Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Review Article - (2016) Volume 4, Issue 1
Fucoxanthin is a carotenoid present in brown micro- and macro algae, that induces apoptosis or autophagy in cancer cells grown in vitro. In vivo studies confirmed its interest as a natural anticancer compound, as it exerts antitumoral, antimetastatic and antiangiogenic activities in animal models. Studies focused on the pharmacology of fucoxanthin in cancer cells and tumors have revealed that it affects a wide panel of cellular, molecular and tissular processes, suggesting that its biological activity may be related in part to a nonspecific integration in cell membranes, and possible interaction with lipid rafts. Thus, preliminary data confirming this interaction of fucoxanthin with lipid rafts were obtained in mast cells and hepatoma WIF-B9 cells. We here discuss this hypothesis, in view of the critical function of lipid rafts in cancer cell survival, invasivity and communication with the tumoral microenvironment.
<Keywords: Algae; Apoptosis; Cancer; Carotenoid; Fucoxanthin; Lipid raft; Phytoplankton; Pigment
Plant and algae carotenoids have a great potential as natural anticancer compounds as they exert cytotoxic, antitumoral, antimetastatic, and antiangiogenic activities [1,2]. Particularly, fucoxanthin (Figure 1), a marine carotenoid purified from brown micro- and macroalgae, attracts extensive interest owing to its antiproliferative, pro-apoptotic and pro-autophagic activities, demonstrated in a large panel of cancer cells [3-14].
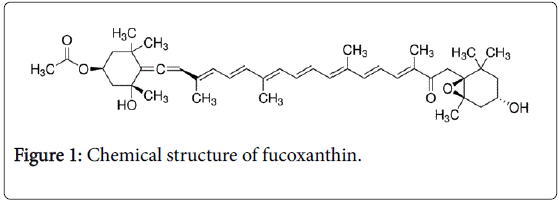
Figure 1: Chemical structure of fucoxanthin.
Its antitumoral activity was validated in animal models, in which it limits tumor growth, angiogenesis and invasivity, suggesting a possible use for clinical oncology [1,15]. Studies focused on the pharmacology of fucoxanthin in cancer cells and tumors have revealed that it induces cell cycle arrest in G0/G1 phases [5], cyclins and CDK downregulation, CDK inhibitors up-regulation [16], as well as the common hallmarks of apoptosis (disturbance of the mitochondrial transmembrane potential, modulation of the expression of the Bcl-2 family proteins, activation of caspases/PARP/CAD, DNA fragmentation and phosphatidylserine exposure) (for review see [3]).
Fucoxanthin was also reported to inhibit mammalian topoisomerases [2,3], disturb the MAPK and NF-kB pathways [17], enhance Gap junctional intercellular communication by up-regulating the expression of connexins [12], and reverse multi-drug resistance by interfering with drugs efflux systems [3,13]. It also inhibits matrix metalloproteases involved in tumor tissular invasivity [8,18], and induces autophagy in some cancer cell models [19]. This multimodal activity has led us to hypothesize that the cytotoxic activity of fucoxanthin may be related, at least in part, to a nonspecific integration into plasma and/or intracellular membranes, triggering a wide variety of cellular responses eventually leading to cancer cells apoptosis or autophagy. In support of this hypothesis, fucoxanthin contains two polar heads, separated by a polyene chain, that confer to the molecule a bolaphile behaviour and the appropriate size for its integration into plasma and intracellular membranes. Accordingly, it is assumed that the first evolutionary function of carotenoids was to rigidify the cell membranes of archaebacteria [20]. In algae, fucoxanthin locates in thylakoid membranes where it is tightly bound to chlorophyll molecules and apoproteins to form functional lightharvesting complexes [21]. In the same way, dietary carotenoids such as lutein and zeaxanthin concentrate in the rod outer segment membranes in human retina [22] where they play a key function in eye photoprotection against the deleterious wavelengths of blue light [23]. The first indication that carotenoids may interfere with lipid rafts in the plasma membrane has come from the observation that fucoxanthin, astaxanthin, zeaxanthin and b-carotene inhibit the antigen-induced degranulation of basophiles and mast cells [24]. In these models, it is assumed that fucoxanthin integration into lipid rafts blocks the translocation and aggregation of high affinity IgE receptors (FcεRI) and subsequently inhibits the signaling cascade triggering degranulation [24]. To address this hypothesis in other cell types, we recently investigated the interaction of fucoxanthin with subcellular fractions obtained by ultracentrifugation of Wif-B9 hepatoma cells using the flotation method [25]. The WIF-B9 hepatic cell line, obtained by fusion of Fao rat hepatoma cells and WI 38 human fibroblasts, are highly differentiated and polarized cells, developing morphologic features close to primary rat and human hepatocytes. They constitute a relevant model for hepatotoxicity assays, xenobiotic metabolism studies and evaluation of xenobiotic interaction with lipid rafts [25]. In these preliminary investigations using cells treated with 10 μM fucoxanthin for 24 h, we observed a tendency of fucoxanthin to preferentially incorporate into subcellular domains containing lipid rafts (66.5 %) when compared to those devoided of lipid rafts, according to absorbance increases at 446 nm per mg proteins (unpublished data). This observation is in line with the data obtained in mast cells, and suggests that fucoxanthin probably interacts with lipid rafts in several cell types. If confirmed, the demonstration of this interaction opens new perspectives to discuss the biological activity of fucoxanthin and understand why it exerts cytoprotective effects in normal cells, while it induces apoptosis or autophagy in cancer cells.
Fucoxanthin is usually considered as a potent antioxidant because of the presence of an allenic bond and hydroxyle function in its structure. In normal cells, in a context of low oxidative stress, it is conceivable that fucoxanthin integrates cell membranes in domains containing or not lipid rafts, and scavenges ROS that could trigger lipid oxidation and inflammation. Some studies suggest that the antioxidant activity of fucoxanthin may also be related to its ability to undergo ring-opening reactions on its 5,6 monoepoxide function and exert a pro-oxidant activity on surrounding molecules [26], eventually leading to an activation of the transcription factor Nrf2 (nuclear factor (erythoidderived 2)- like2, stimulating the expression of antioxidant enzymes, such as glutathione peroxidase, and NAD(P)H quinone oxydoreductase 1 [26,27]. In cancer cells, because of an increased ROS production and high oxidative stress, integration of fucoxanthin in cell membranes may induce the generation of high levels of ROS, that exceed the capacity of antioxidant enzymes and eventually trigger apoptosis or autophagy. In support of this hypothesis, it was shown that fucoxanthin exerts a pro-apoptotic activity in human leukemia cells through a ROS-mediated BCL- xL pathway [28] and that this proapoptotic activity is inhibited by the antioxidant N-acetylcysteine. In addition to its pro- and antioxidant activity, it is reasonable to speculate that fucoxanthin interaction with lipid rafts may participate to its cytotoxicityin cancer cells. Lipid rafts are cholesterol and sphingolipid-enriched microdomains found in the plasma and Golgi membranes, characterized by their resistance to solubilization in nonionic detergents at low temperature and ability to float and concentrate in low-density fractions of a sucrose gradient after ultracentrifugation [29,30]. They are mobile and play a major function in protein trafficking and integration of signal transduction complexes, as they can rapidly include or exclude proteins and mediate proteinprotein interactions. Lipid rafts are thus involved in key cellular functions, such as the control of cytoskeletal dynamics for cell adhesion and migration, or regulation of ion channels and signaling complexes trafficking to the plasma membrane. As such, they are also critically involved in adhesion, motility, invasivity, survival and proliferation of cancer cells, and represent an innovative target for the development of new anticancer drugs [31]. It has been shown that alteration of the lipid rafts composition (e.g. decrease in the cholesterol content) or integration of exogeneous lipids into lipid rafts (e.g. alkylphospholipid analogues such as edelfosine) could trigger cancer cells apoptosis [32-35]. It is also demonstrated that cell death can be initiated by activation of death domain receptors, located in lipid rafts [33]. Previous studies have documented that some cancer cells contain an increased level of cholesterol-rich lipid rafts [36] and that a cholesterol-depleting agent such as methyl-b-cyclodextrin triggered BcL-XL down regulation, caspase-3 activation and Akt inactivation [36]. These changes, that correspond to classical observations associated with apoptosis, have also been reported in cancer cells treated with fucoxanthin [3]. In this view, the selective cytotoxicity of fucoxanthin towards cancer cells may be explained in part by the elevated level of lipid rafts in cancer cells and increased ability of fucoxanthin to integrate their plasma membranes. Lipid rafts have been involved in the triggering of death receptor-mediated apoptosis through the recruitment and activation of Fas/CD95 death receptor [38]. Activation of Fas leads to the subsequent formation of the deathinducing signaling complex (DISC),that triggers apoptosis. Further studies should thus be undergone to determine if fucoxanthin cytotoxicity in cancer cells may be related to its ability to trigger Fas activation in lipid rafts and formation of the DISC. It is also well established that cancer cell migration and metastatic behaviour is controlled in part by ion channels associated in functional membrane complexes located in lipid rafts. As an example, the interaction between the SK3 potassium channel and Orai-1 calcium channel was documented as critical for the metastastic potential of breast cancer cells [38]. Accordingly, integration of apolar drugs such as ohmline into lipid rafts has proved efficacy to disrupt the interaction of SK3 and Orai-1 channels in lipid rafts and inhibit the metastatic potential of cancer cells [38]. The demonstration that fucoxanthin can integrate lipid rafts opens the way for further studies to understand if its antimetastatic activity may be related to a disturbance in the interaction and functionality of such ion channels complexes located in lipid rafts. Additionally, dysregulation of ions channels expression is often observed in cancer cells [39]. These alterations promote cell proliferation, tissular invasivity and insensitivity to growth inhibitory signals. Fucoxanthin may counteract such dysregulations by disturbing ions channels integration and association in functional complexes in the plasma membrane. Ions channels and connexins also play a pivotal role to control the molecular dialogue between cancer cells and the tumor microenvironment. Fucoxanthin was reported to stimulate the expression of connexins 32 and 43 in cancer cells, enhance Gap junction intracellular communication and increase the concentration of intracellular Ca2+ [12]. Stimulation of the gap junctional communication facilitates the distribution of anticancer drugs within neighbouring tumor cells, and may explain in part why fucoxanthin is able to sensitize tumor cells to some anticancer drugs. Increase in the intracellular calcium concentration induced by fucoxanthin may also explain in part how fucoxanthin triggers cell cycle arrest, caspase activation and cytoskeleton reorganization. Changes in the whole cell shape and cell movement involve rapid and transient reorganization of membrane micro domains that are mostly mediated by the cytoskeleton dynamics in response to activation of membrane signaling pathways and local calcium concentration changes. Thus, integration of fucoxanthin in lipid rafts may induce major disturbances in these regulatory pathways and dysfunctions of the cytoskeleton. Fucoxanthin was also reported to decrease the expression of CD44 [8], a cell surface glycoprotein involved in cell-cell interactions, cell adhesion and migration, and interaction with matrix metalloproteases (MMPs), that plays a critical function in invasion of the tissular environment of the tumor by cancer cells. Lipid rafts play a crucial role in the localization and functionality of CD44 as proved using cholesterol-depleting agents, that stimulate CD44 shedding and supress cancer cell migration [40]. Consequently, inhibition of MMPs by fucoxanthin and its physiological metabolite fucoxanthinol may be related to their integration into lipid rafts and disturbance of CD44 expression. Fucoxanthin was also reported to inhibit angiogenesis in various tumor models or endothelial cells (e.g. HUVEC) by supressing the mRNA expression of FGF-2 and its receptor FGFR-1, as well as their trans-activation factor, EGR-1 [40]. It also induces a decrease in the phosphorylation of FGF-2-mediated intracellular signaling proteins such as ERK1/2 and Akt, eventually leading to a repression of the migration of endothelial cells and inhibition of their differentiation into tube-like structures in Matrigel [40]. It it possible to hypothesize that incorporation of fucoxanthin may interfere with the function of growth factors receptors, that are located in lipid rafts. Finally, fucoxanthin may also disrupt protein palmitoylation or myristoylation, and alter their submembrane localization. The possible incorporation of fucoxanthin into organelle membranes, particularly mitochondrial membranes, and interaction with proteins of the Bcl-2 family, may also not be excluded. To conclude, the rigorous demonstration that fucoxanthin interacts with lipid rafts may provide a new clue to understand the cellular and molecular mechanisms involved in its cytotoxicity in cancer cells. Continued exploration of fucoxanthin cytotoxicity in cancer cells should confirm if its interaction with lipid rafts is a key event triggering apoptosis or autophagy (Table 1).
| Topic | Reference numbers |
|---|---|
| Structure and biological activity of fucoxanthin and carotenoids in cancer cells | [1-18] |
| Biological functions of carotenoids in cell membranes | [19-22] |
| Interaction of carotenoids with lipid rafts | [23] |
| Cell model to study the integration of xenobiotics into lipid rafts | [24] |
| Antioxidant and pro-oxidant activity of fucoxanthin in animal cells and tissues | [25-27] |
| Structure and function of lipid rafts in cell membranes | [28,29] |
| Targeting lipid rafts in cancer cells | [30-36] |
| Ion channels, lipid rafts and cancer cells | [37,38] |
| Lipid rafts, cytoskeleton, cancer cells invasivity and tumor angiogenesis | [30,40] |
Table 1: References cited in the text.
This research was financially supported by the French cancer league (Comité 17 de la Ligue Nationale contre le Cancer). We also thank the “Cancéropôle Grand Ouest, axe Valorisation des produits de la mer en cancérologie and the Brazilian Society of Pharmacognosy for scientific support.
The authors declare no conflict of interest.