Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Editorial - (2014) Volume 2, Issue 2
AUXIN BINDING PROTEIN1 (ABP1) has long been proposed as a candidate of auxin receptor to mediate auxin action in plants. It is essential for plant organ development. The function of ABP1 in the development of ramie (Boehmeria nivea L.) remains unclear. Here, we examined the role of the ABP1 gene in the development of shoot in ramie, especially in leaf venation ontogeny, by transforming ABP1 antisense construct into the plants. We showed that ABP1 expression was decreased in ABP1 antisense transgenic plantlets. The decrease of ABP1 expression led to defects in plant growth including dwarf plant, and decreased apical dominance in the intact plant. In addition, ABP1 antisense lines exhibited a serious retard of leaf development involving small and twisted leaves, sparse trichome, and a decreased cell expansion. Moreover, the decreased ABP1 expression repressed the development of leaf venation. ABP1 antisense lines developed an incomplete leaf venation, which became smaller and lacked in some leaf tissues. Though the cell arrangement was normal in midrib and lateral vein, cell size was decrease and some cell layers were missing in midrib. The abnormal phenotype of leaf venation was attributed to the decreases in cell number and size. Our data support that ABP1 is necessary for shoot growth in ramie. More importantly, it plays a key role for the development of ramie leaf venation by regulating both cell expansion and division.
Keywords: Boehmeria nivea L.; Auxin binding protein 1; Expression decrease; Shoot development; Leaf venation
The phytohormone auxin is critical for plant growth and triggers many developmental processes [1]. Its effects on plant development are mediated by two distinct signaling pathways: the nuclear TIR1/AFB pathway and the ABP1 dependent pathway [2]. The nuclear TIR1/AFB pathway is well-characterized, controls a remarkable number of auxin mediated process, and plays an important role in plant development [3,4]. However, the ABP1 dependent pathway remains elusive until now. It has shown that ABP1 may be an important candidate for auxin receptor, and essential for a set of auxin responses, including early auxin response at the plasma membrane, cell division and expansion, and auxin regulated gene expression [5,6]. It is important throughout the life of plants, such as embryogenesis and postembryonic shoot and root development [7]. But there are still many unresolved problems concerning the molecular function of ABP1.Thus it still has attracted widespread attention of researchers.
Leaf venation, or leaf vascular tissue, which acts as a conduit of both nutrient and signaling material, is a network of cells organized in different pattern [8]. The pattern and ontogeny of leaf venation are precisely regulated by a variety of substances [9,10]. Auxin has been implicated in controlling all stages of vascular strand formation [11-15]. Auxin distribution and transportation were visualized in Arabidopsis leaf vascular development by using expression analysis of auxin carrier gene PINs and the synthetic auxin responsive promoter DR5 [11-12,15]. The results have shown that auxin accumulates in the positions of the epidermis, at where vascular tissue is differentiated. Vascular differentiation is obstructed in the presence of auxin transport inhibitors [12]. Many of leaf venation pattern mutants exhibit auxin defects, and contain mutations in genes involved in auxin response or polar auxin transport [16-18]. Many auxin response mutants reveal vascular defects in the leaf [19].
It is reported that mutations in genes, such as auxin response factor 5 (AFR5), auxin resistant 6 (AXR6), and bodenlos (BDL)/IAA12, which are involved in the nuclear TIR1/AFB pathway, have resulted in incomplete vascular systems and defects [17,19,20]. However, little is known about the role of ABP1 in the pattern and ontogeny of leaf venation.
In this study, we investigated the role of ABP1 gene in ramie shoot development, especially in leaf vascular development, by transforming ramie plants with ABP1antisense expression gene construct. ABP1 antisense lines had abnormal phenotypes. Leaf vascular tissue was incomplete or smaller. The abnormal phenotypes of leaf veins were attributed to the decreases in cell size and number. Thus we conclude that the auxin-ABP1 signal pathway is involved in shoot growth in ramie. More importantly, it is necessary for the development of ramie leaf venation.
Construction of ABP1 antisense expression vector
The coding region of ABP1 cDNA from ramie (Boehmeria nivea L., cv. Xiangzhu 3) was amplified by PCR using both the forward primer (BnABP1F:5’-GCTCTAGAATGGGTTGGTCTTCGATTCC-3’) and the reverse primer (BnABP1R:5’-CGGGATCCTTACAGTTCATCC TTTTGAT-3’), digested with XbaI and BamHI and then cloned into the pWM101 vector downstream of the 35S promoter in the antisense orientation (Figure 1). After sequencing to verify the sequence, the construct was inserted into Agrobacterium tumefaciens LBA4404 by electroporation.
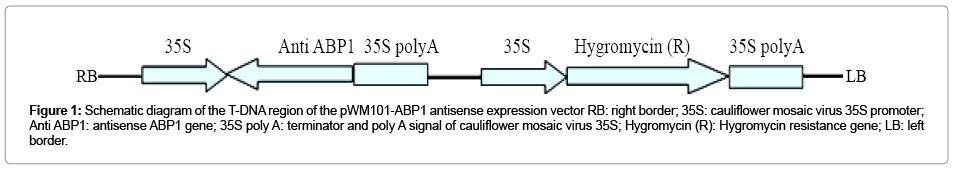
Figure 1: Schematic diagram of the T-DNA region of the pWM101-ABP1 antisense expression vector RB: right border; 35S: cauliflower mosaic virus 35S promoter; Anti ABP1: antisense ABP1 gene; 35S poly A: terminator and poly A signal of cauliflower mosaic virus 35S; Hygromycin (R): Hygromycin resistance gene; LB: left border.
Plant transformation and growth
A ramie cultivar “Xiangzhu 3” was used as transformation material. The seeds were germinated on half-strength MS medium according to the method as described by Wang [21]. The leaves were used for transformation at the upper one to two nodes of 10d old seedlings. The plant transformation and regeneration were carried out essentially according to the procedure described previously [21]. The medium for pre-culturing explants were MS medium supplemented with 0.5 mg.L-1 thidiazuron (TDZ), and 0.01 mg.L-1 IAA. The selection regeneration medium were MS medium supplemented with 0.5 mg.L-1 TDZ, 0.01 mg.L-1 IAA, 15 mg.L-1 hygromycin and 500 mg.L-1 cefotaxime. The rooting medium was 1/2 MS medium supplemented with 8 mg.L-1 hygromycin and 0.05 mg.L-1 NAA. Rooted shoots with four leaves were transplanted to pots containing soil, vermiculite, and sand (2:1:1, w/w/w) for two weeks, and then moved to the greenhouse for further growth. The hygromycin resistant transgenic plants were identified by PCR.
All cultures were maintained at 25°C with 16h of photoperiod provided by cool-white fluorescent lights (Phillips, Holland) at a light intensity of 37.5 umol m-2 s-1. Controls and transgenic lines were grown under natural light in the greenhouse at Hunan agricultural university.
Genomic DNA Isolation and PCR
Genomic DNA was isolated from young leaves of transgenic lines and non-transgenic plants using the CTAB method. A pair of primers was used to determine the presence of the 35S promoter and the ABP1 transgene: 35S-F(5’-AGTGGATTGATGTGATATCTCCACT-3’)and BnABP1 (5’-GGTCTGGAATACAAACGAAC-3’). This primer pair generated a PCR product at 356 bp.
Semi-quantitative RT-PCR Analysis
Total RNA was isolated from young leaves of transgenic lines and non-transgenic plants using the total RNA isolation system (Promega, USA). cDNA was synthesized by RevertAid First Strand cDNA Synthesis Kit (Fermentas, Canada). For semi-quantitative RTPCR analysis, the cDNA sample was used to amplify an ABP1 segment with primers: BnABP1 (5’-GGTCTGGAATACAAACGAAC-3’) and BnABP2 (5’-GTCATATACATGATTATCTATG-3’). Actin (GenBank : DQ665832) was used as the internal control with primers BnActin1 (5’-GTTGAACCCTAAGGCTAACAGAG-3’) and BnActin2(5’- TCAGCTCCGATTGTG ATGAT-3’).
Growth Measurements and Microscopy
Potted after four weeks, the transformed phenotypes were clearly distinguishable from the control plants. Growths were measured and were recorded as total heights. Leaves and internodes were numbered basipetally. The trichomes on leaves at the first node of main stems were observed. For light microscopy, the leaves at the third node of main stems were collected and embedded into paraffin after formaldehyde acetic acid fixation and progressive dehydration. Leaf venation cross sections stained with safranin and fast green, were cut using a microtome, and visualized with an Olympus microscope.
Generation of transgenic ramie plantlets
ABP1 gene fragment was introduced into the pWM101 vector downstream of the 35S promoter in the antisense orientation to generate the ABP1 antisense expression construct (Figure 1). And then the binary construct was delivered into Agrobacterium tumefaciens strain LBA4404. Ramie plants were transformed with ABP1 antisense expression construct by means of Agrobacterium mediator. Twenty four hygromycin-resistant plants (T0 generation) were obtained. To ensure that ABP1 antisense expression vectors were introduced into the resistant plantlets, a primer pair was targeted inside the 35S promoter and the ABP1 transgene, and used to amplify a 356bp fragment from the resistant plant genome (Figure 2A). Eighteen resistant plantlets showed PCR positive. The levels of ABP1 in leaves of transgenic plants were detected by semi-quantitative RT-PCR analysis. The results showed that the transcript levels of ABP1 in four independent transgenic lines (3, 5, 6 and 8) were significantly reduced (Figure 2B), which indicated that the expressions of the ABP1 gene were down-regulated by antisense constructs. The others (9 and 10) had the expression levels of ABP1 indistinguishable from wild type, and were used as negative controls in experiments presented here.
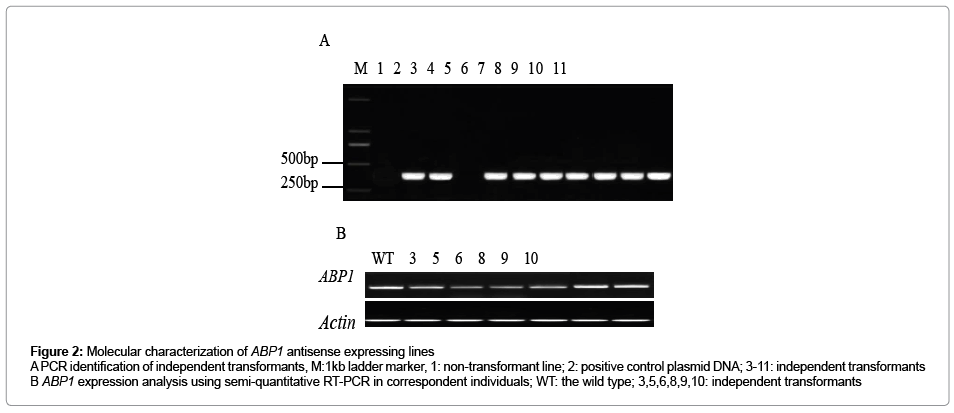
Figure 2: Molecular characterization of ABP1 antisense expressing lines A PCR identification of independent transformants, M:1kb ladder marker, 1: non-transformant line; 2: positive control plasmid DNA; 3-11: independent transformants B ABP1 expression analysis using semi-quantitative RT-PCR in correspondent individuals; WT: the wild type; 3,5,6,8,9,10: independent transformants
Decrease of ABP1 expression led to repression of plant growth
ABP1 antisense lines showed dramatic growth reduction compared with control seedlings (Figure 3A-3E).
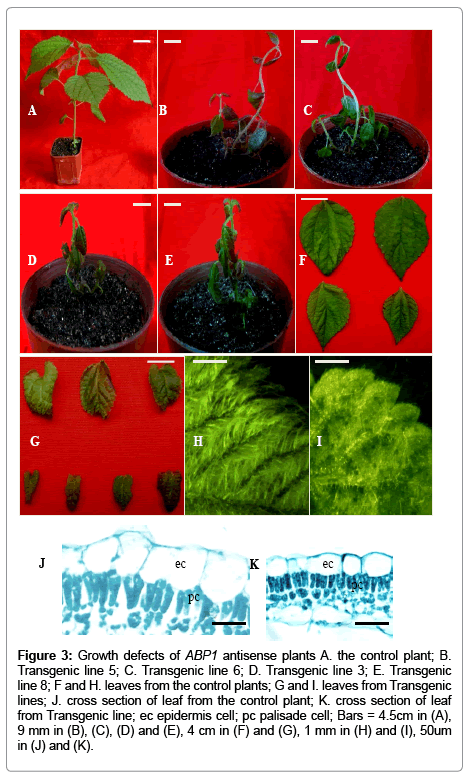
Figure 3: Growth defects of ABP1 antisense plants A. the control plant; B. Transgenic line 5; C. Transgenic line 6; D. Transgenic line 3; E. Transgenic line 8; F and H. leaves from the control plants; G and I. leaves from Transgenic lines; J. cross section of leaf from the control plant; K. cross section of leaf from Transgenic line; ec epidermis cell; pc palisade cell; Bars = 4.5cm in (A), 9 mm in (B), (C), (D) and (E), 4 cm in (F) and (G), 1 mm in (H) and (I), 50um in (J) and (K).
ABP1 antisense lines potted after four weeks reached the heights of 9.5cm, whereas the controls attained the heights of 25.5cm. ABP1 antisense lines had decreased apical dominance (Figure 3A-3E). In addition to this general reduction in growths, leaf phenotypes were also affected. Leaves of ABP1 antisense lines displayed decreases both in lamina width and length, curling, and reduced leaf trichomes (Figure 3F,3G,3H and 3I). To research which cellular alterations were correlated with leaf growth defects, we performed an analysis of leaf histology. Histological analysis of leaves from ABP1 antisense lines showed that all expected cell types were present and that these were arranged in an appropriate pattern, but some epidermis cells and palisade cells became smaller (Figure 3J and 3K).
Decrease of ABP1 expression repressed leaf vein development
The ABP1 antisense lines showed suppression of leaf vein development (Figure 4). The patterns of leaf venations were abnormal (Figure 4A-4D). And the leaf venations became smaller and obscure. In addition, some regions of leaves exhibited lacks of leaf venations. To investigate whether the observed leaf venation phenotypes of ABP1 antisense plants were related to altered internal structures of the leaf venations, we performed a histological analysis of leaf venations. The defects in the development of leaf venations were clearly visible in transverse sections (Fig 4E-4J). In the cross sections of the midribs of the transgenics, though the phloem, parenchyma and xylem were arranged in an appropriate pattern, the cells were all less in amount and smaller in size, and the collenchyma cells were a single layer, whereas they were several layers in the midrib of the controls (Figure 4E and 4F). Similar changes also were observed in histologys of the lateral veins (Figure 4G-4J). The decreases in cell number and size resulted in the abnormal phenotypes.
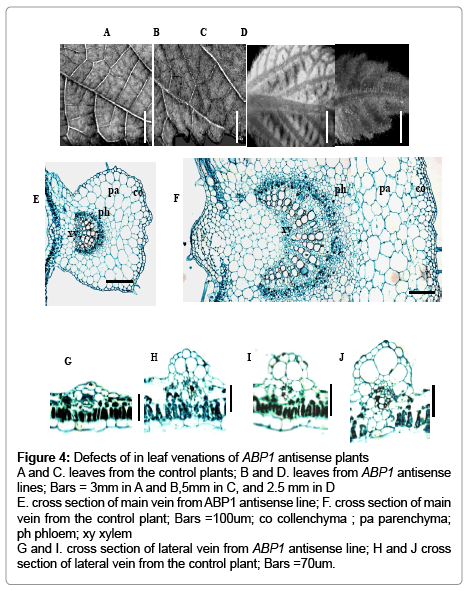
Figure 4: Defects of in leaf venations of ABP1 antisense plants A and C. leaves from the control plants; B and D. leaves from ABP1 antisense lines; Bars = 3mm in A and B,5mm in C, and 2.5 mm in D E. cross section of main vein from ABP1 antisense line; F. cross section of main vein from the control plant; Bars =100um; co collenchyma ; pa parenchyma; ph phloem; xy xylem G and I. cross section of lateral vein from ABP1 antisense line; H and J cross section of lateral vein from the control plant; Bars =70um.
ABP1 has long been prospected as an important mediator of auxin action [5-7]. Because the null abp1 mutant confers embryo lethality, it frustrates any conclusions on ABP1’s role during plant post embryo development [22]. By taking advantage of weak alleles and an antisense approach, ABP1 is proved to act as an important role in organ development in plant [23-26]. In this article, we used an antisense approach to decrease ABP1 gene expression level in ramie. Decrease of ABP1 expression led to a repression of plant growth. ABP1 antisense lines displayed decreased apical dominance, resulting in dwarf statures. Apical dominance depends on the polar auxin transport [27,28]. It is likely that the loss of apical dominance may be ascribed to a defect related to auxin physiology in ABP1 antisense lines. Leaf size and shape of ABP1 antisense lines were both altered, revealing that ABP1 is essential for appropriate leaf growth and development of ramie. This observation is in accordance with the previous reports [22-23,29,30]. The transgenic Arabidopsis plants, in which the activity of ABP1 can be conditionally repressed, reveal occasionally a lack of vascular tissue, the occurrence of double vascular mid-ribs or twisted veins in the distal tip of the leaves [23]. We have shown that reduced levels of ABP1 expression were associated with the abnormal development of leaf venation, suggesting that ABP1 has a role in development of leaf venation, thus complementing the previous results [23]. The abnormal phenotypes of leaves and leaf venations were attributable to decreases in size and number of cells. The decreased number and size of cells indicated that cell division and expansion were impaired in ABP1 antisense lines. An involvement of ABP1 in the control of cell division and expansion is consistent with the previous reports [22-24,29-30]. The processes of cell expansion and division depend on distinct and finely tuned levels of intracellular auxin. We measured endogenous auxin content in leaves of ABP1 antisense lines and the controls. No significant differences were observed, suggesting that no change in auxin content in leaves of ABP1 antisense lines occurred. It has been shown that basipetal auxin transport is reduced in heterozygous abp1/ABP1 mutant roots [25]. Auxin transport in cells are largely regulated by the PM-residing PIN auxin efflux carriers transporters [31-33]. Membrane vesicles carrying PIN proteins undergo dynamic recycling to and from the PM and auxin regulates its own transport by inhibiting the endocytic step of this recycling [34-36]. It is believed that ABP1 mediates the effect of auxin on endocytosis and that ABP1 can control the clathrin-mediated internalization of PIN proteins, which regulate the amount and dynamics of PIN auxin efflux carriers at the PM [37-41]. It is likely that ABP1 has an influence on cell expansion and division through being involved in the regulation of polar auxin transport, which affects leaf growth and vascular patterning in ramie. Moreover, ABP1 inactivation can decrease mRNA accumulation of D-type CYCLINS and CYCB1.1, which are early regulators of the G1/S transition and the G2/M marker, and hinder the G1/S and G2/M phase transitions to inhibit cell division [23-24,42-43].It has been shown previously that ABP1 inactivation results in a reduced level of endoreplication [23]. Endoreplication contributes to leaf cell expansion, and a positive correlation has been shown between endopolyploidy and cell size [44]. Thus, it is probably that decreased ABP1 expression in ramie directly effects cell division and endoduplication, which results in decreases in size and number of cells. Additionally, decreased ABP1 activity repressed trichome development. Leaf trichomes are derived from the epidermal cells and often undergo endoduplication and rapid elongation [45]. The mutant phenotype in leaf trichomes may result from defects in endoreplication and cell expansion.
In conclusion, our results provided direct evidence that the expression of ABP1 gene in ramie is crucial for the development of shoot, especially leaf vein. The ABP1 as an auxin receptor is involved in auxin polar transportation regulation and auxin mediated cell expansion and division, thus functions in the development of leaf venation.
This work was supported by grants from the National Science Foundation of China (31071457), the Innovative Fund of Graduate Student of Hunan Provincial Education Department (11C0665), the Fund of Scientific and Technological Plan of Hunan Province (2012NK3062), and the Fund of Hunan Provincial Key Laboratory for Germplasm Innovation and Utilization of Crop (12KFXM11).