Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2017) Volume 8, Issue 4
Examination of some Teleost fishes captured in the Maga detention lake located in the Far North Region of Cameroon, revealed the presence of three new species of Myxosporidia of the genera Myxidium Bütschli, 1882 and Myxobolus Bütschli, 1882 of which complete description is given in the present study. These species are: Myxidium tetraodoni sp. nov., parasite of the urinary bladder of Tetraodon lineatus Linnaeus, 1758 (Tetraodontidae) that form ellipsoidal spores with a turgid middle part and rounded ends. They measured 11.6 (10.5-12.5) μm long × 8.2 (7.2-9.6) μm broad; the spherical polar capsules are of equal size and measure 3.7 (3.0-4.3) μm. Myxidium anisocapsularis sp. nov., a parasite of the gall bladder of Distichodus engycephalus Günther, 1964 (Distichodontidae) form spindle-shaped and elongated spores, that measure 15.2 (14.0-16.2) μm longx6.0 (5.0-6.7) μm broad; its polar capsules are quite unequal and respectively measure 6.0 (5.0-6.5) × 3.3 (3.0-3.8) μm for the larger and 4.7 (4.0-5.5) × 3.0 (2.3-3.3) μm for the smaller. Myxobolus magai sp. nov., a gill parasite of Labeo batesii Boulenger, 1911 (Cyprinidae) form ovoid spores with the anterior end larger with small protuberance, that measure 10.6 (9.0-12.0) × 6.3 (5.5-7.0) μm. Its polar capsules measure 2.8 (2.4-3.4) × 2.3 (2.0-3.0) μm.
Keywords: Myxozoa; Myxidium tetraodoni sp. nov.; Myxidium anisocapsularis sp. nov.; Myxobolus magai sp. nov. ; Parasite; Freshwater fish; Cameroon
Due to the increasing needs for animal protein production worldwide, fish industry is a major activity in human populations today [1]. However, fishes are vulnerable to various parasitic infections, including Myxosporidiosis. Myxosporidia can infect numerous fish organs and cause a wide variety of damages [2]. The extent of damage to host tissues and organs depend on the parasite species, its life cycle, infection intensity, and host response [3]. Many pathogenic species can be responsible of severe epizootics in fish farms and hatcheries. Indeed, in natural and artificial environments, Myxosporidia can cause the loss of production and death of fishes. Some parasitized fishes are unsightly and considered unfit for human consumption. Human health can be affected by Myxosporidia when immunodepressed people are contaminated with infected fish [4].
Knowledge of fish parasites is a prerequisite for a rapid diagnosis of the pathogen responsible for epizootic diseases. Early diagnosis can lead to preventive measures that remain the best way to reduce epidemics [5]. Thus it is necessary to record and report new parasites and pathological conditions when they are discovered because such information may be useful in the future as a baseline data for assessing the ecosystems health in the face of global warming [6]. Based on the morphological and metric characteristics of the spores, the number of Myxosporidia described around the world is estimated at 2200 species belonging to 64 genera and 17 families [7].
Eiras et al. [8,9] provided a synopsis of 905 species of Myxobolus Bütschli [10]; this genus has the largest global distribution and the high number of Myxosporidia species. With more than 232 species, the genus Myxidium Bütschli [10] is numerically the second genus of Myxosporidia [11]. Species of the genus Myxidium are predominantly coelozoic (rarely histozoic) infecting the gall bladder, the urinary bladder and the urinary tubules of the kidneys [2,12,13]. The number of Myxosporidia parasites of African freshwater fish is estimated at about 270 species [14,15]. Those of the freshwater fishes in Cameroon are represented by about 80 species, with forty of them belonging to the genus Myxobolus and nine to the genus Myxidium [14,16,17]. Taxonomic studies and description of new species of Myxosporidia throughout the world suggested that the diversity of species in this group of parasites is largely underestimated.
During a study of Myxosporidia parasites of teleosts of great importance of food and economic in Cameroon, we found three new species whose complete descriptions are given in the present paper. These species are: i sp. nov. a parasite of Tetraodon lineatus Linnaeus, 1758 (Tetraodontidae), Myxidium anisocapsularis sp. nov. found in Distichodus engycephalus Günther, 1964 (Distichodontidae) and Myxobolus magai sp. nov. a gill parasite of Labeo batesii Boulenger, 1911 (Cyprinidae).
Fish examined were sampled in the Maga detention lake from March to May 2016. Maga is a small village of the Mayo-Danay Division (situated between latitude 10° and 13° North and longitude 14° and 16° East), in the Far North Region of Cameroon. The Maga reservoir lake is located at 77 km from the city of Maroua and covers an area of 39000 hectares. The climate in this locality is the Sahelo- Sudanian type, which is characterized by a long dry season from October to April and a short rainy season from May to September. The average temperature is 28°C [18]. Maga is located in a floodplain and its vegetation consists of the shrub savannah (weakly wooded) and the herbaceous steppe with grass. Its soil, predominantly clay-sandy, take a clayey-silty shade around the lake [18].
Fish were captured using gill nets. In the field, once the fish were caught, a buttonhole was made on the ventral surface of each host individual. The latter were immediately immersed in a 10% formalin solution contained in a plastic container, before being brought to the laboratory. Systematic position of sampled fishes was taken from Lévêque et al. [19,20]. The species captured are: Tetraodon lineatus (Tetraodontidae), Distichodus engycephalus (Distichodontidae) and Labeo batesii (Cyprinidae).
In the laboratory, the fish were firstly examined with naked eye (eyes, fins, operculum, scales, skin) and then with the Olympus BO61 binocular stereoscopic microscope for the presence of cysts. After dissection of the fish hosts, internal organs such as gills, heart, liver, kidneys, spleen, gallbladder, gonads, intestine and urethra were taken individually and examined. The contents of the cysts were identified with the objective 100X of an IVYMEN light microscope. The content of the gall bladder, urinary bladder and swim bladder was also examined between glass and glass cover slide under the microscope. Smears of kidneys, spleen, liver, gonads, heart and urethra were carefully examined at the 40X objective of the microscope. Spore smears were fixed with methanol and stained with May-Grünwald- Giemsa. Drawings of fresh spores were performed using a Wild M-20 microscope equipped with a camera Lucida. Measurements were carried out on 50 spores using an objective micrometer. Variables taken into account are those proposed by Lom et al. [21]. Microphotographs of spores were performed using an Olympus BH-2 microscope equipped with a microphotograph device.
Myxidium tetraodoni sp. nov.
Vegetative form: Circular and polysporous plasmodia are found in the urinary bladder of the host. Ectoplasm is clear and smooth. In the granular endoplasm, spores are formed in pairs within pansporoblasts (Figure 1).
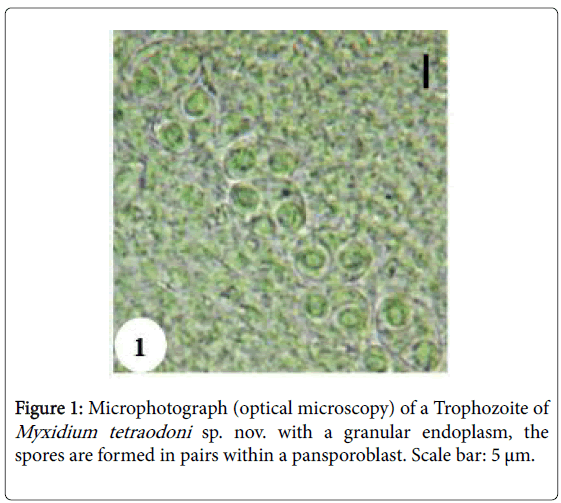
Figure 1: Microphotograph (optical microscopy) of a Trophozoite of Myxidium tetraodoni sp. nov. with a granular endoplasm, the spores are formed in pairs within a pansporoblast. Scale bar: 5 μm.
Spores: Of small size (11.6 μm length on average), spores are ellipsoidal with a bulging middle part and rounded ends (Figures 2, 10 and 11). Shell valves striated, one can count 9 to 12 fine striations on each (Figure 11). Polar capsules are spherical and equal (Figures 3 and 11). Within each of them, 4 to 5 coils of polar filament can be found (Figure 10). A granular sporoplasm is located between polar capsules.
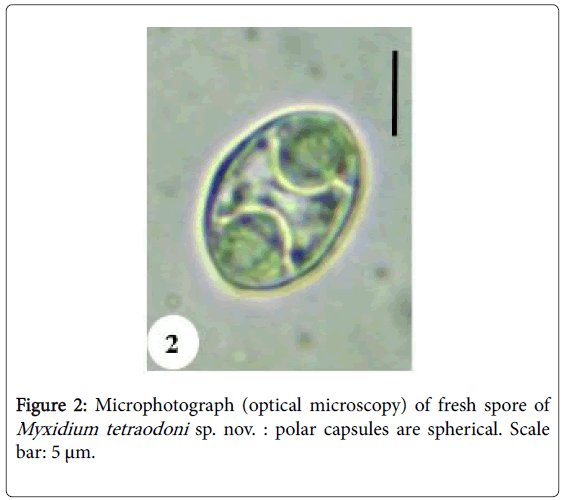
Figure 2: Microphotograph (optical microscopy) of fresh spore of Myxidium tetraodoni sp. nov. : polar capsules are spherical. Scale bar: 5 μm.

Figure 3: Microphotograph (optical microscopy) of spores of Myxidium tetraodoni sp. nov. stained with May-Grünwald-Giemsa. Scale bar: 5 μm.
Measurements of the spore: (Table 1).
| Myxidium species | Host | Site of infection | Country | LS | WS | PC | LPC | WPC | FC | StV | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| M. tetraodoni | Tetraodon lineatus | urinary bladder | Cameroon | 11.6 (10.5–12.5) | 8.2 (7.2–9.6) | = | 3.7 (3–4.3) | 3.7 (3–4.3) | 4–5 | 9–12 | Present study |
| M.brienomyri | Brienomyrus brachyistus | gall bladder | Cameroon | 13.7 (12.2–16.2) | 6.5 (5.5–9) | = | 4.2 (3.5–5) | 4.2 (3.5–5) | 3–5 | 6–12 | Fomena et al. [22] |
| M. latesi | Lates niloticus | gall bladder | Chad | 15.4 (15–16) | 8.3 (8–9) | = | 3.3 (3–3.5) | 3.3 (3–3.5) | - | ± | Kostoïngué et al. [23] |
| M. macrocapsulare | Scardinius erythrophthalmus Grunnian Aplodinotus | gall bladder | Europe America | 10–12 | 6 | = | 3–4 | 3–4 | - | ± | Auerbach [24] |
| M. macrocheili | Catostomus macrocheilus | gall bladder | North America | 11.7 ( 10.0–14.4) | 6.6 (5.5–8.0) | = | 4.0 (3.0–5.5) | 3.5 (2–4.5) | 4–6 | 9–11 | Mitchell [26] |
| M. moxostomatis | Myxostoma sp. | gall bladder | USA | 8.5–10.5 | 5–6 | = | 3 | 3 | - | 10 | Kudo [25] |
| M. nyongensis | Barbus jae, B. aspilus, B. guirali, B. martorelli | gall bladder | Cameroon | 12.3 (10.8–14.4) | 6.4 (4.7–9.4) | = | 3.2 (2.0–4.5) | 3.2 (2.0–4.5) | - | 12 | Fomena et al. [22] |
Averages of the parameters measured are followed by minimal and maximal values in brackets.
LS: length of the spore; WS: width of the spore; PC: relative length of the polar capsules (=: equal); LPC: length of the polar capsules; WPC: width of the polar capsules; FC: number of polar filament coils; StV: Number of striations on shell valves;–: absent or not reported in the species description; ±: shell valves striations are present, but the number was not reported in the species description.
Table 1: Comparative description of Myxidium tetraodoni sp. nov. with morphologically similar species (measurements in micrometre).
Host: Tetraodon lineatus Linnaeus, 1758 (Tetraodontidae).
Location: Maga (in the reservoir lake) in Cameroon (Central Africa).
Location in the host: Urinary bladder
Prevalence: 76.9% (20 individuals fish parasitized out of 26 examined).
Myxidium anisocapsularis sp. nov.
Vegetative form: Not observed; spores, sometimes abundant, were found free in the bile.
Spores: Of medium size (15.2 μm long on average), they are spindleshaped, elongated (2.5 times longer than wide), with a turgid medial part and acuminate extremities (Figures 4 and 5). Each valve carries 8 to 11 longitudinal striations (Figures 5 and 13). The suture line is straight. The polar capsules are piriform and quite unequal (Figures 6 and 13). The larger polar capsule contains 5 to 6 loops of the filament and the smaller 4 to 5 (Figure 12). A finely granular sporoplasm is located in the extra-capsular space.
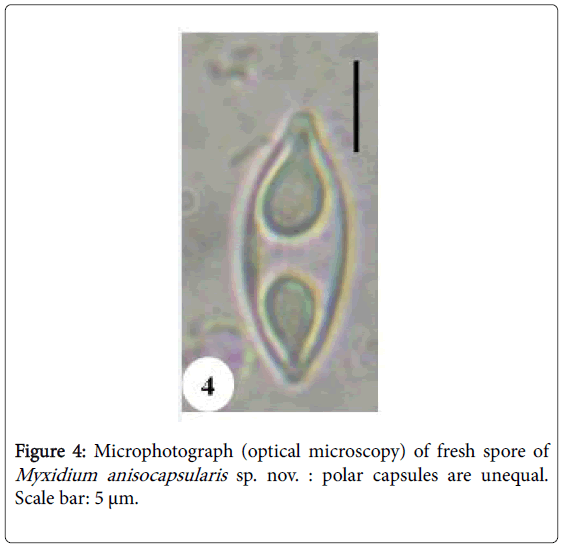
Figure 4: Microphotograph (optical microscopy) of fresh spore of Myxidium anisocapsularis sp. nov. : polar capsules are unequal. Scale bar: 5 μm.
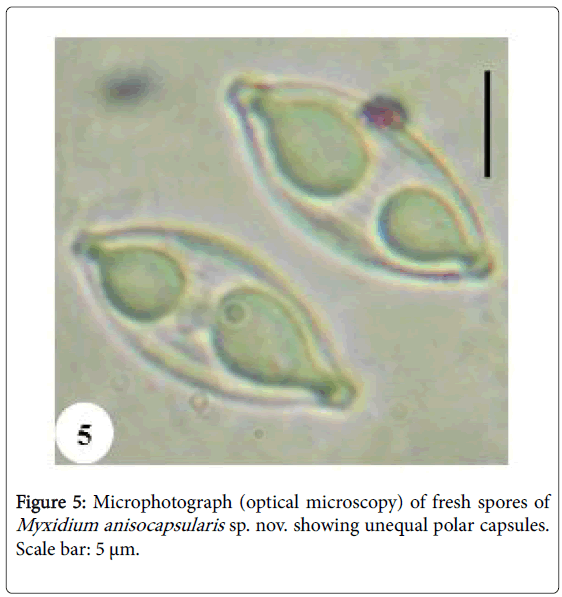
Figure 5: Microphotograph (optical microscopy) of fresh spores of Myxidium anisocapsularis sp. nov. showing unequal polar capsules. Scale bar: 5 μm.
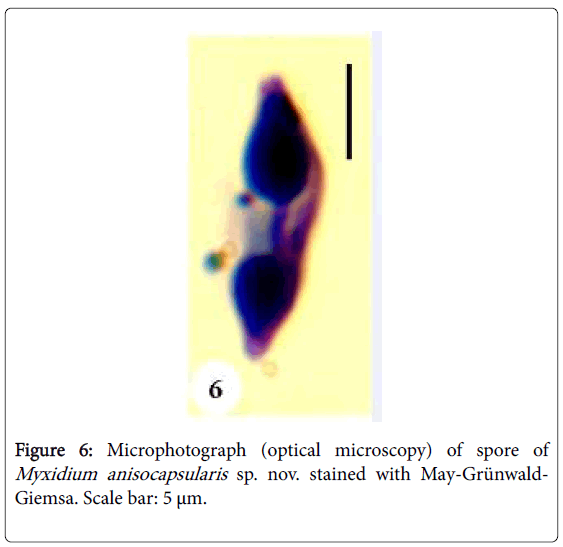
Figure 6: Microphotograph (optical microscopy) of spore of Myxidium anisocapsularis sp. nov. stained with May-Grünwald- Giemsa. Scale bar: 5 μm.
Measurements of the spore: (Table 2).
| Myxidium species | Host | Site of infection | Country | LS | WS | PC | LPC | WPC | FC | StV | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| M. anisocapsularis | Distichodus engycephalus | gall bladder | Cameroon | 15.2 (14.0–16.2) | 6.0 (5.0–6.7) | ≠ | 6 (5.0–6.5)* | 3.3 (3.0–3.8)* | 5–6* | 8–11 | Present study |
| 4.7 (4.0–5.5)** | 3. (2.3–3.3)** | 4–5** | |||||||||
| M. aydai | Caesio suevicus | gall bladder | Egypt | 23.0 (22.0–24.0) | 5.6 (5.0–6.0) | = | 7.2 (7.0–8.0) | 3.4 (3.0 –4.0) | 8–9 | - | Abdel-Baki [29] |
| M. birgii | Aphyosemiom bivittatum | gall bladder | Cameroon | 20.0 (17.7–22.5) | 8.9 (7–11) | = | 8.1 (7.1–9.1) | 3.7 (2.5–4.8) | 7–9 | 8–14 | Fomena et al. [22] |
| M. camerounensis | Neolebias ansorgei | gall bladder | Cameroon | 22 (19.2–25.5) | 6.4 (5–8.4) | = | 8.9 (7–10.9) | 3.4 (2.8 –3.9) | 7–9 | 6–10 | Fomena et al. [22] |
| M. distichodi | Distichodus engycephalus | gall bladder | Chad | 16.3 (16–17) | 6.5 (6–7) | = | 4.9 (4.5–5.5) | 3.2 (3–3.5) | - | ± | Kostoïngué et al. [23] |
| M. mendehi | Barbus guirali, B. martorelli | Kidneys | Cameroon | 9.9 (7.8–13.2) | 4.1 (3.1–4.9) | = | 3.4 (2.7–4.5) | 2.3 (1.8 –3.1) | 5–7 | 5 | Fomena et al. [28] |
| M. parachannae | Parachanna obscura | gall bladder | Benin | 23.4 (21–25) | 4.3 (3–5) | = | 5.8 (5–7) | 2.2 (1.5–3) | - | - | Sakiti [27] |
| M. petrocephali | Petrocephalus simus | gall bladder | Cameroon | 24.1 (21.5–27) | 8 (6.5–9.8) | = | 10.3 (8.5 –11.6) | 4.3 (3.8–5) | 13 | 12 | Fomena et al. [22] |
Averages of the parameters measured are followed by minimal and maximal values in brackets.
LS: length of the spore; WS: width of the spore; PC: relative length of the polar capsules (=: equal; ≠: unequal); LPC: length of the polar capsules; WPC: width of the polar capsules; FC: number of polar filament coils; StV: Number of striations on shell valves; –: absent or not reported in the species description; ±: shell valves striations are present, but the number was not reported in the species description;
*:Referring to the largest polar capsule;
**:Referring to the smallest polar capsules.
Table 2: Comparative description of Myxidium anisocapsularis sp. nov. with morphologically similar species (measurements in micrometre).
Host: Distichodus engycephalus Günther, 1964 (Distichodontidae).
Location: Maga (in the reservoir lake) in Cameroon (Central Africa).
Location in the host: Gall bladder.
Prevalence: 15.38% (04 parasitized fish out of 26 examined).
Myxobolus magai sp. nov.
Vegetative form: This Myxosporidia forms whitish and ovoid cysts, located between the secondary gill lamellae. These plasmodia are polysporous and measure 172–300 × 116 μm-200 μm. one can find 10 to 132 cysts per individual fish host (Figures 2-11).
Spores: of small size (10.6 μm long on average), mature spores are ovoid with the anterior end larger and hillocky, the posterior end being narrow (Figures 7, 8, 14 and 15). The wider part of the spore is observed at the mid length of the polar capsules. Polar capsules are ovoid and symmetrical (Figures 7 and 8). They occupy the 1/3 of the spore cavity, converge and open at the anterior end of the spore (Figure 8). Each of them contains 4 to 5 coils of polar filament (Figure 14). The iodinophilous vacuole is found in a developped sporoplasm (Figures 7 and 15).
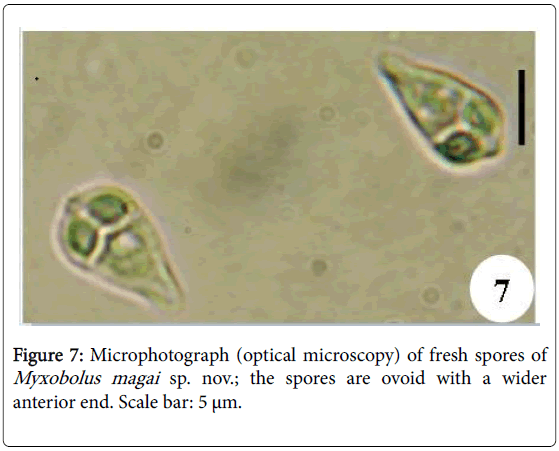
Figure 7: Microphotograph (optical microscopy) of fresh spores of Myxobolus magai sp. nov.; the spores are ovoid with a wider anterior end. Scale bar: 5 μm.
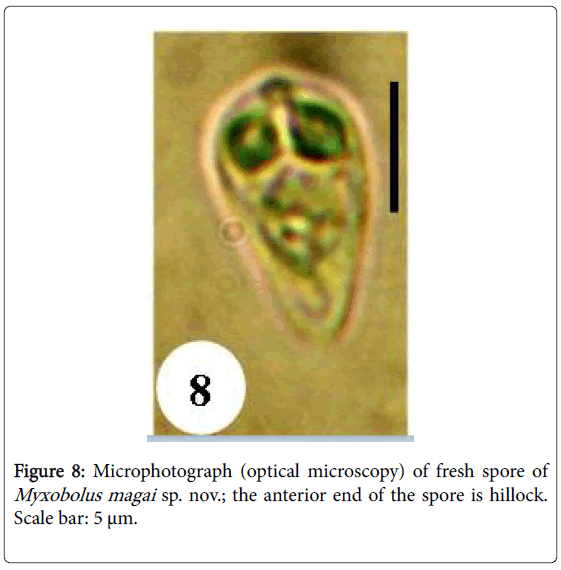
Figure 8: Microphotograph (optical microscopy) of fresh spore of Myxobolus magai sp. nov.; the anterior end of the spore is hillock. Scale bar: 5 μm.
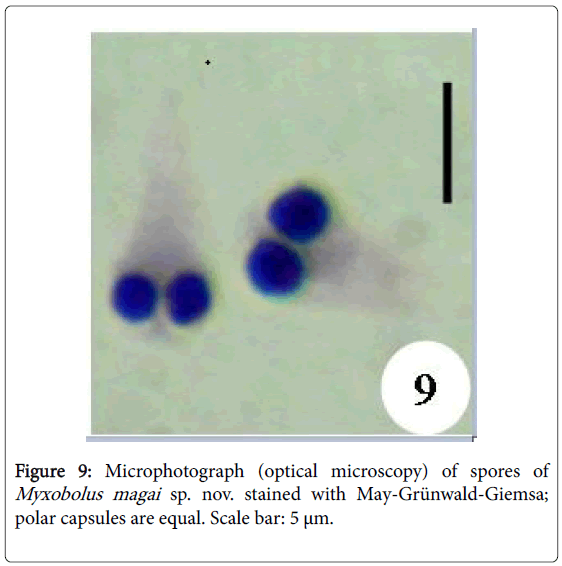
Figure 9: Microphotograph (optical microscopy) of spores of Myxobolus magai sp. nov. stained with May-Grünwald-Giemsa; polar capsules are equal. Scale bar: 5 μm.
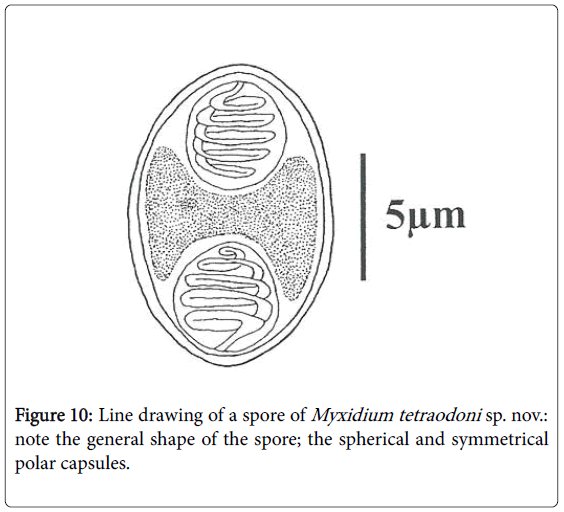
Figure 10: Line drawing of a spore of Myxidium tetraodoni sp. nov.: note the general shape of the spore; the spherical and symmetrical polar capsules.
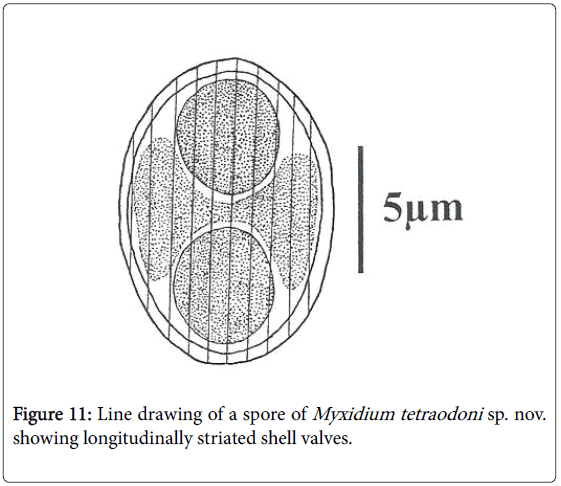
Figure 11: Line drawing of a spore of Myxidium tetraodoni sp. nov. showing longitudinally striated shell valves.
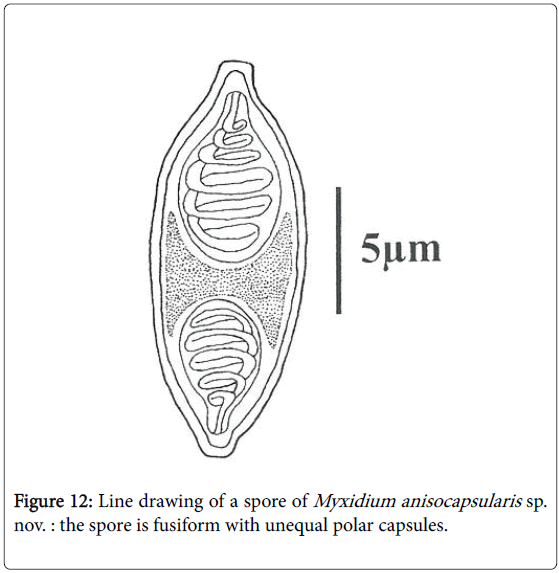
Figure 12: Line drawing of a spore of Myxidium anisocapsularis sp. nov. : the spore is fusiform with unequal polar capsules.
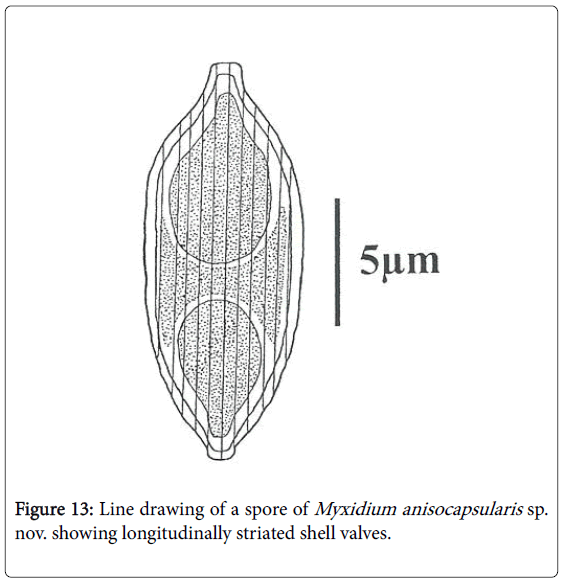
Figure 13: Line drawing of a spore of Myxidium anisocapsularis sp. nov. showing longitudinally striated shell valves.
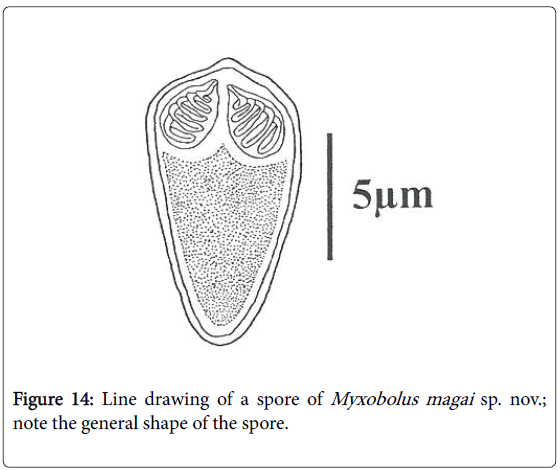
Figure 14: Line drawing of a spore of Myxobolus magai sp. nov.; note the general shape of the spore.
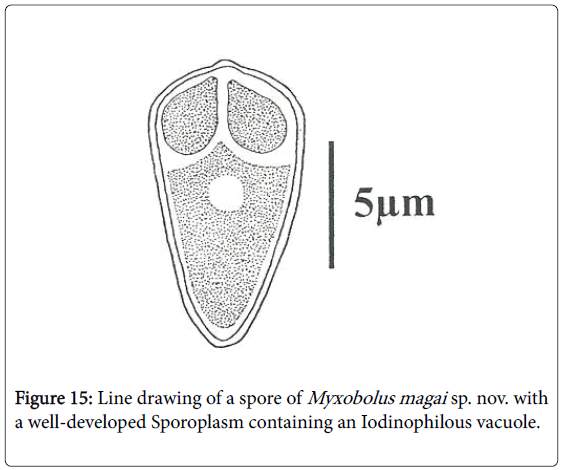
Figure 15: Line drawing of a spore of Myxobolus magai sp. nov. with a well-developed Sporoplasm containing an Iodinophilous vacuole.
Measurements of the spore: (Table 3).
| species | Host | Site of infection | Country | LS | WS | PC | LPC | WPC | FC | IP | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Myxobolus magai | Labeo batesii | Gills | Cameroon | 10.6 (9.0–12.0) | 6.3 (5.5–7.0) | = | 2.8 (2.4–3.4) | 2.3 (2.0–3.0) | 4–5 | A | Present study |
| Myxobolus cichlidarum | Oreochromis niloticus ; Sarotherodon galilaeus | Gills, fins, eye, kidneys, spleen, liver | chad | 15.0 (13.5–16) | 9.6 (8.5–10.8) | = | 5.8 (5.2–6.5) | 3,2 (2,8–3,5) | 4–5 | P | Abakar-ousman [31] |
| Myxobolus eirasi | Cirrhina mrigala | caudal fin | India | 8.4–8.8 (8.6) | 6.5–6.9 (6.7 | = | 3.1–3.3 (3.2) | 1.4–1.7 (1.5) | 3–4 | A | Kaur et al. [32] |
| Myxobolus galilaeus | Sarotherodon galilaeus | Kidneys, spleen | Israel | 11.9 (10.3–13.1) | 9.1 (7.9–10.0) | = | 3.5 (3.1–4.0) | 2.8 (2.3–3.1) | 4–5 | A | Landsberg [30] |
| Myxobolus gandiolensis | Tilapia guineensis | kidneys | Senegal | 11.3 (10–12) | 10.3 (9–12) | = | 3.8 (3-5) | 3.8 (3–5) | - | A | Fall et al. [33] |
| Myxobolus nounensis | Sarotherodon galilaeus, Tilapia mariae | Spleen, kidneys | Cameroon | 14.3 (13–15) | 12.8 (11.5–14) | = | 5.8 (5–6.5) | 4.5 (4–5) | 4–5 | P | Fomena et al. [35] |
| Myxobolus sarigi | Oreochromis aureus x O. niloticus | Spleen | Israel | 11.3 (9.9 –13.1) | 8.4 (7.9–9.6) | = | 4.5 (4.1–5.2) | 3.2 (2.9–4.0) | 4–5 | A | Landsberg [30] |
| Myxobolus yogendrai | Cirrhina mrigala | under scales | India | 9–9.5 | 7.2 | = | 2.8–3.6 | 2.8–3.6 | - | A | Landsberg et al. [34] |
Averages of the parameters measured are followed by minimal and maximal values in brackets.
LS: length of the spore; WS: width of the spore; PC: relative length of the polar capsules (=: equal); LPC: length of the polar capsules; WPC: width of the polar capsules; FC, number of polar filament coils (-: not reported in the species description); IP: intercapsular process (A: absent; P: present).
Table 3: Comparative description of Myxobolus magai sp. nov. with morphologically similar species (measurements in micrometre).
Host: Labeo batesii Boulenger, 1911 (Cyprinidae).
Location: Maga (in the reservoir) in Cameroon (Central Africa).
Location in the host: gills.
Prevalence: 28.5% (04 parasitized fish out of 14 examined).
Myxidium tetraodoni sp. nov.
By the spore shape, this myxosporidia come close to some species previously described in Africa and other continents. Myxidium brienomyri Fomena et al. [22], develops large trophozoites in the gall bladder of Brienomyrus brachyistus (Mormyridae) in Cameroon. Spores of this species are longer (13.7 μm on average) and less wide (6.47 μm on average); although spherical, the polar capsules are more developed (4.2 μm diameter on average).
Myxidium nyongensis Fomena et al. [22], develop circular and large trophozoites in the gall bladder of Barbus jae , B. aspilus, B. guirali and B. martorelli (Cyprinidae) in Cameroon. Its spores are ellipsoidal with rounded ends but longer (12.38 μm on average) and less wide (6.47 μm on average) compared to those of the species in description.
The spores of Myxidium latesi Kostoïngué , Faye et al. [23] (host: Lates niloticus in Chad) are oval, with pointed ends. They are considerably more developed (15.44 μm long × 8.33 μm wide) compared to those of the present species.
Myxidium macrocapsulare Auerbach [24] (host: Scardinius erythrophthalmus in Europe and Grunnian Aplodinotus in America) forms free spores in the gall bladder with slightly curved ends pointed in the opposite directions. These spores are narrower (5.3 μm-6.8 μm) with polar capsules rather ovoid (4.6 μm × 3.8 μm on average).
The shape of spores of Myxidium moxostomatis Kudo [25], recalls that of the species we are describing; however, this parasite of the gall bladder of Myxostoma sould. in America forms much smaller spores (8.5 μm-10.5 μm × 5-6 μm).
The spores of Myxidium macrocheili Mitchell [26] (host: Catostomus macrocheilus in North America) have comparable length with those of our parasite (11.7 μm long on average); however, they are less wide (6.6 μm on average) with ovoid polar capsules which measure 4 × 3.5 μm on average.
The parasite of Tetraodon lineatus , which differ from other species by numerous characteristics, is probably new. We propose the name Myxidium tetraodoni sp. nov. refering to the generic name of the fish host. This is the first time a Myxosporidia is described in Tetraodon lineatus .
Myxidium anisocapsularis sp. nov.
The only Myxidium species so far described in African Distichodontidae is Myxidium distichodi Kostoïngué [23], a parasite of the gall bladder of Distichodus engycephalus in Chad. The spores dimensions of M. distichodi (16.3 (16-17) μm × 6.5 (6-7) μm) are comparable to those of the species being described but their polar capsules are equal (constant character).
The general form of spores of the species in description recalls that of Myxidium parachannae Sakiti [27], parasite of the gall bladder of Parachanna obscura in Benin. However, the spores of M. parachannae are much longer (21 μm-25 μm) and less wider (3 μm-5 μm).
Myxidium birgii Fomena et al. [22], found in the gall bladder of Aphyosemiom bivittatum (Cyprinodontidae) in Cameroon form fusiform spores, with striated valves and a bulging medial part, but longer (17.7 μm-22.5 μm long). The spores shape of Myxidium mendehi Fomena et al. [28], recalls that of the species we are describing; however, this parasite of the kidneys of Barbus guirali and B. martorelli (Cyprinidae) form smaller spores (9.9 μm × 4.1 μm in average).
The spores of Myxidium camerounensis Fomena et al. [22], although fusiform with a bulging medial part, are longer (22.04 μm on average), with equal polar capsules containing 7 to 9 loops of polar filament.
The spores of Myxidium petrocephali Fomena et al. [22] (host: Petrocephalus simus in Cameroon) are slightly arched, more developed (24.14 μm × 8.05 μm on average) with symmetrical and larger polar capsules (10.3 μm × 4.3 μm on average).
Myxidium aydai Abdel-Baki [29], parasite of Caesio suevicus in Egypt produces spores measuring 23 μm long on average with symmetrical polar capsules measuring 7.2 μm on average.
Out of all Myxosporidian species of the genus Myxidium described so far in the world, no one form spores with unequal polar capsules. We believe that the parasite of Distichodus engycephalus is new and propose the name Myxidium anisocapsularis sp. nov., refering to the asymmetry observed on its polar capsules.
Myxobolus magai sp. nov.
By the spore’s general shape, the present parasite resemble some species of Myxosporidia previously described.
Myxobolus galilaeus Landsberg [30], forms diffuse spores in melano-macrophage centers of the kidneys and spleen of Sarotherodon galilaeus in Israel. These spores are ovoid with the anterior end flattened. They are larger (11.9 μm × 9.1 μm on average) and 3 to 12 folds are found on the suture line. Myxobolus sarigi Landsberg [30], a systemic parasite of Cichlidae in Israel, forms spores with the anterior end wider. These spores are wider (8.4 μm on average) with larger polar capsules (4.5 μm × 3.2 μm on average).
Myxobolus cichlidarum Abakar-ousman [31], parasitizes fins, gills, eyes, spleen, kidneys and liver of Oreochromis niloticus and Sarotherodon galilaeus in Chad. Although ovoid with a wider anterior end, the spores of this species, differ from those of our species by the following characteristics: larger size (15 μm × 9.6 μm on average), more developed polar capsules (5.8 μm × 3.2 μm on average), presence of an intercapsular appendix.
The spores of Myxobolus eirasi Kaur et al. [32] (parasite of Cirrhina mrigala in India), are morphologically comparable to those of our parasite, but shorter (8.4 μm-8.8 μm) with smaller polar capsules (1.4 μm-1.75 μm).
Fall et al. [33] described Myxobolus gandiolensis in the kidneys of Tilapia guineensis in Senegal. This Myxosporidia differs from the species in description by its wider spores (10.3 (9-12) μm) and its spherical polar capsules.
In India, Myxobolus yogendrai (Triparthi, 1952) Landsberg et al. [34], parasitize Cirrhina mrigala ; its spore, although ovoid with a wider anterior end, possess an intercapsular triangle and four valvular folds.
Myxobolus nounensis Fomena et al. [35], is a parasite of the spleen and kidneys of Sarotherodon galilaeus and Tilapia mariae (Cichlidae) in the Noun River in Cameroon. Spores of this species are more developed (14.3 μm × 12.8 μm on average), same as its polar capsules (5.8 μm × 4.5 μm on average) and a well-developed intercapsular appendix is present.
These differences lead to think that the parasite of Labeo batesii is a new species. We propose the name Myxobolus magai sp. nov., refering to the locality of Maga where the fish host were captured.
Although Africa has one of the most diverse ichthyological fauna, and the genus Myxidium is the second major group of Myxosporidia, the description of Myxidium tetraodoni sp. nov. and M. anisocapsularis sp. nov. Only brings up to 18 the number of species of the genus Myxidium described in freshwater fishes of this continent, and to 11 the number of species of the genus described in freshwater fishes of Cameroon. M. tetraodoni is the first Myxosporidia described in fishes of the family Tetraodontidae and M. anisocapsularis is the only species of the genus Myxidium hitherto found with unequal polar capsules. Species of the genus Myxidium are generally coelozoic and rarely histozoic in fish host. The two new species of Myxidium described in the present work are coelozoic. These coelozoic Myxosporidia are generally less pathogenic than histozoic species, their pathogenic action seems to be less evident apart from the occlusion of bile and urinary ducts. In Africa, the species of Myxidium that are identified so far affect hosts belonging to 12 families. In decreasing importance order, these host families can be classified as: Cyprinidae (5 species); Claroteidae (2); Distichodontidae (2); Mochokidae (2); Mormyridae (2); Anabantidae (1); Aplocheilidae (1); Channidae (1); Cichlidae (1); Clariidae (1); Latidae (1); Tetraodontidae (1). Myxosporidia of the genus Myxobolus are generally known as histozoic parasites in freshwater fish. They form cysts of varying size in various organs in their host (gills, fins, stomach wall, intestinal wall, skin, heart, liver, operculum, eyes ...). Eiras et al. [8,9] estimated at about 905 species the number of Myxobolus described in fishes around the world. 33.25% of these species affect the gills in their hosts. The description of M. magai sp. nov. in the gills of Labeo batesii confirms the preference of this organ by the Myxosporidia of the genus Myxobolus.
Animals used followed a protocol approved and authorized by Institutional Animal Care and Use Committee at Animals Biology and Physiology Department, Faculty of Science, University of Yaounde I, Cameroon.
The authors are thankful to the Faculty of Science, University of Yaounde I, Cameroon, for providing all the facilities to complete this work.
The authors declared: There is no conflict of interest.