Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2013) Volume 1, Issue 2
Antarctic benthic communities below anchor ice and ice scour are subjected to intense biotic interactions, and this has propitiated the development of chemicals to avoid predation, competition, and/or fouling avoidance. In particular, Sterechinus neumayeri, a common eurybathic sympatric sea urchin species, can have a negative effect on early recruitment of benthic organisms through grazing. An adaptive response of these co-existing benthic animals may consist in displaying cytotoxic activities, poorly investigated so far in Antarctic ecosystems, in order to prevent the settlement of embryos and larvae of this sea urchin on them or near and, consequently, to decrease its grazing pressure. Cytotoxic activities of Antarctic benthic organisms, mainly from the deep waters of the poorly surveyed area of the eastern Weddell Sea, were assessed against embryos and sperm of the sea urchin S. neumayeri. A new methodology based in a standardized protocol was adopted for this species. Bioassays were performed at Deception Island (South Shetland Islands) during the Austral Summers of 2008-2009 and 2009-2010, using ether extracts at different concentrations from different benthic organisms collected in previous Antarctic expeditions. A high percentage (80%) of the species tested (29) from eight different phyla (Porifera, Echinodermata, Cnidaria, Chordata, Bryozoa, Annelida, Nemertea and Hemichordata) were toxic in one or both experiments. In the bioassay of cytotoxicity against embryos, seven of the 14 species (50%) tested did not reach the blastula stage at the highest extract concentration. In the case of the cytotoxicity against sperm, a total of 20 species were tested and all the extracts except one were toxic to the sperm at the maximum concentration. In addition, 6 species were dissected into parts to investigate the possible allocation of defensive compounds. Our results highlight the important role of cytotoxic activity of some Antarctic benthic organisms as a possible chemical defense and/or mechanism of competition for space or food against this common sea urchin.
<Keywords: Chemical defenses; Chemical ecology; Early life stages; Optimal defense theory; South Shetland Islands; Weddell Sea
Recent studies on Antarctic biodiversity have described a surprisingly rich, varied and eurybathic benthic fauna [1-5]. This richness and diversity may be related to the biogeographic isolation and old age of this continent [6,7]. In general, the predominant groups inhabiting these Antarctic benthic communities are sessile suspension feeders, e.g. ascidians, sponges and bryozoans [8,9], conferring surprising tridimensional structures [10]. Below anchor ice and ice scour (8,11]. All this makes it likely that many of the organisms inhabiting these waters have evolved chemical mechanisms to avoid predation [12-15], competition for space or food [16,17] and fouling [18-22], thus creating a complex chemical ecological network [23-25].
The bulk of work conducted so far, demonstrating the relevance of this kind of chemical defenses, has been conducted in temperate and tropical waters [26-31]. To date, however, little work has been carried out in areas such as Antarctica when compared to other geographic areas [23,24,32]. Moreover, there is very limited information about chemical ecology of deep-water species from the eastern Weddell Sea [23,24,33]. In particular, new natural products have been isolated from Antarctic organisms, although most of their prospective ecological roles are still unknown [23]. Furthermore, few studies on sponges, ascidians and molluscs have described the location of active compounds within Antarctic organisms [20,21,34,35]. In this sense, according to the Optimal Defense Theory (ODT) based on the body allocation of chemical defenses, the active compounds are postulated to be located in the most vulnerable and/or valuable parts or the most exposed to predation, in order to improve their efficiency in protection and to invest the minor possible cost in their production [36].
As part of the Antarctic benthic vagile fauna, echinoderms play a remarkable role in Antarctic benthic ecosystems and control the populations of many benthic organisms [37]. Among the echinoderms, Sterechinus neumayeri Meissner, is considered to be one of the most abundant and common species found in Antarctica [38]. Sterechinus (Order Echinoidea, Family Echinidae) is the only genus of the Antarctic echinoids, comprising 5 species, S. neumayeri being the most abundant [38]. This sea urchin has a circumpolar distribution and a wide bathymetric record, ranging from few meters to 810 m depth [39,40]. This species, as most marine invertebrates, produces larvae, which may travel large distances before settling in a suitable substratum [41]. The larval settlement is considered very important in mediating the population structures of this sea urchin and other invertebrates and therefore, the chemical defenses in eggs, embryos and larvae are very important to reduce their mortality [42-44].
Sterechinus neumayeri has been used as a model organism in previous studies and its biology is well known [41,45-48], but only a few studies have investigated these indirect chemically mediated interactions against the development of embryos of this species [43,49]. In fact, S. neumayeri can be the responsible, through grazing, for significant mortality of settling larvae and juveniles of benthic organisms [18,37,50]. Thus, the intense predation pressure exerted by this sea urchin could have lead to develop cytotoxic compounds in their preys, that might act at small scale against embryos and larvae of this sea urchin to prevent the settlement on them or near and, consequently, the grazing pressure [43,49,51,52].
Here, we use the sea urchin sperm and eggs of S. neumayeri in two bioassays to determine: 1) whether extracts from eight different phyla (Porifera, Echinodermata, Cnidaria, Chordata, Bryozoa, Annelida, Nemertea and Hemichordata) from the eastern Weddell Sea, Livingston Island and Deception Island, with a wide bathymetric range, possess natural products that may serve as embryo/sperm Cytotoxics, 2) whether there is a location of active compounds in the body of selected species, using the ODT approach and 3) whether interspecific/ intraspecific variability exists in some species.
Samples collection and identification
Benthic organisms were collected from the eastern Weddell Sea and two islands in the South Shetland archipelago (Antarctica). Samples from the eastern Weddell Sea were obtained from a wide bathymetric range (9 m to 597 m) and collected during the ANTXV/3 (January- March 1998) and ANT XXI/2 (November 2003-January 2004) cruises on board the R/V Polarstern, from the Alfred Wegener Institute for Polar and Marine Research (Bremenhaven, Germany).
Samples from the South Shetland Islands were collected on board the R/V BIO-Hespérides in Livingston Island (December 2005 - January 2006) using Agassiz trawl, bottom trawl, epibenthic sledge and benthic traps (Table1) and by scuba diving at Deception Island (Austral Summers of 2008-2009 and 2009-2010). Organisms collected on board were sorted on deck and immediately frozen at -20°C, and later transported to the laboratory. A voucher specimen or a portion of each sample was fixed in 10% formalin or 70% ethanol for taxonomical determination and it is stored at the Dept. of Animal Biology (Invertebrates), University of Barcelona.
| Species | Location | Latitude (S) | Longitude (W) | Gear (a) | Depth (m) | Activity (b) |
|---|---|---|---|---|---|---|
| Annelida, Polychaeta | ||||||
| Nephtyidae sp. | Weddell Sea | 71° 06.30' | 11° 32.04' | AT | 175 | ST |
| Polyeunoa sp.(1) | Weddell Sea | 70º 52.6' | 10º28.6' | AT | 248 | ST |
| Polyeunoa sp.(2) | Weddell Sea | 70º 52.2 | 10º29.0 | AT | 246 | ST |
| Polyeunoa sp.(3) | Weddell Sea | 71º 08.6 | 12º286.6 | AT | 433 | ST/ ET |
| Travisia sp. | Weddell Sea | 61º 33.9 | 58º12.2 | AT | 417 | ST |
| Bryozoa, Gymnolaemata | ||||||
| Alcyonidium flabelliforme Kirkpatrick, 1902 | Weddell Sea | 71° 07.15' | 11° 26.23' | AT | 228 | ST/ ET |
| Cellaria diversa Livingstone, 1928 | Weddell Sea | 71° 06.30' | 11° 32.04' | AT | 175 | ST |
| Klugeflustra antarctica Hastings, 1943 | Livingston Is. | 62º46.43' | 60º27.02' | AT | 42 | ET |
| Cnidaria, Anthozoa | ||||||
| Alcyonium roseum van Ofwegen, Haaäussermann & Foörsterra, 2007 |
Weddell Sea | 71° 04.30' | 11° 33.92' | BT | 308 | ST |
| Alcyonium haddoni Wright & Studer, 1889 | Deception Is. | 62° 59.55’ | 60° 33.68’ | D | 9 | ET |
| Primnoidae sp. | - | - | - | - | - | ST |
| Thouarella laxa Versluys, 1906 | Weddell Sea | 72° 51.43' | 19° 38.62' | BT | 597 | ST |
| Hemichordata, Pterobranchia | ||||||
| Cephalodiscus sp. (1) | Weddell Sea | 70° 50.08' | 10° 34.76' | AT | 274 | ST |
| Cephalodiscus sp.(2) | Weddell Sea | 70° 57.11' | 10° 33.32' | BT | 337 | ST |
| Cephalodiscus sp. (3) | Weddell Sea | 70° 52.75' | 10° 51.24' | BT | 294 | ST |
| Nemertea, Anopla | ||||||
| Parborlasia corrugatus McIntosh, 1876 | Livingston Is. | 62º43.449 | 60º27.58 | TR | 52 | ST/ET |
| Porifera, Demospongiae | ||||||
| Iophon sp. | Livingston Is. | 62º46.43' | 60º27.02' AT | 42 ST | ||
| Iophon sp. + Ophiurolepis (1) | Weddell Sea | 71° 06.44' | 11° 27.76' | AT | 277 | ST |
| Iophon sp. + Ophiurolepis (2) | Weddell Sea | 71° 06.30' | 11° 32.04' | AT | 175 | ST |
| Isodictya verrucosa Topsent, 1913 | Weddell Sea | 71° 06.44' | 11° 27.76' | AT | 277 | ET |
| Mycale (Oxymycale) acerata Kirkpatrick, 1907 | Weddell Sea | 71° 04.30' | 11° 33.92' | BT | 308 | ST |
| Porifera, Hexactinellida | ||||||
| Anoxycalyx (Scolymastra) joubini Topsent, 1916(1) | Weddell Sea | 70° 52.16' | 10° 43.69' | BT | 290 | ET |
| Anoxycalyx (Scolymastra) joubini Topsent, 1916(2) | Weddell Sea | 70° 56.43' | 10° 31.47' | AT | 279 | ET |
| Anoxycalyx (Scolymastra) joubini Topsent, 1916(3) | Weddell Sea | 70° 57.11' | 10° 33.52' | BT | 337 | ET |
| Rossella fibulata Schulze & Kirkpatrick, 1910 | Weddell Sea | 70° 57.11' | 10° 33.52' | BT | 337 | ET |
| Rossella nuda Topsent, 1901 | Weddell Sea | 71° 04.30' | 11° 33.92' | BT | 308 | ET |
| Rossella sp. Carter, 1872 | Weddell Sea | 70° 55.92' | 10° 32.37' | AT | 288 | ET |
| Rossella vanhoffeni Schulze & Kirkpatrick, 1910 | Weddell Sea | 72° 28.91' | 17° 51.28' | ES | 882 | ET |
| Porifera sp. | Weddell Sea | 71° 06.44' | 11° 27.76' | AT | 277 | ST |
| Porifera sp. | Weddell Sea | 71° 06.44' | 11° 27.76' | AT | 277 | ET |
| Chordata, Ascidiacea | ||||||
| Cnemidocarpa verrucosa Lesson, 1830 (1) | Weddell Sea | 70° 56.67' | 10° 32.05' | BT | 302 | ET |
| Cnemidocarpa verrucosa Lesson, 1830(2) | Weddell Sea | 70° 57.00' | 10° 33.02' | BT | 332 | ET |
Table 1: Collection data from the benthic organisms used for this study. Sampling gears: a) AT, Agassiz Trawl; BT, BottomTrawl; ES: Epibenthic sledge; TR, Benthic traps; SD, Scuba diving. Activity b) ST: sperm toxicity, ET: embryo toxicity.
Specimens of the sea urchin Sterechinus neumayeri to be used during the cytotoxic experiments were collected by scuba diving at Whalers Bay (Deception Island: 62° 59.369′ S, 60° 33.424′ W) from 10- 15 m depth. The size of sea urchins used in the experiments ranged between 40-60 mm in diameter. After the experiments, the sea urchins were brought back alive to the sea at Whalers Bay.
Chemical extractions
Chemical extraction of frozen organisms was done in the laboratories of the Faculty of Biology, University of Barcelona. When possible, animals were carefully dissected into different sections, in order to locate the compounds within the body of the organisms. The sections were separated according to the taxonomic group (polyps and axis, in gorgonians; disc, in ophiuroids; external/internal parts, and apical/basal regions, in sponges; external tunic or internal part and gill slits, in tunicates).
These body sections were then extracted with acetone, and sequentially partitioned into ether and butanol extracts. All steps were repeated thrice, except for the butanol which was done once. Organic solvents were then evaporated under reduced pressure, resulting in dry ether and dry butanol extracts, and an aqueous residue. An aliquot of all the ether extracts were used for bioassays at in three concentrations (1 mg ml-1, 0.5 mg ml-1, and 0.05 mg ml-1) for the experiments. We selected the ether extracts for testing the experiments because the nature of most marine natural compounds with known chemical activity is lipophilic [26]. Butanol extracts and water residues were kept for further analysis.
Common procedure for the bioassays
In the Antarctic marine benthic environment, S. neumayeri is one of the most abundant and common grazer species with a varied diet and its biology is well known [41,45,52], being thus selected as model organism for our bioassays. The experimental work took place at the ‘Gabriel de Castilla’ Spanish Antarctic Base at Deception Island during two Antarctic cruises within the frame of the ACTIQUIM project (Austral Summer 2008-2009 and 2009-2010). Ether extracts previously prepared were transported frozen to the Antarctic Base. After collection, sea urchins were acclimatized during three days in tanks with fresh seawater. After acclimatization, sea urchins were induced to spawn by injecting 1 ml of 0.5 M KCl solution into the coelom through the peristome.
Sperm toxicity procedure
This test was developed following a similar procedure using the sea urchin S. neumayeri [53,54]. Sperm from the sea urchin was obtained as described above. A portion of 0.1 ml of sperm solution in filtered sea water (0.22 µm) was added to chambers containing 0.9 ml of filtered seawater, to achieve a total volume of 1 ml. Every day that a sperm toxicity test was conducted two negative controls and a positive control were also run: the first negative control consisted of 0.9 ml of filtered sea water; the second negative control consisted of 0.05 ml of methanol added to 0.9 ml of filtered sea water; the positive control consisted of a solution of 0.4 ml of filtered sea water, 0.5 ml of methanol. Methanol was allowed to evaporate under the hood for about 1h. Once methanol was evaporated, 0.1 ml of sperm solution was added to the chambers. Finally, tests consisted of 0.05 ml of the ether extract solved in methanol, added to 0.9 ml of filtered sea water. Each extract was tested at three arbitrary concentrations: 1mg ml-1, 0.5 mg ml-1, and 0.05 mg ml-1. Chambers with the extracts were left to evaporate the solvent under the hood for a period of about 1h. After that, 0.1 ml of the sperm solution was added to each chamber for every extract. Ten replicates at each concentration were tested and ten replicates for each of the controls were run simultaneously. Sperm mobility was checked under the light microscope (40x) 20 min after the sperm solution was added to each chamber. Extracts were considered toxic when >25% of the sperm was inactive.
Embryo toxicity procedure
The cytotoxicity test procedure was developed following to the protocol proposed by Ghirardini, et al. [55] developed using the Mediterranean sea urchin Paracentrotus lividus. This protocol is commonly used worldwide because is a simple, inexpensive and rapid procedure. The present work introduces some modifications in the original protocol, mainly focused on the volume of sea water used and the time of exposure of the extract, in order to adapt it to the particularities of the Antarctic sea urchin, S. neumayeri.
Sterechinus neumayeri gamete release was induced to at least 9 individuals in order to obtain a minimum of 3 males and 3 females with a good gamete emission. Sperm and eggs obtained were transferred separately to a 50 ml beaker with filtered sea water, thus obtaining a sperm and eggs suspension diluted in sea water. The egg suspension was diluted to a final concentration ranging from 50 to 200 eggs·ml-1 in order to test the best concentration to be used. After that, 0.1 ml of the sperm suspension was transferred to 10 ml incubation chamber with filtered sea water. After one hour, 1 ml of egg suspension was added directly to the sperm solution and left for 20 min to ensure eggs fertilization.
Fertilization success was verified by the presence of the fertilization membrane using a microscope. When a minimum of 95% of the eggs was fertilized, they were filtered (20 µm) and 0.5 ml of the solution of fertilized eggs was then exposed to the chambers test (controls and test). Every day that a cytotoxicity test was conducted, five replicates were conducted at three different concentrations (1, 0.5 and 0.05 mg ml-1) of controls and tests assays. The first control consisted of 4.5 ml filtered sea water; the second control consisted of 4.5 ml of filtered sea water with methanol. Finally, tests consisted of 4.5 ml of filtered seawater with the ether extract diluted in methanol. Methanol was allowed to evaporate under the hood for about 1 h. Once methanol was evaporated, 0.5 ml of fertilized eggs solution was added to every chamber. During the tests, the water temperature ranged between -1 to 2°C and pH = 8.
The percentage of blastula stage for each treatment was determined by counting fertilized eggs under the microscope (10 x) using a plankton counting camera. For a period of seven days, the progress of embryonic development in the sea urchin was monitored in both controls and treatments every 12-16 hours. The solution of fertilized eggs was tested to a final concentration ranging from 50 to 200 eggs·ml-1 in order to find the optimal concentration of exposed embryos to be employed for this method. In order to evaluate the mortality of fertilized eggs at different concentrations, an S regression model was performed. SPSS regression nonlinear module was used.
TThe S regression model was calculated between the percentage of embryos reaching the blastula stage (Y) and the initial concentration of eggs (X) only using the data from the values of controls to obtain a representation of the normal behavior (without the influence of the extracts) of the experimental conditions. Also, Confidence Intervals (CI) of prediction (upper and lower: UCL and LCL) of 95% coverage were calculated to detect active extract samples without the influence of mortality by the initial concentration of eggs (points outside CI).
Comparing the results of the two experiments, a high percentage (23 out of 29; 79%) of the species tested belonging to 24 out of 34 samples were toxic in one or both experiments. Regarding taxa, all cnidarians, tunicates and polychaetes showed activity. Moreover, most of the bryozoans (67%) and hemichordates (67%) were also active. The extracts of the nemertean Parborlasia corrugatus and the polychaete of genus Polyeunoa were active in both experiments. In contrast, the bryozoan Alcyonidium flabelliforme was toxic against sperm but not against embryos.
Sperm toxicity
In the sperm toxicity experiment, all the species except one (Cephalodiscus sp. 1) (94%) were toxic to sperm at the maximum concentration (1 mg ml-1) and 16 out of the 19 species tested (84%) were active at the intermediate concentration (0.5 mg ml-1). Twelve (63%) belonging to 14 out of the 25 tested samples were toxic to sperm at the concentration of 0.05 mg ml-1 (Table 1 and 2).
| Group | Identification | Body location (a) | 1 mg ml-1 | 0,5 mg ml-1 | 0,05 mg ml-1 | Activity (b) |
|---|---|---|---|---|---|---|
| ANNELIDA | Nephtyidae sp. | whole | + | + | + | ST |
| Polyeunoa sp. (1) | whole | + | + | + | ST | |
| Polyeunoa sp. (2) | whole | + | + | + | ST | |
| Polyeunoa sp. (3) | whole | + | + | + | ST/ET | |
| Travisia sp. | whole | + | + | - | ST | |
| BRYOZOA | Cellaria diversa | whole | + | + | - | ST |
| Alcyonidium flabelliforme | whole | + | + | - | ST | |
| CNIDARIA | Alcyonium haddoni | whole | + | - | - | ET |
| Alcyonium roseum | whole | + | - | - | ST | |
| Primnoidae sp. | POL | + | + | + | ST | |
| AXI | + | + | - | ST | ||
| Thouarella laxa | POL | + | + | - | ST | |
| AXI | + | + | - | ST | ||
| HEMICHORDATA | Cephalodiscus sp. (2) | whole | + | - | - | ST |
| Cephalodiscus sp. (3) | whole | + | + | + | ST | |
| NEMERTEA | Parborlasia corrugatus | whole | + | + | + | ST/ET |
| PORIFERA | Iophon sp. | whole | + | + | + | ST |
| Isodictya verrucosa | API-INT | + | + | + | ET | |
| API EXT | + | - | - | ET | ||
| Mycale acerata | whole | + | + | + | ST | |
| To be determined | whole | + | + | + | ST | |
| Rossella fibulata | BAS INT | + | - | - | ET | |
| Rossella sp. | EXT | - | - | + | ET | |
| PORIFERA-OPHIURA | Iophon sp. + Ophiurolepis sp. (1) | SPO | + | + | + | ST |
| DIS | + | + | + | ST | ||
| Iophon sp. + Ophiurolepis sp. (2) | SPO | + | + | + | ST | |
| DIS | + | + | + | ST | ||
| CHORDATA | Cnemidocarpa verrucosa (1) | FEN | + | - | - | ET |
| INT | + | + | - | ET | ||
| Cnemidocarpa verrucosa (2) | EXT | + | + | - | ET | |
| FEN | + | - | - | ET | ||
| INT | + | - | - | ET |
Table 2: Active samples from the cytotoxicity experiment against sea urchin embryos and sperm. (a) Body location: EXT: external, INT: internal, API: APICAL, FEN: gill slits, POL: polyps from the gorgonian, AXI: axis from the gorgonian, SPO: sponge, DIS: disc from the ophiura. (b) Activity tested: b) ET: embryo toxicity, ST: sperm toxicity (if both are in the list, the symbols refer to both tests). +: active; -: in active.
By taxonomical groups, all the sponges and all the polychaetes except Travisia sp., the association of the sponge-ophiura, the nemertean Parborlasia corrugatus, as well as one of the hemichordates, were toxic for the sperm of Sterechinus neumayeri at all tested concentrations. Regarding negative results, most of the cnidarians (4 of 5) and the two bryozoans tested did not exhibit toxicity when tested at the lowest concentration. One of the extracts of Cephalodiscus sp. is the only one that did not cause any effect to S. neumayeri sperm.
Regarding the body allocation of compounds, the cnidarian Thouarella laxa was active at 1mg ml-1 and 0.5 mg ml-1 concentrations in the two parts (axis and polyps), demonstrating thus, no tendency on locating secondary metabolites in the different body parts.
Interspecific variability was found in two out of the three samples of the genus Cephalodiscus (2 and 3), which were toxic against sperm. Additionally, only species 3 was active at all concentrations. In contrast, the two samples of the sponge Iophon sp. and the ophiura Ophiurolepis sp. and the three samples of the polychaete Polyeunoa sp. were toxic against sperm at all tested concentrations.
Cytotoxicity against sea urchin embryos
In the experiment of cytotoxicity during different stages of the development of S. neumayeri embryos, 7 out of 14 species tested (50%) belonging to 7 of the 17 samples did not reach the blastula stage at the highest extract concentration (1 mg ml-1), either with the entire animal extract or with some of its parts (Table 1 and 2). Only two of the species tested (the annelid Polyeunoa sp. and the nemertean Parborlasia corrugatus) did not reach the blastula stage at any tested concentration. The controls reached the blastula stage after 5-6 days. An extract of Isodictya verrucosa inhibited the development of almost all the blastula at 1 mg ml-1 (Table 2).
The results on development show a S curve (Y = exp (0.702 + 124, 928/X); R2 = 0.6125) that represents the percentage of the embryos reaching the blastula stage starting at different concentrations of initial fertilized eggs (Figure 1). In this model, a relationship between the number of the initial eggs and the embryonic mortality suggests that the embryonic mortality increases with the increase in density of initial eggs. Moreover, this model suggests that the optimal concentration of exposed embryos to be used is around 50 eggs, when the value of mortality is lower. The points below the lower range of the confidence intervals of 95% indicated the tested extracts that inhibited the blastula stage (with a minimum mortality caused by the initial concentration of eggs). Conversely, the points above the upper range represented the extracts that favor the blastula development. In total, 7 species from 14 showed activity in some of the tested concentrations (Table 2). For some extracts, the blastula stage was not inhibited but the number was significantly smaller from the control treatment.
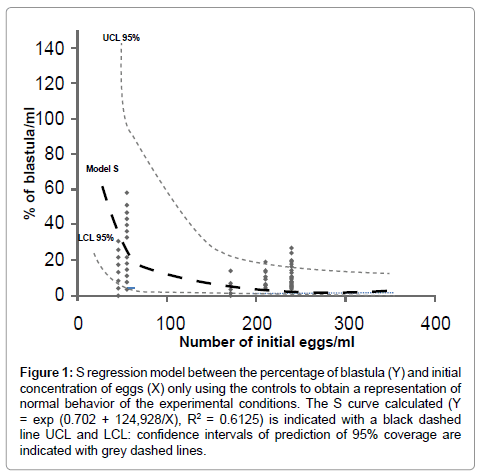
Figure 1: S regression model between the percentage of blastula (Y) and initial concentration of eggs (X) only using the controls to obtain a representation of normal behavior of the experimental conditions. The S curve calculated (Y = exp (0.702 + 124,928/X), R2 = 0.6125) is indicated with a black dashed line UCL and LCL: confidence intervals of prediction of 95% coverage are indicated with grey dashed lines.
Regarding the body allocation of compounds, one sample of the common Antarctic solitary ascidian Cnemidocarpa verrucosa was active at the concentration of 1 mg ml-1 at all tissues (external, internal and gill slits), indicating that its defensive secondary metabolites are homogenously distributed. In contrast, another specimen of C. verrucosa collected from a different station of the Weddell Sea did not present activity in the external part at the same concentration. The internal part of species 1 and the external part of species 2 were active at 0.5 mg ml-1. In this case, no activity was detected to the minimum concentration. Thus, the ascidian C. verrucosa exhibited an intraspecific variation in activity depending on location. In the case of the sponges Isodictya verrucosa and Rossella fibulata, the toxic compounds were only located in the apical or basal part, respectively. Moreover, intraspecific variability was not detected in three samples of the sponge Scolymastra joubini tested.
Our results demonstrate that a high percentage (79%) of the species tested (23 out of 29) were active in one or both cytotoxic experiments, thus revealing that the majority of Antarctic marine organisms tested are toxic to sperm and early life stages of the Antarctic sea urchin S. neumayeri. All cnidarians, tunicates and polychaetes and most of the bryozoans (67%) and hemichordates (67%) were active, demonstrating high cytotoxic activity in the majority of groups tested. However, due to uneven sampling efforts, the percentages of activity in the different phyla are not comparable. A similar activity in most of the cnidarians and bryozoans tested was described in temperate areas by Martí, et al. [56]. In that study, only 36% of the sponges were toxic, contrasting to the high percentage of bioactive sponges species found in other studies [57,58]. Moreover, the extracts tested here revealed an evident toxic effect by causing 100% mortality of embryos in some concentrations of eleven extracts, and cytotoxicity against sperm was especially remarkable, with activity in 94% of the tested species and also observed in 63% of species at the minimum concentration tested. Generally, marine invertebrates with pelagic larvae settle on or near their prey [59] and the potential negative effects of extracts on sea urchin displayed in our experiments could be a relevant chemical mechanism of defense or competence against this species reducing the recruitment of this sea urchin and, consequently, the grazing pressure and the colonization of the surface [17,49]. Remarkably, a few species did not show any activity, thus indicating they may have other types of mechanisms of defense/competence. To our knowledge, only a few studies have reported that embryonic development of sea urchins, as the Mediterranean Paracentrotus lividus, may be inhibited by organic extracts from different benthic invertebrates [57]. Conversely, few studies have proved the potential activity from the extracts of Antarctic benthic organisms against the eurybathic S. neumayeri, and even less from deep-water organisms [43,49,53,54].
Porifera is one of the most represented groups in Antarctic benthic communities [8,60]. Although most chemical ecology research has usually focused on this phylum, in temperate and tropical waters [31], few compounds from Antarctica have been tested for their ecological functions [23,32]. In particular, sponges from other regions, infrequently fouled, have an abundance of novel compounds, some of them with interesting ecological functions as to deter invertebrate larval settlement [59]. In our study, Mycale acerata showed toxicity against sperm in all concentrations, revealing a strong activity. However, M. acerata did not show anti-predatory activity against the Antarctic sea star O. validus, demonstrating that these toxic compounds are not repellents [33]. Similar toxicity results have been observed in mycalosides isolated from congeneric Caribbean sponge M. laxissima, which inhibited the fertilization of eggs by sperm of the sea urchin Strongylocentrotus nudus [61]. On the other hand, other studies carried out by Peters and collaborators [19] showed that M. acerata inhibited the growth of the bacteria Alteromonas elyakovi. Moreover, different species of the genera Iophon and Isodictya caused mortality in diatoms. In our case, one species of Iophon and Isodictya verrucosa also showed a high toxicity against sperm and embryo, respectively. Also, the toxic compound(s) of I. verrucosa against embryos tested here are only located in its apical part. This result is in agreement with the hypothesis based on the accumulation of defensive compounds in external body tissues of sponges. According to the Optimal Defense Theory (ODT), the localization of compounds in the apical part could prevent settling of larvae, which could make filtration difficult. In different studies, it was found that the congeneric sponge Isodictya erinacea possessed feeding deterrents against sea stars and amphipods, as well as inhibitors of fouling or infectious organisms [33,62,63], thus representing a genus potentially active. Also, the extracts of one species of Rossella, R. fibulata, tested here were toxic against embryo development. This species was rejected as food by the Antarctic amphipod Cheirimedon femoratus [64], showing thus, the presence of both antifeeding and cytotoxic activity. Additionally, bioassays in congeneric species of Rossella from Weddell Sea showed cytotoxicity against human tumour cell lines [65], feeding repellence against C. femoratus [64] and deterrent activity against Antarctic sea stars [21,33,66,67]. Therefore, this genus seems to be well protected by diverse chemical strategies.
Cnidarians are also generally well represented in shallow and deepwater benthic fauna in the southern polar region [1]. New compounds of cnidarians, especially octocorals (soft corals and gorgonians), have been discovered recently from diverse areas [68-70]. In our study, all octocorals tested were toxic against at least one experiment. In particular, our assays indicated the existence of cytotoxic compounds against embryos and sperm in the Antarctic soft corals Alcyonidium haddoni and A. roseum, respectively. Moreover, these species were unpalatable towards the Antarctic amphipod Cheirimedon femoratus [64], demonstrating the existence of compounds with diverse roles, or alternatively, different strategies. Different metabolites have been isolated from congeneric species of Alcyonium of diverse Antarctic regions [23]. In particular, Slattery and collaborators [43] demonstrated that the Antarctic congeneric species A. paessleri exhibited an antimicrobial activity, and this species also was toxic to larvae of S. neumayeri. Soft corals lack a calcium carbonate skeleton, although they contain internal sclerites, thus these chemical strategies are a very useful defense. The flexible gorgonian Thouarella laxa was also active against the sperm in our study, and rejected by C. femoratus [64], representing thus an active species possessing cytotoxic and repellent mechanisms.
Ascidians are considered significant elements in the Antarctic benthic communities [71]. Although this group is also an important source of natural products with pharmaceutical properties [23], few studies have focused on the chemical interactions that these compounds play in the ecosystems [19,30,33,64,72,73]. In particular, some colonial and solitary ascidians possess chemical defenses to protect them from predators [20,64,72]. Although the Antarctic solitary tunicate Cnemidocarpa verrucosa is known to be fouled by different invertebrates, it seems to be well defended in our study against both the development and the sperm of S. neumayeri. In recent studies, the tissues of C. verrucosa proved to be unpalatable to O. validus [35,73]. Conversely, in another study this species did not show antifeeding activity against the same sea star [33]. Considering the specific body allocation of the compounds of this species, one out of the two samples of C. verrucosa possessed activity in all tissues and therefore, no tendency on locating secondary metabolites in the body parts. In contrast, another specimen of C. verrucosa collected from a different station of the Weddell Sea did not present activity in the external part at the same concentration, indicating intraspecific variability in reference to the location of the toxic compounds. This variability could be related to the ascidian itself or to possible symbionts. This interesting issue remains to be further investigated.Bryozoans of the Southern Ocean comprehend a rich and varied fauna [8]. Some species from diverse regions along the world have demonstrated to be good sources of pharmacologically interesting substances, such as cytotoxic alkaloids [74]. However, research on chemical ecology of bryozoans continues to be limited and even more, in Antarctica [23,32]. In our study, Alcyonidium flabelliforme showed cytotoxic activity against the sperm of the sea urchin. Alcyonidium, being a ctenostome, shows a soft and fleshy texture and therefore, this genus is not protected by a calcarious skeleton.
This supports the hypothesis that their compounds may represent a chemical protection. In agreement with the activity of this ctenostome, a recent study demonstrated that this species also displayed significant repellency towards an Antarctic sea star, Odontaster validus [33]. Therefore, this species may show different defense chemical strategies against diverse Antarctic predators. In particular, two species of the genus Alcyonidium from diverse regions produce bioactive compounds [74]. For example, the encrusting species A. nodosum that lives on the shells of marine gastropod Burnupena papyracea prevents predation of the whelk by the rock lobster Jasus lalandii [75]. In our study, Cellaria diversa also presented activity against the sperm of S. neumayeri. In agreement with this, the extracts of a congeneric species, C. pilosa, showed antibacterial activity [76]. Moreover, this species was repellent to the sea star O. validus [33], suggesting also the presence of diverse chemical mechanisms. Additionally, another study tested the haemolytic activity of five species of Antarctic bryozoans against erythrocytes derived from different mammals, and the only active species was the weakly calcified Carbasea curva, which lacks avicularia [77]. The lack of these mechanical defenses may be the related to the chemical activity. In contrast, in our results, the weakly calcified bryozoan Klugeflustra antarctica of the same family (Flustridae) was not toxic against embryos of sea urchin, while this species also lacks avicularia and has not other obvious mechanical defensive structures. Therefore, other chemical mechanisms could act against the predation pressure by this sea urchin, such as the possession of repellent compounds.
Few studies investigating the chemical defenses of cold-water marine polychaetes have been done so far [23,32]. In our results, the polychaete Polyeunoa sp. (Polynoidae) was active at all concentrations in both experiments, demonstrating that this organisms show a high potential biocide activity against at least two strategies of defense (embryo and sperm toxicity). In contrast, the Antarctic species P. laevis did not cause repellence against the Antarctic sea star Odontaster validus [33]. This species is frequently found in symbiotic association with gorgonians of the genus Thouarella, suggesting their use as host gorgonians, being a potential chemical shelter to avoid predation [78]. Our results would be in agreement with this. On the other hand, Travisia sp. was active against the sperm in our tests, and showed a strong deterrent activity against the sea star O. validus [33], being thus a well-protected species.
In the case of Antarctic nemerteans, Parborlasia corrugatus was active at all concentrations in both experiments, indicating also a high level of activity and the presence of a minimum of two types of cytotoxic mechanisms. Heine, et al. [53] exposed also extracts of P. corrugatus to the sperm of S. neumayeri. Sperm was killed at the highest concentration and they suggested that the low pH (3.5) of the nemertean epithelial mucus might be responsible for this toxicity. Conversely, this species did not display significant deterrent activity against the Antarctic sea star Odontaster validus [33]. However, our study suggested that probably not only the low pH may defend this organism against this sea urchin, but also the presence of toxic natural compounds, which were present in the extracts tested here.
As for hemichordates, two of the three samples of Cephalodiscus were active, demonstrating cytotoxic activity and variability in this genus. In agreement with this, a remarkable cytotoxic activity against certain human cancer cell lines was exhibited by a cephalostatin isolated from an Indian congeneric species, C. gilchristi [79]. In contrast, another sample of an Antartic Cephalodiscus did not exhibit repellence against the Antarctic sea star Odontaster validus [33]. This could be related to the habits of this genus, living usually partially buried in the sediment.
In this study, we tested extracts of deep-water organisms from the Weddell Sea, a region that has received little attention in the past. Additionally, our study reveals the existence of cytotoxic compounds in many diverse benthic organisms. Thus, both activities (sperm and embryos toxicity) may represent a suitable combination of strategies against this sea urchin, being these chemical defenses very useful to prevent its settlement. Moreover, the intraspecific differences found in some of tested species may indicate the responses to different microhabitats, diet or bacteria associated, as well as prove that diverse congeneric species can possess different active compounds with different functions. Further work is in progress to test more extracts and to determine the chemical compounds involved in these activities.
We are thankful to L. Núñez-Pons and J. Cristobo for their laboratory support. In this work we used the extracts and isolated compounds from previous projects (ECOQUIM projects), for this reason we want to thank W. Arntz and the R/V Polarstern and the BIO-Hespérides crews. Thanks are also due to the taxonomists for the identification of samples: J. Cristobo and P. Ríos (Porifera), L. Núñez-Pons (Cnidaria), M. Ballesteros, A. Bosch and N. Campanyà (Echinodermata). We would like to thank also the Unidad de Tecnología Marina (UTM) and the crew of Las Palmas vessel for all their logistic support. Special thanks are also given to the “Gabriel de Castilla BAE” crew for their help during the ACTIQUIM-1 and -2 Antarctic expeditions. This research was developed in the frame of the ACTIQUIM projects (CGL2007-65453/ANT, CTM2010-17415/ANT). Finally, we would like to thank PharmaMar for financially supporting some of the authors during part of this work and Banco Popular for providing video equipment.