Anthropology
Open Access
ISSN: 2332-0915
ISSN: 2332-0915
Perspective - (2018) Volume 6, Issue 1
The basis for the late stages of brain growth in our immediate human ancestors is unknown, but generally attributed to the development of language associated with increasingly complex social settings. We present arguments for an additional or alternative potential contributor to endocranial expansion. As ancestral humans emerged from tropical forests into open savannahs, loss of their body hair facilitated the dissipation of body heat. But their newly-exposed cutaneous surfaces would have been bombarded by exogenous stimuli that could have stimulated an increase in brain size required to centrally process a plethora of new sensory information arriving from the periphery. Two lines of evidence support this hypothesis. First, the common embryologic origin of epidermis and central nervous system (CNS) from the primitive neuroectoderm, endowed the skin with a broad suite of neuro mechanisms that mirrors the CNS, including outward-facing ‘physical’ receptors that monitor changes in visual, acoustic and olfactory stimuli, as well as many neuromediators that impact memory, mood and wakefulness in the CNS. Second, two groups of marine organisms, cephalopods and electric fish, exhibit large brain: body mass ratios associated with heightened cutaneous sensory capabilities. Human nakedness also stimulated new psychosocial and stress responses, mediated by cutaneous oxytocin and glucocorticoids, respectively. Together, these insights suggest that the onset of hairlessness could have stimulated an increment of endocranial expansion, shortly before the emergence of modern humans.
Keywords: Brain; Epidermis; Evolution; Hairlessness; Neuroreceptors; Neurosensors; Skin; TRPVs
CNS: Central Nervous System; GABA: γ-Amino- Butyric Acid; mya: Million Years Ago; O2: Oxygen; TRPV: Transient Receptor Potential Vanilloid
While australopithecines exhibited endocranial volumes that were comparable to their great ape ancestors [1], brain volumes began to expand ≈3 million years ago (mya) with the transition from a predominantly vegetarian, arboreal to an increasingly omnivorous, ambulatory lifestyle [2]. Because these new sources of protein were widely dispersed in tropical forests, these ancestral humans established centralized campsites where groups of hunters developed new weapons and food processing tools, as they ventured further afield. Together with the increased stress of group living, which placed a premium on intragroup cooperation, these forces together selected for a further expansion in brain volume [3]. The next increment of brain growth began 1-1.5 mya as ancestral hominins (H. erectus) aggregated at campsites, centered around fires [4,5]. Under such crowded circumstances, interactions required the acquisition of cooperative social skills, reasoning, and perhaps the development of proto-languages [6,7], but competition and conflict likely also flourished [8]. Together, these forces likely selected for further brain growth, made possible by cooking of dietary staples, which likely increased the bioavailability of additional nutrients needed to accomplish brain expansion [9]. Thus, the social brain hypothesis (SBH) argues that large brain size in primates represent an evolutionary response to complex social interactions and information-rich social environments.
Yet, the basis for last increments of brain growth that occurred shortly before, and perhaps concurrent with the emergence of modern humans [10,11], is speculative. As still-furred early humans ranged further and further from forest cover, they risked overheating as they exerted themselves in open savannahs. It has been suggested that bipedalism reduced the impact of solar radiation fluxes and made possible reduction of body hair (setting them apart from savannah quadrupeds) [12]. But, rapid, widespread dispersion of cutaneous eccrine glands in service to thermoregulation [13] would have been impeded by a dense furry mantle, risking excessive insolation (i.e., heat retention) under the harsh environmental conditions that prevailed in Sub-Saharan Africa ≈1-1.5 mya (e.g., [14]). Hence, eccrine gland proliferation likely selected for a subsequent, dramatic reduction in hominins’ furry pelage, necessitated by the need to dissipate heat aided by not only sweating, but also by insensible, constitutive transepidermal water loss (TEWL) [15]. TEWL provides a further evolutionary advantage by allowing a more efficient conservation of electrolytes. Thus, the high rates of TEWL likely would also have selected for new mutations that improved epidermal permeability barrier function, the dominant imperative for life in a xeric terrestrial environment [15-17].
Common Origin of Epidermis and Central Nervous System: Because newly exposed, naked hominin skin would have suddenly been bombarded by an array of new sensations, the need to integrate this new burden of sensory information could have selected for a further increment in brain growth. This hypothesis is anchored in the common origin of the central nervous system (CNS) and epidermis from the layer of embryonic neuroectoderm that coats the developing embryo. As the CNS buds off the neuroectodermal layer, this layer, which gave rise to the epidermis, remains endowed with a full repertoire of the same neuro mediators and neuroreceptors that are present in the CNS.
The Neurosensory Consequences of Hair Loss: As noted above, a central pillar of this hypothesis; i.e., that shedding of hair could have stimulated further brain enlargement, relates to the common embryological origin of the epidermis and the CNS, resulting in the epidermis’ retention of a full suite of neuro mechanisms that are traditionally associated with nervous tissues (Tables 1 and 2). While the roles of several of these neuroreceptors in the skin have been identified, many are still ‘orphan receptors,’ because their functions in epidermis remain incompletely characterized. Nonetheless, most persisted, and the ones that have been characterized clearly provide the skin with a remarkable panoply of sensory capabilities. These include not only a broad suite of ‘physical receptors’ that monitor the external environment for changes in osmotic, visual, auditory, mechanical, temperature and olfactory stimuli [18] (Table 1), but also a plethora of neuro mediators associated with ‘higher’ functions, including emotional responses, memory and seizure activity (Table 2). This plethora of cutaneous neurosensors would have remained relatively dormant, while hominin skin remained coated with a dense pelage, but the wide-spread loss of body hair would have exposed the newly hairless epidermis of our immediate hominin ancestors to a sudden onslaught of sensory stimuli [19,20]. Although other mammals, like some primates and elephants, display comparable brain:body mass ratios to ours, the number of neurons in human brains exceeds that of any other vertebrates [21]. Thus, an urgent need for central processing of this sudden sensory bombardment from the periphery could have stimulated a late increase in hominin brain size (Figure 1).
| Transient Receptor(TRPV)Family | Role | References |
|---|---|---|
| TRPV 1, 3 4 | Osmotic pressure pH (<6.6), Temperature (>35-43° c), Barrier homeostasis |
Liedtke W. 2007 Exp physiol. 92: 507-512 Denda M. 2011 Adv Exp Med Biol.704:847-860 Denda M. 2007 J Invest Dermatol 127: 654-659 |
| TRPA-1 | Temperature (<17°), pain, mechanical stress, Barrier homeostasis |
Talavera k. 2009 Nat Neurosci 12: 1293-1299 Denda M. 2010 J invest Dermatol 130: 1942-1945 |
| TRPM-8 | Temperature (<22°C), Barrier homeostasis |
Tsutsumi M. 2010 J invest Dermatol 130: 1945-1948 Denda M. 2010 Exp Derm 19(9) 791-795s |
| Visual light receptors | ||
| Rhodopsin, Opsins | Barrier homeostasis | Tsutsumi M. 2009 Exp Derm 18(6) 567-570 |
| Melatonin | Barrier homeostasis | Denda M. 2005 Dry skin and moisturizers, 2nd Edition 155-160 |
| Acoustic | ||
| High - Frequency sound (20-100Khz) |
Hypersonic effects Barrier homeostasis | Kawai N. 2001 Neuro reports 12: 3419-3423 Oohashi T. 2006 Brain Res 1073-4: 339-347 Denda M. 2005 Br J Dermatol 162: 503-507 |
| Olfactory | ||
| OR2AT4 | Wounding healing, epidermal Proliferation |
Busse D. 2014 J Invest Dermatol 134(11) 2823-2832 |
Table 1: Physical receptors at the cutaneous- environmental interface.
| Neurotransmitters | Role | Partial List of References |
|---|---|---|
| Histamine | Inflammation, Itch, Barrier homeostasis | Yamamaura K, J toxicol sci 34(4) 427-431, 2009 Ashida Y, JID 116(2) 261-265, 2001 Giustizieri ML, JACI 114(5) 1176-1182, 2004 |
| Dopamine | Barrier homeostasis +other? | Fuziwara S, JID 125(4) 783-789, 2005 |
| Serotonin | - | Denda M, 2005 Skin Moisturizer, 2nd edition 155-160 |
| Nicotinic | Antimicrobial Défense, Barrier homeostasis | Radek K, Cell host micro 27(7) 277-289, 2010 Denda M. JID 121(2) 362-367, 2003 |
| GABA | Barrier homeostasis+ keratinocyte proliferation | Denda M, JID 119(5) 1041-1047, 2002 |
| Glycerine | Barrier homeostasis+ keratinocyte proliferation | Denda M, JID 121 (2) 362-367, 2003 |
| Aldrenergic | Barrier homeostasis+ keratinocyte proliferation | Denda M, JID 121 (1) 142-148, 2003 |
| Glutamate | Barrier homeostasis+ keratinocyte proliferation | Fuziwara S, JID 120(6) 1023-1029, 2003 |
| Adenosine | Barrier homeostasis+ keratinocyte proliferation | Denda M, JID 119(5) 1034-1040, 2002 |
| Neurohormones | ||
| cannabinoids | Barrier homeostasis, Inflammation | Roelandt T, Exp Dermatol 21(9) 688-693, 2012 |
| opioids | Inflammation, Itch | Bigliardi PL, Exp Derm 18(5) 424-430, 2009 |
| CRF, ACTH, GC | Stress responses | Slominski A, Physiol Rev 80(3) 979-1020, 2000 |
Table 2: Other neuro mediators at the cutaneous-environmental interface.
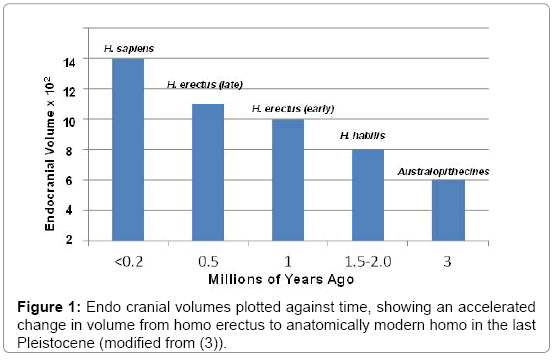
Figure 1: Endo cranial volumes plotted against time, showing an accelerated change in volume from homo erectus to anatomically modern homo in the last Pleistocene (modified from (3)).
Epidermal ‘Physical Receptors’: Transient receptor potential vanilloids (TRPV) sense changes in temperature, among other physical and chemical factors in both the epidermis [22] and CNS [23]. TRPVs are activated in response to changes in skin surface pressure, temperature, acidity and osmolarity, which in turn regulate epidermal permeability barrier homeostasis [24-26]. But visible light exposure also influences epidermal permeability barrier homeostasis [27], and certain proteins in keratinocytes, including rhodopsin, blue-opsin, red-green opsin, transducin and phosphodiesterase 6, that play crucial roles in retinal visual perception [28,29]. Although their function in skin is still unclear, exposure to strong light modifies systemic hormone levels, even in humans afflicted with congenitally impaired vision [30]. Moreover, the epidermis synthesizes melatonin, a classic light-sensitive hormone as well as abundantly expressing melatonin receptors [31]. Therefore, the skin acts as a peripheral sensor of visual signals en route to processing in the brain.
Likewise, sound, via still unidentified, cutaneous auditory receptor(s), also influences whole-body physiology and epidermal homeostasis. For example, live music (at sound frequencies >20 kHz) influences performers’ brain waves and hormone levels, but the acoustic stimuli have little impact when music of this frequency derives from compact disk sources [32,33]. Yet, when a sound barrier separates test subjects from the live music source, the impact of live music disappears (Ibid.), demonstrating a putative role for peripheral auditory processing. Pertinently, puffs of air applied to the skin surface influence human auditory perception, and even inaudible bursts of air applied to the hands or neck result in mishearing ‘b’ as ‘p’ [34]. Finally, exposure of hairless mouse skin to auditory frequencies of 10-30 kHz significantly enhances permeability barrier homeostasis [23,35]. Thus, the exposed skin of humans recognizes and processes small bursts of sound via cutaneous auditory receptors, but whether these responses require prior central processing is not known. In summary, detection of high-frequency sounds by exposed, hairless skin might have provided a further stimulus for brain growth as well as a survival advantage for ancestral, newly-hairless hominins threatened by large predators or natural disasters.
It should also be noted that keratinocytes express a plethora of (>20) olfactory receptors whose role in normal skin physiology remains largely unexplored. But activation of one of these orphan receptors, OR2AT4, by a sandalwood odorant, stimulates wound healing, while promoting proliferation and migration of human keratinocytes [36]. Once again, whether these epidermal responses require prior central processing is not yet known.
Additional Epidermal Neuromechanisms- the ‘Brain in the Skin’: Beyond the ‘physical neuroreceptors,’ human epidermis also deploys an array of additional neurotransmitters and receptors (Table 2), including dopamine, serotonin, histamine and γ-amino-butyric acid (GABA), whose activation traditionally is associated with regulating higher CNS functions, such as memory, emotion, seizure activity and wakefulness. Moreover, epidermis also elaborates (and receives) signals from a suite of neurohormones, including opioids, cannabinoids, adrenocorticoids, and oxytocin [37]. The last mediates important social cues, including the establishment of intimacy and the ability to empathize, both of which are crucial for interpersonal bonding [38,39]. Clearly, stimulation of these peripheral neuroreceptors must stimulate central processing. Finally, psychosocial studies suggest that cutaneous stimuli, facilitated by the loss of a dense pelage, influence human emotion [40]. Light physical contact, tends to increase generosity [41,42], and could have been critical for social communication as language systems developed [6,43]. Thus, naked skin not only enabled us to interact more efficiently with our physical environment, but it also plays an important role in social interactions and psychological well-being [44].
Relevant examples from other organisms
While the sudden, widespread shedding of a dense furry mantle would have uncovered newly exposed, epidermal neuroreceptors to continuous bombardment by often urgent peripheral sensations, there are other examples in nature of ‘naked skin’ associated with unusual brain size or complexity. The octopus provides pertinent evidence that a plethora of peripheral sensations can stimulate brain growth [45]. Because of its radial symmetry, the octopus must integrate sensory inputs not only from its eight, separate peripheral ‘brains,’ but also from its highly ‘sentient’ skin, and enormous, human-like eyes. Likely as a result, the octopus evolved a walnut-sized brain with 6-fold more neurons than occur in rodents [46,47]. Not only is the octopus intellect considered comparable to canines, perhaps exceeding even the most sentient of avians [48]. The combination of a peripheral and central sensory system has further endowed the octopus with a suite of ‘humanlike’ characteristics, including a capacity to learn, use tools, distinguish between individuals, playfulness that can become malevolent, and perhaps even human-like, emotional responses [49]. Cutaneous visual mechanisms, with which cephalopods are richly endowed, allow rapid camouflage responses in a wide variety of habitats [50].
An even more dramatic example of brain expansion in response to peripheral stimuli is the electric fish of the genus Gnathonemus. These fish possess electroreceptors in their epidermis that allow organisms to accurately assess the distance, size and shape of surrounding objects by transmitting an ‘electric image’ to the brain [51]. As a result, the brain:body mass ratio of these fish is higher than any known vertebrate, including humans, also accounting for 60% of bodily oxygen (O2) consumption, compared to the 2%-8% of O2 utilization in resting humans [52]. The hypertrophy of Gnathonemus’ brains in response to peripheral electroreceptors provides further support for the concept that the cutaneous boundary layer could influence brain size.
Another group of mammals with naked skin, Cetaceans, that occupy a marine environment and display hierarchical social structures, are considered ‘intelligent’ and show an astonishing similarity to humans and other primates in social, behavioral and neuroanatomical complexity [53]. Fox et al. provides a compelling view of the social and cultural roots of encephalization in whales and dolphins. These creatures “show overwhelming evidence for sophisticated social and prosocial behavior, including complex alliance relationships, social transfer of hunting techniques, cooperative hunting, complex vocalizations including regional group dialects, vocal mimicry and signature whistles unique to individuals, interspecific cooperation with humans, allopreening and social play.” These characteristics all rely on a highly-developed sense of touch (Ibid.). Cetacean epidermis, like human skin, displays several unique adaptations for aquatic life, including a new, complete absence of pilosebaceous units (i.e., hair) [15]. Thus, a late increment of brain growth in ancestral humans could have occurred in response to the evolution of hairlessness.
It is our hypothesis that loss of hair, with exposure of the epidermis created a new boundary layer that receives and responds to multiple external sensations and signals, as well as social interactions based on changes in appearance and emotional cues (blushing, blanching, touch, pheromones). These peripheral stimuli could have placed additional stress on the CNS, leading to further encephalization, a concept supported by several analogous examples in other vertebrate and invertebrate organisms.
Ethics approval and consent to participate: N/A
Consent for publication: N/A
Availability of data and material: Printed/identified publications
Competing Interests: None
Research reported in this publication was supported by the National Institute of Arthritis, Musculoskeletal and Skin Diseases of the National Institutes of Health under Award Number AR057752, administered by the Northern California Institute for Research and Education, with additional resources provided by a Veterans Affairs Medical Center Merit Review, San Francisco, CA.
MD: Wrote initial draft, and provided initial concept
PME: Edited manuscript, and added pertinent references
GM: Provided critical information about evolutionary aspects of hair in mammals.
Ms. Joan Wakefield provided invaluable editorial assistance. This content is solely the responsibility of the authors and does not necessarily represent the official views of either the National Institutes of Health or the Department of Veterans Affairs.