Journal of Glycobiology
Open Access
ISSN: 2168-958X
![]() +44 1478 350008
+44 1478 350008
ISSN: 2168-958X
![]() +44 1478 350008
+44 1478 350008
Research Article - (2012) Volume 1, Issue 1
Background: Undergalactosylated IgA1 has been found to be increased in IgA nephropathy (IgAN) by an ELISA assay using Helix aspersa agglutinin (HAA) that recognizes N-acetylgalactosamine (GalNAc). In this study, we developed a polyclonal antibody (anti-sHGP antibody) against a synthetic IgA1 hinge peptide with five GalNAc residues.
Methods: The specificity of the anti-sHGP antibody was evaluated through the incremental treatment of IgA with corresponding glycosidases. Then, the susceptibility of the IgA to anti-sHGP antibody was compared among IgAN patients (n=39), patients with other forms of kidney diseases (OKD, n= 36) and healthy controls (n=37), using ELISA assay. The association of the binding abilities between anti-sHGP antibody and HAA were evaluated blindly using same 85 sera.
Results: The binding ability of the anti-sHGP antibody was increased relative to the incremental treatments of neuraminidase (desialo-IgA), galactosidase (desialo/degalacto IgA). The binding levels of anti-sHGP antibody against serum IgA were significantly higher in IgAN patients compared to both healthy controls (P=0.008) and those with OKD (P=0.049). The binding levels of anti-sHGP antibody were closely related to those of HAA ELISA in the same patient sera (RR2=0.5964).
Conclusions: It was certified that the anti-sHGP antibody recognized GalNAc residues in the hinge peptide of human IgA1 as well as HAA. The increased antigenicity of IgA against the antibody in IgAN suggested that a serum IgA1 exposing GalNAc residue was increased in IgAN. It would be necessary to identify the precise structure of O-glycans specific to IgAN for developing a more specific antibody.
Keywords: IgA nephropathy; IgA1 hinge region; N-acetylgalactosamine; Galactose; Anti-IgA1 hinge antibody
IgA: Immunoglobulin A; GalNAc: N-acetylgalactosamine; sHGP: synthetic Hinge Glycopeptide; OKD: Other forms of Kidney Diseases; HAA: Helix Aspersa Agglutinin; MALDI-TOFMS: Matrix-Assisted Laser Desorption Ionization Time Of Flight Mass Spectrometry; KLH: Keyhole Limpet Hemocyanin
There is increasing evidence for the involvement of aberrantly glycosylated IgA1 in the pathogenesis of IgAN [1-8]. A variant with terminal GalNAc or sialylated GalNAc is rare in normal serum IgA1, but it has been suggested that the presence of the truncated O-glycan with an exposed N-acetylgalactosamine (GalNAc) residue is more common in the IgA1 of IgAN patients. In previous studies, O-glycans in the hinge region of IgA1 were evaluated by ELISA assay using Helix asperse agglutinin (HAA), a lectin specifically binding to GalNAc residues [9,10]. The assay suggested the increased binding of HAA to serum IgA1 in IgAN with high specificity and sensitivity. However, it is known that the affinity of lectin is much less than that of antibody. Therefore, in this study, we tried to develop a polyclonal antibody specifically binding to undergalactosylated IgA1, expecting that the antibody would be available as a clinical tool for the diagnosis of IgAN.
Patients and test sera
Thirty-nine patients with biopsy-proven IgAN were examined. The serum samples were obtained at the time of renal biopsy. An additional 36 patients with other forms of kidney diseases (OKD) were also examined (Table 1). The absence of glomerular IgA deposits were confirmed in all OKD patients by renal biopsy. The itemization of OKD and patient number of each disease were described in Table 1. Thirty-seven healthy control subjects were selected from healthy individuals matched for gender and age to the IgAN patients. A 19 mer synthetic peptide with five GalNAc residues at 4, 7, 9, 11 and 15 [VPST (GalNAc) PPT (GalNAc) PS (GalNAc) PS (GalNAc) TPPT (GalNAc) PSPS-NH2] was purchased from Peptide Institute, Inc. (Osaka, Japan). The binding sites were determined according to the report of Mattu et al. [11] in that GalNAc residues frequently bind to these sites in human IgA1 hinge region. The purity and molecular weight were confirmed by HPLC and MALDI-TOFMS. For immunization, sHGP was conjugated with Keyhole Limpet Hemocyanin (KLH, Sigma). The sHGP–KLH conjugate was subcutaneously injected with Freund’s complete adjuvant to domestic mongrel white rabbits (n=2). The immunization was performed four times in every two weeks. Then, the antisera were isolated following exsanguination. The antisera were certified to bind to sHGP at the dose – response manner. IgG fraction was then obtained by ion exchange chromatography.
| Diseases | Number of patients |
|---|---|
| Minimal change nephrotic syndrome Membranous nephropathy Anti-neutrophil cytoplasmic autoantibody associated nephritis Sarcoidosis Lupus nephritis Acute glomerulonephritis Membranoproliferative glomerulonephritis Rheumatoid arthritis Amyloidosis Focal segmental glomerulosclerosis Chronic nephritis Interstitial nephritis Thin basement membrane disease Choresterol crystal embolism Obese nephritis Diabetic nephropathy Nephrosclerosis |
6 4 4 3 3 2 2 2 2 1 1 1 1 1 1 1 1 |
| Total | 36 |
Table 1: Itemizing of other kidney diseases (OKD).
The serum IgA1 was isolated from the sera of healthy individuals using jacalin affinity chromatography according to our previous study [7]. The purified serum IgA1 samples were digested with corresponding glycosidases according to our previous study in an incremental manner [4]. Firstly, The IgA1 (called native IgA1) was digested with neuraminidase (NA) from Streptococcus 6646K (1U/ml; Seikagaku Co. Osaka, Japan) (Desialo IgA1). Some of the asialo IgA1 were then treated with β-galactosidase (GA) from bovine testis (1.0 U/ml; Sigma Chemical Co., St. Lous, MO) (Desialo/Degalacto IgA1). The naked (deglycosylated) IgA1 was then isolated from Desialo/Degalacto IgA1 using α-N acetylgalacosaminidase from Acremonium sp (Seikagaku Co.). The specificity of the anti sHGP antibody against native, desialo-, desialo/degalacto- and naked-IgA1 molecules was compared using ELISA described below.
We observed the dose–response curve of IgA bound to the antisHGP antibody using serially diluted (1/100–1/10000) human serum samples, to ascertain the specificity of the antibody. From the dose – response curve, 500 diluted sera were regarded as an optimal dilution to compare the binding among IgAN, OKD and HC groups. A 50 μl aliquot of 0.05 M bicarbonate buffer (pH 9.6), containing the F(ab)2 fragment of goat anti-human serum IgA (Jackson Immuno Research Labs, West Grove, PA, USA, 2.6 μg/ml), was added into the wells of a polystyrene microtiter plate (Corning, NY, USA) and incubated overnight at 4°C. The plates were washed with 0.01M PBS–0.05% Tween-20 (PBST) three times for each step. Coated plates were blocked with 1% bovine serum albumin (BSA; Sigma Chemical Company, St Louis, MO, USA) in PBST for 1 h at room temperature (RT). A 50 μl/well aliquot of the 500 diluted sera in blocking solution was added to the coated wells, and incubated overnight at RT. For removing the terminal neuraminic acid, the samples were incubated with 100 μl of 20 mU/ml neuraminidase (Roche Diagnostic Corp., Indianapolis, IN, USA) in 0.01M acetate buffer (pH 5) for 3 h at 37°C. After washing, 50 μl of anti-sHGP antibody (7.6 μg/ml) was added to each well. Following a further 2h of incubation at 37°C, and three times washing, 50 μl of peroxidase conjugated goat anti rabbit IgG (80 ng/ml; Sigma) was added. After one hour incubation at RT and washing, the wells were then developed with 50 μl of O-phenylenediamine (0.5 mg/ml)–H2O2 (0.1%, Pierce) in 0.1 M citrate phosphate buffer. The colour reaction was stopped with 50 μl of 1 M sulphuric acid and absorbance at 490 nm was measured.
Each sample was tested in triplicate and the median values were regarded as the absorbance levels of each sample. Among the 112 total serum samples, HAA-IgA binding assay had been performed in the 85 serum samples in the previous study [10]. The results of the two assays were obtained blindly and the association of the results was evaluated. The Mann-Whitney U-test with Bonferroni correction was used for comparing the binding levels among patients with IgAN, OKD, and healthy controls. It was regarded as statistically significant if the p value was less than 0.05.
The binding ability of the anti-sHGP antibody was increased relative to the incremental treatment of neuraminidase (desialo-IgA), galactosidase (desialo/degalacto IgA). The anti-sHGP antibody also bound significantly to the naked IgA (Figure 1). However, the binding was slightly decreased compared to that of desialo/degalacto IgA. The levels of antibody–IgA binding were significantly higher in the IgAN group compared to OKD (P = 0.049) and to HC (P = 0.008) groups (Figure 2). There was no significant difference between HC and OKD (p=1.839). The binding levels of the anti-sHGP antibody were closely correlated to those of HAA ELISA in the same patient sera (RR2=0.5964, Figure 3).
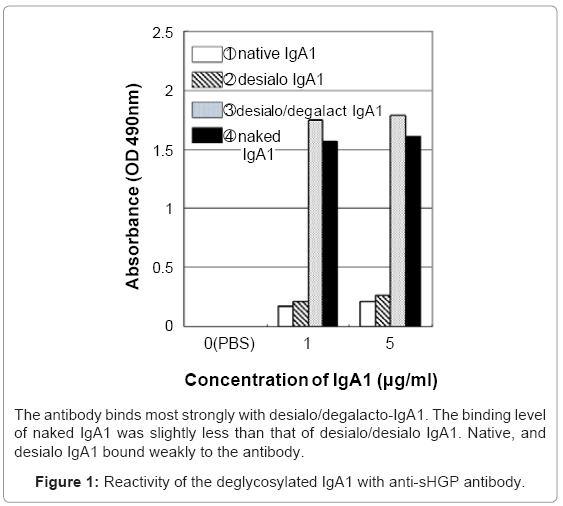
Figure 1: Reactivity of the deglycosylated IgA1 with anti-sHGP antibody.
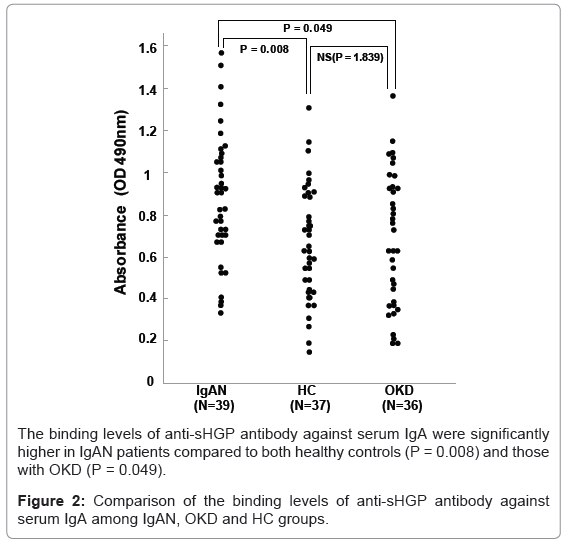
Figure 2: Comparison of the binding levels of anti-sHGP antibody against serum IgA among IgAN, OKD and HC groups.
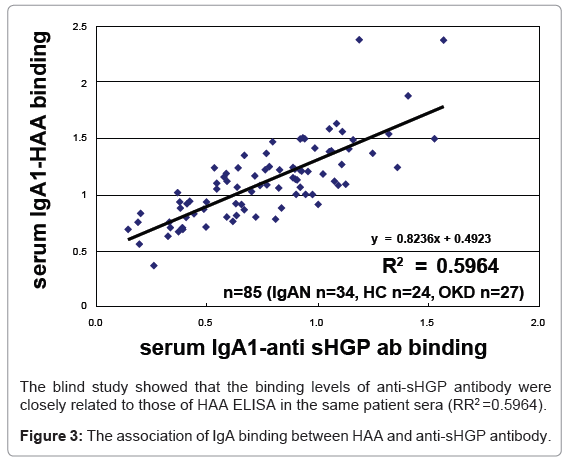
Figure 3: The association of IgA binding between HAA and anti-sHGP antibody.
Human IgA1 molecule has a unique hinge structure due to its mucin-like O-linked oligosaccharides [11]. The core peptide consists of a proline-, serine, and threonine-rich amino acid sequence in which serine and threonine residues are able to bind to O-glycans consisting of neuraminic acid, galactose and N-acetylgalactosamine with microheterogeneity. In the past two decades, it has been demonstrated that there are some abnormalities in the O-glycan structure of the IgA1 hinge region in IgAN. Although the abnormalities were suggested to be underglycosylated (especially undergalactosylated), the precise structure is not yet clear. Further, although the precise analyses were performed by mass spectrometry, the analyses limited the number of samples in the comparable study. The HAA-assay seemed to overcome this problem, clarifying the significant increase of exposed GalNAc residues in IgAN. In other words, IgAN patients had less Gal residues in IgA1 hinge region. However, the relatively weak binding ability of lectins results in the assay having rather poor reproducibility for clinical application.
In this study, we developed a polyclonal antibody against synthesized hinge glycopeptide which was regarded as a model of undergalactosylated IgA1 hinge. The binding sites of GalNAc to synthesized hinge peptide were determined according to the report of Mattu et al. in that GalNAc residues frequently bind to these sites in human IgA1 hinge region. It was certified that the polyclonal antibody produced by the immunization of the artificially synthesized glycopeptide definitely recognized the hinge peptide with five GalNAc residues in human IgA1 as well as HAA [11]. The increased antigenicity of IgA against the antibody in IgA nephropathy suggested that IgAN patients have serum underglycosylated IgA1 exposing GalNAc residues and/or hinge core peptide. However, the antibody did not have sufficient specificity for clinical application in the diagnosis of IgAN. It would be necessary to clarify the precise structure of O-glycans specific in IgAN to develop a more specific antibody.