changes of cannabinoids upon binding to the receptor site of CB1. In this article, we report the experimental and theoretical OPA and ECD of THC and CBD, as well as the theoretical TPA and TPCD spectra of the two most pharmacologically interesting cannabinoids, THC and CBD, calculated both in gas phase and when bound to the receptor site. Our results demonstrate the possibility of using TPCD for monitoring the conformational changes of cannabinoids docking to CB1 and determining the mechanism of activation of CB1 induced by cannabinoids.
THC and CBD were purchased in methanol from Lipomed AG. OPA measurements were performed on a single-beam spectrophotometer (Agilent 8453 Diode Array UV-Vis) in a 1 cm cell in 20 μM solutions. ECD measurements were carried out on 10-4 M and 10-3 M solutions of THC and CBD respectively on an AVIV 215 CD Spectropolarimeter with a path length of 0.1 cm and a scan speed of 1 nm/s. The alkyl chain attached to the aromatic ring of cannabinoids leads to a large degree of freedom of the molecule, so for each cannabinoid, two conformers (Figure 1) were considered. The structures were first optimized in vacuo using density functional theory (DFT) [25] and Becke’s three-parameter exchange, Lee, Yang and Parr correlation (B3LYP) functional [26-28] using the 6-311G** basis set [29,30] in Gaussian 09 [31]. Each of these conformers were then placed in the receptor site of CB1 to be re-optimized to the same level of theory. In order to optimize the molecules in the site, receptor site residues were isolated and connected using alkyl chains which were frozen during the optimization. OPA and ECD spectra were calculated using equations (1) and (2) for the first 100 excited states of each conformer optimized in vacuo and in site using time-dependent DFT (TD-DFT) [25] at the B3LYP/6-311++G** level of theory in Gaussian 09 [31]. The polarizable continuum model (PCM) [32] was used to account for solvent (meth anol) effects. OPA spectra are reported in molar absorptivity [32-34],

Figure 1: Chemical structure of (-)-trans-Δ9-tetrahydrocannabinol (THC) (top). Lowest energy conformations of THC (bottom). Optimizations were performed with DFT/B3LYP/6-311G** in gas phase using Gaussian 09.
 (1)
(1)
Here ω is the circular frequency of the incident light, ωof is the excitation circular frequency for a 0→f transition, fof is the oscillator strengths and Γ of g(ω ,ωof ,Γ ) is the Lorentzian lineshape function for the linear absorption case.
ECD spectra are reported in molar absorptivity difference from RofECD [33-35],
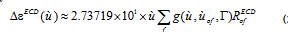 (2)
(2)
Where RofECD is the velocity rotatory strengths. Both have units of mol-1 cm-1 L as long as the elements in equations (1) and (2) are introduced in atomic units. TPA and TPCD spectra were calculated in gas phase for the first 50 excited states using TD-DFT at the B3LYP/6-311++G** level of theory in Dalton 2013 [36]. TPA spectra were calculated using [34,37]
 (3)
(3)
Where äofTPA (ùof) is the orientational two-photon probability for the degenerate case, Ã of g(2ω ,ωof ,Ã ) is a normalized Lorentzian line shape function where Γ is the line width and ω is the excitation frequency. TPCD spectra were calculated as described by Rizzo et al. using the following equation [19,34,38]
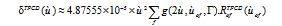 (4)
(4)
Where RofTPCD (ωof) is the TPCD rotatory strength which is obtained from:
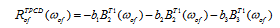 (5)
(5)
where b1, b2 and b3 are scalars that depend on the geometry of the experiment. In this case, it is assumed that two left or right circularly polarized photons are collinear and co-propagating so that b1=6, b2=2 and b3=-1. B1, B2 and B3 are molecular parameters all of which depend on the electric transition dipole moment. B1 and B3 also depend on the magnetic transition dipole moment and B2 depends on the electric transition quadrupole moment. TPA and TPCD spectra obtained from equations (3) and (4) are in units of Göppert-Mayer (GM) (10-50 cm4·s·molecule-1·photon-1) as long as the equation elements are introduced in atomic units.
Validation of theoretical approach
In order to initially validate our theoretical approach, the experimental OPA and ECD spectra of THC and CBD in methanol and their corresponding theoretical spectra calculated in methanol using B3LYP/6-311++G(d,p) are compared in Figure 2. Theoretical spectra were slightly blue-shifted, i.e., -8 nm (THC) and -6 nm (CBD), to obtain a better theoretical-experimental overlap (this is common practice in theoretical-experimental work) [18,39-42]. It should be noted that while the ECD of cannabinoids has previously been reported [43], basic conditions were employed during the measurements, significantly altering the spectra. For each cannabinoid we calculated OPA and ECD for two conformers (red and blue in the figure for conformer 1 and 2, respectively) and averaged the resulting spectra (violet) for comparison with experimental results. We realize that eliminating the alkyl chain might yield spectra that more accurately represent a population of conformers, but the initial position of the chain proved to be especially important during binding. Nevertheless, in both cases the average predicted spectra (violet) match very well with our experimental results. We only observed deviations in the ECD spectrum of CBD on the red side of the spectrum where the bands are more negative than predicted (indicated with orange arrows in the figure). This minor deviation can be attributed to the larger degree of freedom associated with CBD due to the absence of the central ring that maintains the relative rigidity of THC.

Figure 2: Comparative plots of experimental (black dots) and calculated (solid lines) OPA (left) and ECD (right) spectra of cannabinoids in solution. OPA and ECD response for the lowest 100 excited states of optimized structures were calculated with TD-DFT/B3LYP/6-311++G** in methanol using Gaussian 09. Spectra calculated for each conformer (red dotted line for conformer 1 and blue dashed line for conformer 2) were averaged (violet solid line) to match experimental, are only shown within the experimentally measureable range (190-260 nm) and have been shifted: THC (-8 nm) and CBD (-6 nm). Γ=0.35 eV (FWHM) was used for all calculated spectra. Excited states that significantly contribute to the spectral features observed in theoretical and experimental spectra are identified in the SI.
Linear and nonlinear characterization
Having established that the theoretical approach is valid, we computed the OPA, ECD, TPA and TPCD spectra for both conformers of THC and CBD in methanol when optimized in vacuo and in site using B3LYP/6311++G(d,p) and Γ=0.35 eV (FWHM) for OPA and ECD and Γ=0.2 eV (FWHM) for TPA and TPCD. Comparison of theoretical OPA and TPA spectra for each molecule optimized in vacuo and in site revealed insignificant differences in all cases (These spectra can be found in the SI). First we compare the calculated ECD spectra of each conformer of THC. These spectra are presented on the left side of Figure 3. Differences in the ECD spectra in a region between 180-190 nm suggest that both THC conformers in vacuo would be distinguishable from their counterparts in site using this technique. In reality, though, this region start becoming experimentally unreliable due to scattering and linear absorption of the solvent. The calculated TPCD spectra comparing each conformer of THC in vacuo and in site are presented on the right side of Figure 3. Regarding the first conformer, there is a fingerprint around 400 nm that would allow the conformer in site to be distinguished from the one in vacuo. Additionally, the signals for THC1 optimized in vacuo and in site are opposite in sign around 420 nm which would make them clearly distinguishable experimentally. The most dominant transitions in the 380-410 nm region, where we observe the appearance of the fingerprint after docking, change from 31-33 in vacuo to 29 and 41 in site. We attribute these bands to high energy σ→σ* transitions that are affected by the slight movement (less than 3° change in the angle between the scaffold and the chain) of the side alkyl chain in order to avoid steric effects from the rotating tryptophan residue. For THC2, the differences in the TPCD spectra are less significant and occur in a different region than THC1. The signals for THC2 in vacuo and in site are opposite in sign between 460 and 490 nm. The dominant transitions in this region change from 4 and 7 to 6 and 7 and also change from positive to negative. Since the change is attributed to the tricyclic scaffold rotation to a higher degree for this conformer, the position of the bands allows us to assign these relatively lower energy transitions to π→π*. In addition to comparing the conformations in vacuo and in site, we performed the comparison of both conformers of THC in the receptor site to determine whether they could be differentiated experimentally. Examination of the TPCD spectra reveals fingerprints for conformer 1 in site at 400 and 440 nm and the signals are opposite in sign around 400 and 480 nm. This would allow for the experimental determination of the conformation of THC in site for one or a mixture of both conformers which would not be possible using only ECD. Comparison of the ECD spectra of both conformers of the reportedly non-psychoactive component of marijuana, CBD, reveal seemingly significant differences (Figure 4). However, if either spectrum were to undergo a spectral shift, the conformers would be experimentally indistinguishable. Furthermore, comparison of the conformers of each molecule in site confirms that while identification of the cannabinoid may be possible, conformation would be ambiguous. The main difference between the TPCD spectra of CBD and those of THC is the increased amplitude of the signal which is between 3 and 4 times greater for CBD. This may be attributed to the increased degree of freedom associated with this cannabinoid. The TPCD spectra for CBD1 have similar shapes in vacuo and in site. However, the signals are opposite in sign (around 430 nm) due to the negative contributions of transitions 18, 22 and 26 in vacuo and the positive contributions of transitions 20, 25 and 26 in site. These drastic changes, coupled with the fingerprints for conformer 1 around 400 and 460 nm and for conformer 2 around 430 nm would allow for experimental identification of the conformers. Similar to what was observed for THC, the intensification of signal around 400 nm and appearance of a positive band around 430 nm can be attributed to σ→σ* transitions in the alkyl chain and the appearance of bands between 460 and 540 nm indicate π→π* transitions associated with the rotation of the cyclic scaffold. On the other hand, the TPCD spectra of CBD2 are similar in vacuo and in site and the conformers would be challenging to distinguish. However, the disappearance of the signal around 530nm due to the change in dominant contributions from 3 to 1 would indicate the presence of this conformer in site and again could be attributed to low energy π→π* transitions. There is also a slight red shift (about 10 nm) in this peak due to the transitions 34 and 37 in vacuo changing to 29 and 33 in site. This common fingerprint would allow the conformers to be distinguished from THC, but since the signal for both conformers in site is positive across the spectrum, the conformation of CBD would be challenging to identify in site.

Figure 3: Comparative plots of ECD (left) and TPCD (right) spectra of (-)-trans-Δ9-THC conformers. Dotted lines correspond to structures optimized in gas phase, solid lines correspond to structures optimized in the receptor site. ECD response for the first 100 excited states of optimized structures were modeled with TD-DFT/ B3LYP/6-311++G** in methanol using Gaussian 09. TPCD response for the first 50 excited states were modeled with TD-DFT/B3LYP/6-311++G** in gas phase using Dalton 2013. Dominant transitions are labeled, colored arrows indicate regions of the corresponding spectra that could allow for clear identification of which conformer is present. Shaded region of ECD spectra is experimentally unreliable.

Figure 4: Comparative plots of ECD (left) and TPCD (right) spectra of CBD conformers. Dotted lines correspond to the structure optimized in gas phase, solid lines correspond to the structure optimized in the receptor site. ECD response for the first 100 excited states were modeled with TD-DFT/B3LYP/6-311++G** in methanol using Gaussian 09. TPCD response for the first 50 excited states were modeled with TD-DFT/B3LYP/6-311++G** in gas phase using Dalton 2013. Dominant transitions are labeled, colored arrows indicate regions of the corresponding spectra that could allow for clear identification of which conformer is present. Shaded region of ECD spectra is experimentally unreliable.
TPCD was confirmed to be more sensitive than ECD to the conformational changes of cannabinoids associated with binding to CB1. TPCD may act as a method for identifying not only which cannabinoid is bound, but what conformation it is in. Our results have shown evidence to support the scenarios in which the varying effects of cannabinoids may be attributed to the conformational changes experienced both by the receptor and the ligand during binding.
This work was partially supported by the United States National Science Foundation through Grant Number DBI-1422826. The computing time provided by STOKES ARCC is gratefully acknowledged. We acknowledge Dr. Suren Tatulian at the University of Central Florida for allowing us to use his CD Spectrometer. Dr. Mohanraja Kumar at the University of South Florida is also acknowledged for allowing us the use of the CD Spectrometer at that facility.