Journal of Probiotics & Health
Open Access
ISSN: 2329-8901
ISSN: 2329-8901
Research Article - (2018) Volume 6, Issue 1
Aging is associated with an impairment of the regulatory systems, including the nervous and immune systems as well as the communication between them. The consumption of fermented milk containing live probiotic bacteria seems to have neurological and immunological benefits. The aim of the present study was to evaluate the effects of the supplementation with a fermented milk containing yogurt cultures and the probiotic Lactobacillus casei DN-114001 on behaviour and immune functions of old mice as well as on their life span. Also, the in vitro properties of these probiotics on functions of peritoneal leukocytes were analysed. Old female ICR-CD1 mice were supplemented with fermented milk containing probiotics. After 1 and 4 weeks of supplementation, behavioural tests were performed and immunological functions in peritoneal leukocytes studied. The results showed that a short-term (one week) supplementation with fermented milk containing probiotics was able to improve behavioural aspects (such as motor coordination, equilibrium, muscular vigour, exploratory activity and anxiety-related behaviour) as well as immune functions in old mice that had been impaired by age, reaching similar levels to those found in adult controls. Furthermore, a longer-term supplementation (four weeks) was able to maintain these improvements in the immunological parameters. Probiotics seem to have a direct effect on several immune functions. In conclusion, the supplementation with fermented milk containing probiotics could be a good nutritional strategy to improve behaviour and immune system functions in old individuals and, consequently, to promote healthier aging.
Keywords: Aging; Fermented milk with probiotics; Behaviour; Immune system functions
The impairment in aging of the immune and nervous systems has been associated with increased susceptibility to infectious diseases, cancer, and neurodegenerative diseases [1]. It is well known that the nervous and immune systems are intricately connected and share a bidirectional communication. Thus, any alteration exerted on the immune system will promote an effect on the nervous system and vice versa [1,2]. With respect to the central nervous system, there is an agerelated progressive deterioration of a variety of brain and behavioural functions, including memory, cognitive, and motor activity [3,4]. Moreover, old individuals become more susceptible to suffer mental disorders, such as anxiety [5]. Also, the immune system undergoes a variety of changes, denominated immunosenescence, which include altered innate and adaptive immune functions, such as a marked decreased activity of phagocytic cells, anti-tumour natural killer (NK) cytotoxicity and proliferation of lymphocytes [6]. Increasing evidence suggests that the age-related deterioration of the nervous and immune systems are mainly based on oxidative and inflammatory stresses (an imbalance between oxidant and pro-inflammatory compounds versus antioxidant and anti-inflammatory compounds in favour of the former). Although the production of low levels of oxidants and inflammatory compounds is essential for effective immune response, an overproduction of these compounds by immune cells may lead to cellular and tissue damages [7]. In fact, an oxidative-inflammatory theory has been proposed to explain the cause of aging. Moreover, in this theory, the immune system has been described as a critical player in these stresses and consequently in the rate of aging [8].
The composition and functionality of gut microbiota are also affected by aging, the gut microbiota diversity of aged individuals being lower compared with younger people [9,10]. There is also a decrease in beneficial species, such as Bifidobacteria [11]. In this sense, the consumption of fermented milk containing probiotics has gained much attention from scientific and commercial communities [12]. Probiotics are live microorganisms that are claimed to confer health benefits when administered in adequate amounts [13]. Many reports have indicated that certain strains of probiotics are capable of stimulating and modulating the immune response of the host. Thus, there is an increased application of probiotics to treat immune impairments, such as allergic, inflammatory and autoimmune disorders [14-18]. Several studies also indicated anti-aging (ameliorating immunosenescence) [19] and anti-tumour proprieties of probiotics [20]. In addition, growing evidence suggest that probiotics could also affect brain and behavioural functions [21-25]. It seems that probiotics through the modulation of gut microbiota, or directly, could beneficially affect the bidirectional communication between gut and brain, from which has emerged the concept of microbiota-gut-brain axis [21].
Many studies have been conducted in order to analyse the immune modulating properties of probiotics in adults, and several on their behavioural effects. However, little research has been developed to study particularly the repercussion of probiotics on these aspects in older individuals, as well as on life spans, for which is necessary to use experimental animals with very shorter longevity than humans. Thus, the aim of the present work was to evaluate the effects of the supplementation with a fermented milk containing yogurt cultures and the probiotic Lactobacillus casei DN-114001 on behaviour and immune functions of old mice as well as on their life span. The in vitro properties of these probiotics on some functions of peritoneal leukocytes were also analysed to detect possible direct effects.
Animals
Old (80 ± 4 weeks of age) and adult (32 ± 4 weeks of age) female ICR/CD1 mice (Mus musculus) (Janvier SAS, Chassal, Francia) were used. The animals were housed in polyurethane cages (5 animals per cage) and maintained under standard laboratory conditions (12:12 h reversed light/dark cycle; lights on at 8:00 pm, relative humidity of 50-60%, temperature of 22 ± 2°C and adequate ventilation). All mice were fed a standard maintenance diet (Teklad Global 14% Protein Rodent Maintenance Diet, Harlan Interfauna Iberica, Barcelona, Spain) and water at libitum. The experiments were conducted in accordance with the guidelines and protocols of the Royal Decree 53/2013 regarding the care and use of laboratory animals for experimental procedures, and were approved by the Committee for Animal Experimentation of the Complutense University of Madrid.
Experimental groups
Old mice were separated into three groups (n=8-10, per group), namely (1) old mice which were not dietary supplemented; (2) old mice supplemented with fermented milk containing probiotics (a commercial fermented milk drink during four weeks); (3) old mice supplemented with skimmed milk containing the same amount of vitamins B6 and D as the commercially fermented milk for four weeks. An additional group of adult mice (n=8-10) was used as age controls. The commercial fermented milk drink studied contains two bacterial strains commonly used in yogurt, Lactobacillus delbrueckii subsp. Bulgaricus and Streptococcus thermophilus , and the probiotic Lactobacillus casei DN114001 (109 CFU/100 g). The vitamins B6 and D were also present (0.21 mg/100 g and 0.75 μg/100 g, respectively). Weight, food and supplementation intakes were measured every day. After one week of dietary supplementation with ferment milk (probiotics) or skimmed milk (vitamins), a variety of tests and assays were performed in order to evaluate the behaviour of animals and peritoneal leukocytes were obtained for the analysis of several functions. Given that mice are capable of remembering recently performed behavioural tests, only the immunological functions were studied after four weeks of dietary supplementations. Furthermore, in parallel with these groups, another group of old mice (n=5) was used in order to test the in vitro effect of probiotic, vitamins and other components, present in the fermented milk, on several functions of peritoneal leukocytes.
Fermented milk supernatant for the in vitro immunoassays
A sample of the fermented milk drink was centrifuged at 3600 g for 10 minutes at 4°C and two successive centrifugations under the same conditions were performed from the resulting supernatant. The supernatant, composed of microbial probiotic strains (Lactobacillus delbrueckii subsp. Bulgaricus , Streptococcus thermophilus , Lactobacillus casei DN114001), vitamins and other components of the fermented milk drink, was used to evaluate the chemotaxis capacity of lymphocytes and the natural killer cytotoxicity, as well as assessing its chemoattractant activity.
Behavioural tests
Behavioural tests were carried out between 8 am and 10 am during two consecutive days, followed a procedure previously described [2]. On the first day, a battery of tests (reflexes, T-maze, wood rod, corner and tightrope tests) was performed. On the second day, mice were subjected to the holeboard test. The experimental apparatus was cleaned before the performance of each mouse in order to avoid olfactory trails.
Reflexes: Visual and hindlimb extensor reflex: When the mouse was suspended by the tail and lowered toward a solid black surface, the visual placing reflex was evaluated by observing the complete extension of the forelimbs, and the hindlimb reflex was measured by the complete extension of these limbs. The mean response was rated in three trials.
Motor coordination and equilibrium abilities: In order to assess motor coordination and equilibrium, animals were placed in the centre of a wood elevated rod (rod dimensions: 22 cm height, 80 cm length, 2.9 cm width, divided in segments of 10 cm) for one trial of 60 s. Motor coordination and equilibrium were measured by the percentage of mice falling off the rod. In addition, the percentages of mice that cover at least 1 segment and that complete the test were also considered as motor coordination and equilibrium abilities.
Muscular vigour and traction: As an index of muscular vigour, mice were hanged by its forelimbs in the middle of an elevated horizontal tightrope (40 cm height, 60 cm length and divided into 6 segments of 10 cm) [2]. This method is used to evaluate the muscular vigour and traction in one training trial of 5 seconds and a test trial of 60 seconds. Muscular vigour was assessed as the percentage of mice falling off the rope and as the percentages of mice that cover at least 1 segment and that complete the test. Traction was evaluated by analysing the different parts of the body that mice used to remain hanged (forelimbs, hindlimbs and tail) and, subsequently, the percentages of mice displaying the maximum traction capacity (using forelimbs, hindlimbs and tail).
Non-goal-directed: Behaviour vertical exploratory activity: The vertical exploratory activity (studied as the performance of “rearing”, that is, when the mouse stands up on his hindlimbs so that his body becomes more perpendicular to the ground) was evaluated in the corner test and in the holeboard test. The corner test consists in placing the mouse in a new cage (22.0 × 22.0 × 14.5 cm) with bedding during 30 s. In contrast, the holeboard test consists in placing the animal in a box made of wood (60 × 60 × 45 cm) with matte-painted metallic walls, divided into 36 squares (10 × 10 cm), bearing four equally spaced holes (3.8 cm of diameter). All but 20 peripheral squares were considered central. The test was performed during 5 minutes [2].
Horizontal exploratory activity: Horizontal exploratory activity was assessed as the number of corners of the cage visited (in the corner test) and as the peripheral locomotion (number of line crossings in the peripheral area, by the walls) and central locomotion (number of line crossings in the central area) in the holeboard test. In addition, horizontal exploratory behaviour was also recorded using the T-maze test, which is a wooden apparatus with three enclosed arms (short arm: 27 × 10 cm; long arm: 64 × 10 cm; walls: 19 cm high) [2]. The mouse was placed inside the “short” arm of the maze with its head facing the wall. The time elapsed (in seconds) until the animal crosses with both hindlimbs the intersection of the T-maze and the time (in seconds) to explore the entire maze (exploration of all three maze arms) were assessed.
Goal-directed exploratory behaviour: The goal-directed exploratory behaviour of mice was assessed in the holeboard test. An object formed by a white plastic was placed into each of the four holes of the holeboard to attract mice attention. The frequency (percentage and number) of mice that perform head-dips (when the animal places its head into the hole) was considered as a goal-directed exploratory behaviour.
Behaviours related to anxiety-like behaviour: Behaviour related to anxiety-like behaviour was measured in holeboard and T-maze tests. The total number and percentage of mice that perform self-grooming (when mouse cleans, licks or scratches its body) in both tests and the presence of defecation in the T-maze test were recorded. This last parameter was also considered as an index of anxiety-like behaviour.
Collection of peritoneal leukocytes
The peritoneal suspensions were obtained between 8 am and 10 am to minimize circadian variations in the immune system, by a procedure previously described, without animal sacrifice [26,27], which allowed monitoring the life span of the mice. Briefly, 3 ml of Hank´s solution, adjusted to pH 7.4, were injected into the peritoneum, the abdomen was massaged and the peritoneal exudate cells were collected allowing the recovery of 90-95% of the injected volume. The peritoneal leukocytes, consisting of lymphocytes and macrophages, were counted in Neubauer chambers (Blau Brand, Germany). The suspensions were adjusted to a final concentration of 5 × 105 macrophages or lymphocytes/ml in Hank´s solution or 106 leukocytes/ml in Hank’s solution or complete medium (RPMI 1640 enriched with L-glutamine (PAA, Pasching, Austria) and supplemented with 10% heat-inactivated (56°C, 30 min) fetal calf serum (PAA) and gentamicin (100 mg/ml, PAA) with or without phenol red, depending on the type of assay used. Macrophages and lymphocytes were identified by their morphology. The cellular viability was measured using the trypan-blue (Sigma-Aldrich, Madrid, Spain) exclusion test and in all cases was higher than 98%.
The immune functions parameters studied were performed using unfractionated peritoneal leukocytes, which better preserved the physiological environment surrounding the immune cells in vivo [28].
Chemotaxis assay: Chemotaxis of peritoneal leukocytes was evaluated according to a slight modification of Boyden’s method [26], consisting basically of the use of chambers with two compartments separated by a filter with a pore diameter of 3 μm (Millipore, Bedford, MA, USA). Aliquots of 300 μl of the peritoneal suspensions, with macrophages or lymphocytes adjusted to 5 × 105 cells/ml in Hank’s solution, were deposited in the upper compartment, and aliquots of 400 μl of the chemoattractant, formyl-Met-Leu-Phe (10-8 M) (Sigma- Aldrich), were put into the lower compartment. The chambers were incubated for 3 h and then the filters were fixed and stained. The chemotaxis index (CI) was determined by counting, in an optical microscope (100X), the total number of macrophages and lymphocytes on one third of the lower face of the filters.
Phagocytosis assay: Phagocytosis assay of inert particles (latex beads) was carried out following a method previously described [26]. Aliquots of 200 μl of the peritoneal suspensions adjusted to 5 × 105 macrophages/ml in Hank’s medium were incubated in migration inhibitory factor (MIF) plates (Kartell, Noviglio, Italy) for 30 min. The adhered monolayer was washed with pre-warmed PBS (phosphate buffer saline), and then 200 μl of Hank’s medium and 20 μl of latex bead suspension (1.09 μm, diluted to 1% PBS) (Sigma-Aldrich) were added. After 30 min of incubation, the plates were washed, fixed and stained. The number of particles ingested by 100 macrophages was counted using an optical microscope (100X) and expressed as phagocytic index (PI). The percentage of macrophages, which phagocyted at least one latex bead, was also determined and expressed as phagocytic efficiency (PE).
Natural killer assay: An enzymatic colorimetric assay was carried out to measure the cytolysis of tumour cells (Cytotox 96 TM Promega, Boehringer Ingelheim, Germany) based on the determination of lactate dehydrogenase enzyme (LDH), as previously described [26]. Aliquots of 100 μl of peritoneal leukocytes, used as effector cells, were seeded in 96-well U-bottom culture plates (Numc, Roskilde, Denmark) adjusted to 106 leukocytes per ml in RPMI 1640 medium without phenol red. Murine lymphoma YAC-1 cells, used as target cells, were added adjusted to 105 cells per ml. Thus, the effector/target ratio was 10:1. The plates were centrifuged at 250 g for 4 min to facilitate cell contacts. After 4 hours of incubation, lactate dehydrogenase enzymatic activity was measured in 50 μl/well of the supernatants by addition of the enzyme substrate and absorbance recording spectrophotometrically at 490 nm. Three kinds of control measurements were performed: a target spontaneous release, a target maximum release, and an effector spontaneous release. The results were expressed as percentage of lysis of target cells. To determine this percentage the following equation was used: %lysis=((E-ES-TS)/M-ESTS)) × 100, where E is the mean of absorbance in the presence of effector cells; ES, the mean of absorbance of effector cells incubated alone; TS, the mean of absorbance in target cells incubated with medium alone; and M is the mean of maximum absorbance after incubating target cells with lysis solution.
Lymphoproliferation assay: Following the method previously described [27], aliquots (200 μl) of peritoneal leukocytes (106 lymphocytes/ml complete medium) were seeded in 96 well flatbottomed microtitre plates (Numc, Roskilde, Denmark) and 20 μl of concanavaline A (ConA 1 μg/ml; Sigma-Aldrich), 20 μl of lipopolysaccharide (LPS, E. coli , 055:B5 1 μg/ml; Sigma-Aldrich) or 20 μl of complete medium (spontaneous proliferation) were added per well. After 48 h of incubation at 37°C in an atmosphere of 5% CO2, 0.5 μCi 3H-thymidine (Du Pont, Boston, MA, USA) were added to each well. The cells were harvested in a semiautomatic microharvester 24 h later. Thymidine uptake was measured using a beta counter (LKB, Uppsala, Sweden). The results were expressed as 3H-thymidine uptake (cpm).
In vitro immunoassays for testing microbial probiotic strains and other components of fermented milk
Chemotaxis assay: The chemotaxis index of lymphocytes was assessed as previously described above. However, aliquots of 300 μl of the peritoneal suspensions adjusted to 5 × 105 lymphocytes/ml in Hank’s solution were added to 30 μl of Hank’s solution (control) or to 30 μl of fermented milk supernatant and then deposited in the upper compartment of the chamber. Also, for evaluation of the chemoattractant capacity of lymphocytes, 400 μl of the fermented milk supernatant or aliquots of the chemoattractant, formyl-Met-Leu-Phe (10-8 M) (Sigma-Aldrich) were put into the lower compartment of the chamber. The chemotaxis index (CI) was determined by counting, in an optical microscope (100X), the total number of lymphocytes on one third of the lower face of the filters.
Natural killer assay: An enzymatic colorimetric assay was carried out to measure the cytolysis of tumour cells (Cytotox 96 TM Promega, Boehringer Ingelheim, Germany) as previously described above. Nevertheless, it was used as effector cells aliquots of 100 μl of peritoneal leukocytes (adjusted to 106 leukocytes/ml) with 10 μl of fermented milk supernatant. Murine lymphoma YAC-1 cells (target cells) were added adjusted to 105 cells/ml. The results were expressed as percentage of lysis of target cells.
Statistical analysis: SPSS 21.0 (SPSS, Inc., Chicago, USA) was used for the statistical analysis of the results. The data were expressed as mean ± standard deviation (SD). Each value is the mean of the data from an assay performed in duplicate or triplicate. Normality of the samples was checked by the Kolmogorov-Smirnov test and homogeneity of variances with the Levene test. The data were statistically evaluated by the Student’s t-test for independent and paired samples. P< 0.05 was considered statistically significant and 0.05 < P < 0.1 was considered as a trend.
Body weight, food and drink intakes
There were no significant differences in the body weight after four weeks of dietary supplementation. Thus, old mice supplemented with fermented milk (probiotics) exhibited an average weight of 42 ± 6 g, with skimmed milk (vitamins) of 44 ± 6 g and without supplementation of 42 ± 7 g. Also, there were no significant differences in the average daily food intake during supplementation period among the three groups of old mice (5 ± 1 g/day/mouse, in old mice supplemented with fermented milk, 5 ± 1.4 g/day/mouse, in old mice supplemented with skimmed milk and 5 ± 1 g/day/mouse, in old nonsupplemented controls). However, with respect to the average daily water intake, old non-supplemented controls ingested a significant higher amount of water (10 ± 3 ml/day/mouse) than old mice supplemented with fermented milk (7 ± 2 ml/day/mouse) and with skimmed milk (6 ± 3 ml/day/mouse) (P< 0.001). The average daily supplementation intake during four weeks was 8 ml/day/mouse, for the group of old mice supplemented with fermented milk and 9 ml/day/mouse, for old mice supplemented with skimmed milk.
Behavioural tests
Table 1 shows the results obtained in the behavioural assessments of old non-supplemented mice and old mice supplemented with fermented milk (probiotics) or skimmed milk (vitamins).
| Old mice | ||||
|---|---|---|---|---|
| 1 week supplementation | ||||
| Adult crtl | Old crtl | Fermented milk with probiotics |
Skimmed milk with vitamins |
|
| Reflexes | ||||
| % of mice that show visual placing reflex | 100 | 100 | 100 | 100 |
| % of mice that show hindlimb extensor reflex | 100*** | 86 | 100*** | 100*** |
| Motor coordination and equilibrium (Wood rod test) | ||||
| % of mice falling off | 0*** | 29 | 14*** | 12*** |
| % of mice that cover at least 1 segment | 100*** | 71 | 100***### | 75 |
| % of mice that complete the test | 100*** | 57 | 86***### | 62 |
| Muscular vigor (Tightrope test) | ||||
| % of mice falling off | 14*** | 87 | 71**## | 87 |
| % of mice that cover at least 1 segment | 100*** | 0 | 57***## | 37*** |
| % of mice that complete test | 87*** | 0 | 29***## | 12** |
| Traction (Tightrope test) | ||||
| % of mice that show maximum traction capacity | 87*** | 14 | 57***## | 37*** |
| Non-goal-directed exploratory behavior | ||||
| Vertical exploratory activity | ||||
| % of mice that perform rearing (Corner test) | 100*** | 86 | 100***### | 87 |
| Total number of rearings (Corner test) | 5 ± 1.5 | 3.5 ± 2.5 | 6 ± 1.5* | 4.5 ± 2.5 |
| % of mice that perform rearing (Holeboard test) | 86* | 71 | 100***## | 88** |
| Horizontal exploratory activity | ||||
| Number of corners visited (Corner test) | 7 ± 1.5 | 8.5 ± 2 | 11 ± 1.5* | 9.5 ± 4 |
| Total number of line crossings (Holeboard test) | 217 ± 30 | 222 ± 50 | 267 ± 74 | 229 ± 96 |
| Number of line crossings in the peripheral area (Holeboard test) | 118 ± 26 | 137 ± 40 | 149 ± 54 | 148 ± 74 |
| Number of line crossings in the central area (Holeboard test) | 99 ± 13 | 85 ± 31 | 118 ± 33*# | 81 ± 28 |
| Time for crossing the intersection of the maze (s) (T-maze test) | 10 ± 2 | 10 ± 6 | 7 ± 4 | 14 ± 8 |
| Time to complete the test (s) (T-maze test) | 36 ± 9 | 27 ± 11 | 25 ± 13 | 48 ± 30 |
| Goal-directed exploratory behavior | ||||
| % of mice that perform head-dips (Holeboard test) | 100* | 93 | 100* | 100* |
| Number of head-dips (Holeboard test) | 10 ± 3 | 7 ± 4 | 10 ± 3## | 5 ± 2 |
| Behaviors related to anxiety-like behavior | ||||
| Total number of self-grooming (Holeboard test) | 1 ± 1* | 4 ± 3 | 2 ± 0.5* | 2 ± 1 |
| % of mice that perform self-grooming (T-maze test) | 0*** | 14 | 0*** | 0*** |
| % fecal bolei presence (T-maze test) | 0*** | 29 | 14* | 12** |
Table 1: Behavioural tests in old mice supplemented with fermented milk (probiotics) and with skimmed milk (vitamins) during one week. Results are expressed as percentage (%) or mean ± SD in adult control (n=8-10), old control (n=8-10), old mice supplemented with fermented milk (n=8-10) and old mice supplemented with skimmed milk (n=8-10). *P<0.05; **P<0.01; ***P<0.001 with respect to the values of old control mice. #P<0.05; ##P<0.01; ###P<0.001 with respect to the values of old mice supplemented with skimmed milk.
Reflexes: The hindlimb extensor reflex, which is shown in Table 1, was significantly lower in old controls than in adult controls (P< 0.001). Old mice supplemented with fermented milk (probiotics) or skimmed milk (vitamins B6 and D) during one week exhibited a significantly higher performance of the hindlimb extensor reflex than old nonsupplemented controls (P< 0.001). No significant differences were found among the studied groups of mice regarding the performance of the visual placing reflex (Table 1).
Motor coordination and equilibrium: In the wood rod test, old controls showed significantly impaired motor coordination and equilibrium abilities in comparison with adult controls (P< 0.001, Table 1). However, dietary supplementation during one week with fermented milk (probiotics) or with skimmed milk (vitamins) resulted in significantly lower percentage of old animals falling off from the rod than old non-supplemented controls (P< 0.001, Table 1). In addition, the percentage of old mice supplemented with fermented milk that covered at least 1 segment and that completed the test was significantly higher in comparison with old non-supplemented controls and old mice supplemented with skimmed milk (P< 0.001, Table 1).
Muscular vigour and traction: In the tightrope test, old controls showed significantly impaired muscular vigour and traction capacities in comparison with adult controls (P< 0.001, Table 1). However, the supplementation with fermented milk during one week was able to diminish the percentage of mice that fell off from the rope compared with old non-supplemented controls and old mice supplemented with skimmed milk (P< 0.01, Table 1). With respect to the percentage of mice that covered at least 1 segment and that completed the test (Table 1), old mice supplemented with fermented milk (probiotics) (P< 0.001) or with skimmed milk (vitamins) (P< 0.001 and P< 0.01, respectively) displayed significantly better results than old non-supplemented controls. In addition, both supplementations resulted in maximum traction capacity in old mice in comparison with old nonsupplemented controls (P< 0.001, Table 1). However, old mice supplemented with fermented milk showed significantly better response in all parameters that evaluated muscular vigour and traction than old mice supplemented with skimmed milk (P< 0.01, Table 1).
Non-goal-directed behaviour: Vertical exploratory activity: The vertical exploratory activity of mice, which was assessed by the frequency of rearing, is shown in Table 1. The percentage of old controls that performed rearings in the corner test and in the holeboard test was significantly lower than in adult controls (P< 0.001 and P< 0.05, respectively, Table 1). The one-week supplementation with fermented milk resulted in significantly higher frequency (percentage and number) of rearings in the corner test (P< 0.001 and P< 0.05, respectively) and in the holeboard test (P< 0.001) in comparison with old non-supplemented controls (Table 1). In addition, in the holeboard test, there was a higher percentage of old mice supplemented with skimmed milk that performed rearings in comparison with old nonsupplemented controls (P< 0.01, Table 1). Nevertheless, old mice supplemented with fermented milk showed a higher percentage of rearing performance in the corner test and in the holeboard test than old mice supplemented with skimmed milk (P< 0.001 and P< 0.01, respectively, Table 1).
Horizontal exploratory activity: The results of horizontal exploratory activity are shown in Table 1. In the corner test, old mice supplemented with fermented milk during one week displayed a higher number of corners visited than old non-supplemented controls (P< 0.05, Table 1). With respect to the central locomotion in the holeboard test, old mice supplemented with fermented milk exhibited a significantly higher locomotion than old non-supplemented controls and old mice supplemented with skimmed milk (P< 0.05, Table 1). There were no significant differences in the total and in the peripheral locomotion among the studied groups of mice in the holeboard test (Table 1). In addition, no significant differences were observed among the groups of animals in the performance of the T-maze test (Table 1).
Goal-directed exploratory behaviour: The goal-directed exploratory behaviour of mice, which is shown in Table 1, was significantly lower in old controls with respect to adult controls (P< 0.05). However, the one-week supplementation with fermented milk or with skimmed milk resulted in a higher percentage of old mice performing head-dips than old non-supplemented controls (P< 0.05, Table 1). In addition, old mice supplemented with fermented milk performed more head-dips than old mice supplemented with skimmed milk (P< 0.01, Table 1).
Behaviours related to anxiety-like behaviour: The results of the different tests that assessed behaviours related to anxiety-like behaviour of old mice are shown in the Table 1. Old controls showed a higher frequency performing self-grooming in the holeboard test and in the T-maze test than adult controls (P< 0.05 and P< 0.001, respectively, Table 1). The one-week supplementation with fermented milk or with skimmed milk resulted in a lower percentage of old mice performing self-grooming in the T-maze test in comparison with old non-supplemented controls (P< 0.001, Table 1). In addition, in the holeboard test, old mice supplemented with fermented milk showed a lower number of self-grooming with respect to old non-supplemented controls (P< 0.05, Table 1). The presence of fecal bolei in the T-maze test was significantly higher in old controls in comparison with adult controls (P< 0.001, Table 1). The supplementation with fermented milk or with skimmed milk during one week resulted in significantly lower presence of fecal bolei than old non-supplemented controls (P< 0.05 and P< 0.01, respectively, Table 1).
Immune function parameters
Macrophage peritoneal functions: The peritoneal macrophage functions are shown in Figure 1. The chemotaxis index of macrophages in response to a chemotactic gradient (formylated peptide), which mimics the migration of immune cells towards the infection site, was significantly suppressed in old mice compared with adult mice (P< 0.01, Figure 1A). Old mice supplemented with fermented milk during one and four weeks displayed significantly higher values of the chemotaxis index of peritoneal macrophages (Figure 1A) in comparison with old non-supplemented controls (P< 0.05 and P< 0.001, respectively) and old mice supplemented with skimmed milk (P< 0.05). No significant differences were observed between old mice supplemented with skimmed milk (one and four weeks) and old nonsupplemented controls (Figure 1A).
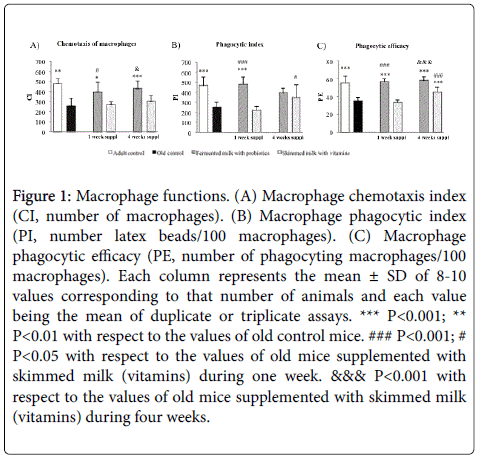
Figure 1: Macrophage functions. (A) Macrophage chemotaxis index (CI, number of macrophages). (B) Macrophage phagocytic index (PI, number latex beads/100 macrophages). (C) Macrophage phagocytic efficacy (PE, number of phagocyting macrophages/100 macrophages). Each column represents the mean ± SD of 8-10 values corresponding to that number of animals and each value being the mean of duplicate or triplicate assays. *** P< 0.001; ** P< 0.01 with respect to the values of old control mice. ### P< 0.001; # P< 0.05 with respect to the values of old mice supplemented with skimmed milk (vitamins) during one week. &&& P< 0.001 with respect to the values of old mice supplemented with skimmed milk (vitamins) during four weeks.
The number of latex beads ingested by macrophages, which is measured by the phagocytic index (Figure 1B), was significantly lower in old mice with respect to adult mice (P< 0.001). The supplementation with fermented milk (probiotics) in old mice, at different times (one and four weeks), resulted in a higher phagocytic index than old nonsupplemented controls (P< 0.001, Figure 1B). The supplementation with skimmed milk (vitamins) was not able to improve this function in comparison with old non-supplemented controls (Figure 1B). However, it was observed a significantly higher phagocytic index in the group of old mice supplemented with skimmed milk (vitamins) during four weeks with respect to the same group supplemented during one week (P< 0.05, Figure 1B). The number of macrophages with phagocytic ability (Figure 1C) was significantly lower in old controls than in adult controls (P< 0.001, Figure 1C). Old mice supplemented with fermented milk (probiotics) during one and four weeks or with skimmed milk (vitamins) during four weeks showed significantly higher phagocytic efficiency than old non-supplemented controls (P< 0.001, Figure 1C). In addition, old mice supplemented with fermented milk (probiotics) during four weeks exhibited higher phagocytic efficiency in comparison with old mice supplemented with skimmed milk (vitamins) (P< 0.001, Figure 1C). There were no statistically significant differences between old mice supplemented with skimmed milk (vitamins) during one week and old nonsupplemented controls (Figure 1C). However, old mice supplemented with skimmed milk during four weeks displayed significantly higher values of phagocytic efficiency than the same group of old mice supplemented during one week (P< 0.001, Figure 1C).
Lymphocyte peritoneal functions: The lymphocyte functions are displayed in Figure 2. The chemotaxis index of peritoneal lymphocytes induced by a chemotactic peptide (Figure 2A) showed significantly lower values in old mice than in adult mice (P< 0.05). The one and four weeks supplementation with fermented milk (probiotics) in old mice resulted in significantly higher chemotaxis index (Figure 2A) than in old mice non-supplemented (P< 0.05 and P< 0.001, respectively) and old mice supplemented with skimmed milk (vitamins) (P< 0.05). Furthermore, old mice supplemented with skimmed milk during four weeks showed higher chemotaxis index of lymphocytes when compared with old non-supplemented controls (P< 0.01, Figure 2A). No significant differences were found between old mice supplemented with skimmed milk during one week and old non-supplemented controls (Figure 2A).
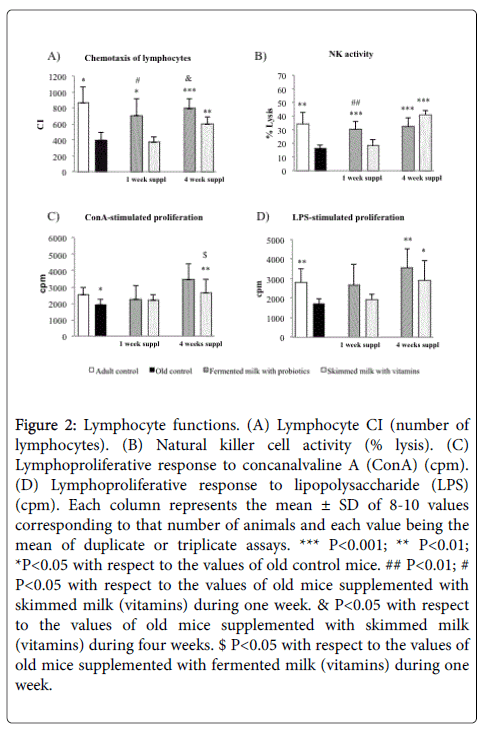
Figure 2: Lymphocyte functions. (A) Lymphocyte CI (number of lymphocytes). (B) Natural killer cell activity (% lysis). (C) Lymphoproliferative response to concanalvaline A (ConA) (cpm). (D) Lymphoproliferative response to lipopolysaccharide (LPS) (cpm). Each column represents the mean ± SD of 8-10 values corresponding to that number of animals and each value being the mean of duplicate or triplicate assays. *** P<0.001; ** P<0.01; *P<0.05 with respect to the values of old control mice. ## P<0.01; # P<0.05 with respect to the values of old mice supplemented with skimmed milk (vitamins) during one week. & P<0.05 with respect to the values of old mice supplemented with skimmed milk (vitamins) during four weeks. $ P<0.05 with respect to the values of old mice supplemented with fermented milk (vitamins) during one week.
The antitumour NK activity (Figure 2B) was significantly suppressed in old mice in comparison with adult mice (P<0.01). Old mice supplemented with fermented milk (probiotics) at different times (one and four weeks) showed significantly higher NK activity than old nonsupplemented controls (P<0.001, Figure 2B). In addition, old mice supplemented with skimmed milk (vitamins) during four weeks resulted in higher NK activity than old non-supplemented controls (P<0.001, Figure 2B). However, old mice supplemented with skimmed milk (vitamins) during one week did not show significant differences in comparison with old non-supplemented controls (Figure 2B).
The lymphoproliferation in response to T-cell mitogen (ConA) and to B-cell mitogen (LPS) (Figure 2C and 2D) was suppressed in old mice with respect to adult mice (P<0.05 and P<0.01, respectively). Old mice supplemented with fermented milk (probiotics) during four weeks displayed significantly higher proliferation of lymphocytes in response to ConA (P<0.01, Figure 2C) and LPS (P<0.01, Figure 2D) than old non-supplemented controls. In addition, it was observed in the group of old mice supplemented with skimmed milk during four weeks higher proliferation of lymphocytes in response to LPS in comparison with old non-supplemented controls (P<0.05, Figure 2D). Nevertheless, old mice supplemented with skimmed milk during four weeks did not show significant differences in the proliferation of lymphocytes in response to ConA in comparison with old nonsupplemented controls (Figure 2C). Furthermore, old mice supplemented with fermented milk or with skimmed milk during one week showed no significant differences in the proliferation of lymphocytes in response to ConA or LPS (Figures 2C and 2D).
In vitro immunoassays: Figure 3 shows the results from the in vitro immunoassays for testing microbial probiotic strains and other components of fermented milk drink. The chemotaxis index of lymphocytes with the presence of fermented milk supernatant, which contains probiotic bacteria, showed significantly higher values than control (P<0.05, Figure 3A). In addition, the chemoattractant capacity of fermented milk supernatant tended to be higher than the chemoattractant capacity of the formyl peptide (P=0.07, Figure 3B). With respect to antitumour NK activity, the fermented milk supernatant added to the effector cells (lymphocytes) showed significantly higher values than control (P<0.05, Figure 3C).
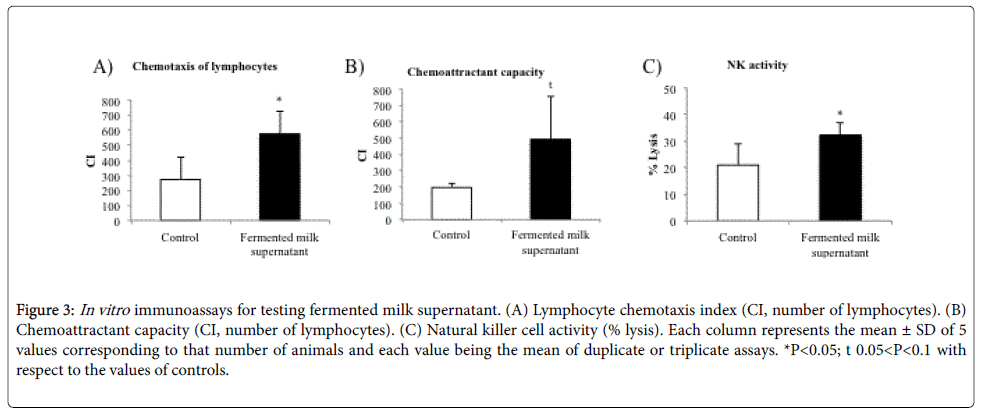
Figure 3: In vitro immunoassays for testing fermented milk supernatant. (A) Lymphocyte chemotaxis index (CI, number of lymphocytes). (B) Chemoattractant capacity (CI, number of lymphocytes). (C) Natural killer cell activity (% lysis). Each column represents the mean ± SD of 5 values corresponding to that number of animals and each value being the mean of duplicate or triplicate assays. *P< 0.05; t 0.05< P< 0.1 with respect to the values of controls.
Life span
The differences were not statistically significant among the groups of mice. The average of life span of old mice supplemented with fermented milk containing probiotics was 108 ± 10 weeks old, with skimmed milk (vitamins B6 and D) was 100 ± 15 weeks and old nonsupplemented controls was 111 ± 15 weeks old.
The consumption of fermented milk containing yogurt cultures plus Lactobacillus casei DN-114001 has demonstrated several immunological and gastrointestinal benefits in the elderly population. Thus, there is evidence that this dietary supplementation is capable of reducing the duration of infections [29,30] and preventing antibioticassociated diarrhoea in old individuals [31]. However, to our knowledge, nothing has been studied so far to show whether this specific fermented milk drink could improve behavioural and immune cell function parameters, which are impaired in older individuals. Therefore, we investigated the effects of the consumption of fermented milk with probiotics on the behaviour and peritoneal leukocyte functions of old mice, as well as their life span. The immune cell function parameters studied here have been used as predictors of health status and consequently of life span. Moreover, the age-related alterations in these parameters are similar to those observed in mice (peritoneal leukocytes) and in humans (blood leukocytes) [6].
The present study found that a short-term (one week) supplementation with fermented milk containing probiotics was able to improve behaviour parameters and immune cell functions of old mice, these reaching similar values to those found in adult controls. Furthermore, a longer-term supplementation (four weeks) was able to maintain these improvements in the same immunological parameters. Nevertheless, the consumption of fermented milk with probiotics did not produce significant differences in the life span of mice. In this sense, it is possible that this consumption at old age is not sufficient to extend the life span, although the benefits found on behaviour and immune response could indicate that this supplementation leads to healthier aging. Similar results were obtained with other lifestyle strategies such as environmental enrichment, which when started in old and adult age improved these parameters of health, but was only able to increase longevity in the latter [32].
The values of the parameters measured in the battery of behavioural tests, which evaluated the abilities of motor coordination, equilibrium, muscular vigour, exploratory activity and anxiety-like behaviour, were significantly impaired in old mice in comparison to adults. However, the one-week supplementation of fermented milk with probiotics was able to improve the performance of these behavioural parameters with respect to old non-supplemented controls. Thus, supplemented animals displayed improvements in motor coordination and equilibrium (assessed by the wood rod test) and in muscular vigour (assessed by the tightrope test). There was also significantly higher exploratory activity, given that old mice supplemented with fermented milk displayed higher percentages of rearings and total number of head-dips in the holeboard test. Moreover, they showed higher locomotion in the central area of the holeboard, which indicates higher exploratory activity and lower anxiety-like behaviour. Previous reports showed that mice tend to stay in the peripheral area (close to the walls) and avoid the central area of the holeboard, which is considered more threatening by these animals [33,34]. In addition, the self-grooming behaviour and the presence of defecation, which is usually considered an index of anxiety in mice [35,36], were significantly lower in old mice supplemented with fermented milk. Currently, there is little investigation about the effect of probiotics on the behaviour of old individuals, given that most of the studies were performed using adults. In these studies, an association was found between the supplementation of probiotics and brain function and behaviour. Thus, Lactobacilli (latic acid bacteria) treatment for two months affected motor behaviour and decreased astrocyte reactivity of growing rats [22]. Also, the probiotic ingestion of Lactobacillus and Bifidobacterium strains has been associated with lower anxiety in adult mice [23,24]. There is also some evidence in humans that probiotics could affect mood and cognition [25]. The mechanisms by which probiotics affect brain function and behaviour remain unclear, but some evidence suggests their association with microbial composition changes, immune activation and production of neurometabolites [21].
The fermented milk with probiotic supplementation used in the present study, was also able to improve macrophage functions of old mice, which suffer an age-related impairment [6], indicating immune-enhancing proprieties of these probiotics. Thus, old mice with the present supplementation displayed enhanced directed migration (chemotaxis) and phagocytic capacity of peritoneal macrophages in comparison to non-supplemented old mice. In agreement, another study demonstrated that the supplementation with fermented milk containing the same probiotics, i.e. yogurt cultures and the probiotic bacteria L. casei DN-114001, for 14 days resulted in increased phagocytic activity in peritoneal macrophages of young mice [14]. In addition, a human-based study showed that the supplementation with the probiotic Bifidobacterium lactis Bi-07 was capable of enhancing phagocytosis of monocytes and granulocytes in healthy elderly individuals [15].
The NK cells, known by their ability to recognize and kill tumour cells, showed a lower activity in old animals [6]. This immune function in old mice improved after supplementation in our study. Previous research on old individuals has also shown the positive effect on NK activity of several probiotics. Thus, the ingestion of a fermented drink containing Lactobacillus casei Shirota for four weeks improved the NK cytotoxicity of healthy aged people [16]. Moreover, another report indicated anti-tumour effects of fermented milk with probiotic consumption [20]. This anti-tumour activity of probiotics seems to be the result of a better immune response in the host (especially due to the improvement of NK cell activity). In addition, the consumption of probiotics appears to favor the decrease of the formation of carcinogenic and mutagenic compounds [20].
The lymphocyte functions, such as migration and proliferation in response to the T cell-specific mitogen (ConA) or the B cell-specific mitogen (LPS), showed lower values in old mice than in adult mice, in agreement with previous results [6]. However, supplementation (one and four weeks) of fermented milk with probiotics led to improvements in the migration of lymphocytes. With respect to proliferation of lymphocytes in response to mitogens ConA and LPS, only a long-term supplementation (4 weeks) with probiotics increased this function in old mice. The effects of probiotics on lymphocyte functions have been more studied using adult subjects. Thus, for example, they increased the response of specific antibodies against infection [17,18]. Nevertheless, there are also several articles showing the positive effects of probiotics on old experimental animals [19, 37,38].
The improvement of the immune function studied seems to be due to the direct action of probiotics since in the experiment carried out in vitro , the fermented milk supernatant containing probiotics enhanced chemotaxis and NK activities. It even showed a chemoattractant capacity.
Interestingly, although with fewer improvements than that with supplementation with probiotics, the control group of old mice, which were supplemented with skimmed milk enriched with vitamins B6 and D, also exhibited some benefits in behaviour and immune cell function parameters (specially after four weeks of supplementation). These results could be expected considering that these vitamins are known by their action on the nervous and immune functions [39-42].
In conclusion, the present study supports the effects of fermented milk containing probiotic supplementation on the function of two major regulatory systems (the nervous and the immune systems). Thus, the consumption of fermented milk with probiotics seems to be a good nutritional strategy to improve several behavioural and immune cell function parameters, which were shown to be impaired in old individuals. For this reason, this strategy could be proposed to promote healthier aging.
The authors declare no conflict of interest.
This work was supported by grants from Ministry of Science and Innovation (BFU2011-30336), the Research Group of Madrid Complutense University (910379ENEROINN), and FIS (PI15/01787) from the Institute de Salud Carlos III-Fondo Europeo de Desarrollo Regional (ISCIII-FEDER) of the European Union.
C.H. is the recipient of a PhD fellowship from CNP-q-Brazil.