Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2018) Volume 6, Issue 2
Many researchers have surveyed damages caused by natural enemies in invasive plants in both native and introduced ranges to testing enemy release hypothesis. Although the physiological impact of natural enemies on invasive plants is important, little research has been conducted to compare the impacts between generalists and specialists. In this study, we report our findings on the physiological and morphological impacts of a native specialist insect (Agasicles hygrophila) and two generalist insects (Atractomorpha sinensis and Hymenia recurvalis) in introduced ranges on an invasive plants Alternanthera philoxeroides in both field trials and controlled environments. Resistances of A. philoxeroides against the generalists and the specialist were also studied. We obtained consistent results in both the field trials and the controlled treatments. Both the generalists and the specialist decreased leaf biomass, photosynthesis, leaf nitrogen content and total leaf non-structural carbohydrate content in A. philoxeroides. However, the specialist decreased leaf mass, photosynthesis, and leaf nitrogen content more acutely than the generalists. Moreover, A. philoxeroides increased both leaf lignin and cellulose concentrations upon the generalists’ attack, but only increased cellulose concentration in response to the specialist. Our results revealed that even under the same population density, the specialists from native ranges caused more severe morphological and physiological damages on A. philoxeroides than the generalists in introduced ranges. Which magnified the consequence of invasive plants suffered lower richness and abundance of natural enemy and contributed to superior performance of invasive plants in introduced regions.
Keywords: Biological invasion; Enemy release hypothesis; Photosynthesis; Resistance; Herbivory
Exotic invasive plant species often attain higher growth rates, extremely greater population densities or broader habitat distributions in their introduced habitats than in their native ranges [1,2]. The enemy release hypothesis explains these phenomena and attributes the success of these exotic plants to the facts that they are liberated from their specialist herbivores and pathogens and that enemies in the introduced regions have a greater impact on native plants than on exotics [3]. Therefore, exotic plants can gain a substantial advantage because their populations are no longer directly suppressed by specialist consumers and pathogens and they also obtain a competitive advantage over native plants that may suffer disproportionately from attacks by enemies in the introduced region.
However, enemy release is limited to the early stages of plant invasion, as native herbivores tend to accumulate on invasive species over time owing to the rapid adaptation of predators to invasive species [4,5]. In addition, native generalist herbivores prefer exotic plants over native plants because exotic plants share no evolutionary history with native herbivores and therefore lack effective defences against them [6-11]. Moreover, the co-evolved specialist enemies of invasive plants might also arrive in the same area either accidentally or deliberately (e.g., through classical biological control releases) [3]. When exotic plants are introduced to a region that contains closely related native congeners, the specialist enemies of those congeners have the potential to attack the exotic plants [12,13].
Herbivory damages the organs and tissues of plants, reduces the leaf area available for photosynthesis, and alters the physiological traits of the affected plant. It can decrease the concentrations of available nitrogen and other important nutrients for foliage and decrease photosynthetic activity [14-16], concluded that the indirect negative effects of reduced photosynthesis (due to herbivory stress) on population biomass were greater than the direct negative effects of herbivores on the loss of plant biomass. To minimize damage, plants have developed resistance and tolerance strategies against herbivores [14], Plants can evolve different strategies against specialists and generalists [17]. However, due to the rapid evolution of invasive plants in introduced regions, invasive plants have either lost or experienced a reduction in the ability to defend themselves against enemies (EICA hypothesis) [18]. Regardless of whether invasive plants have undergone a reduction in their level of protection against specialist herbivores while increasing their protection against generalist herbivores [19,20] or have lost their ability to defend themselves against generalist herbivores [21]. There are differences in the physiology and populations of invasive plants when they are attacked by generalists and specialists.
Alternanthera philoxeroides (Martius) Grisebach (Amaranthaceae) is a perennial amphibious herbaceous plant. The native range of this species is South America, but it has invaded the USA, China, Australia, New Zealand, Indonesia, India and Thailand [22]. In invaded waterbodies, A. philoxeroides often forms dense, interlocking mats and excludes native plants, alters the structure and function of the surrounding ecosystem, blocks irrigation and drainage systems and causes great economic and environmental problems. It has been listed as one of the world’s worst invasive weeds [23,24]. In its native range, A. philoxeroides has more than forty natural enemies [25], among which the specialists feeding on its different organs and tissues regulate its populations [26]. In China, many generalist herbivores also feed on A. philoxeroides [27,28]. However, there have been few studies on the physiological and population-level effects of generalists and specialists on A. philoxeroides and the defence strategies used by A. philoxeroides against these enemies.
In this study, we evaluated how the physiological and population traits of A. philoxeroides respond to herbivory by two generalist insects and a specialist beetle in controlled experiments and investigated some traits used by A. philoxeroides in defence against generalists and specialists. We intended to address the following questions: (a) Do specialists damage populations of A. philoxeroides more intensely than generalists? (b) Do specialists and generalists differ in their physiological impacts on A. philoxeroides ? (c) Does A. philoxeroides defend itself differently against specialists and generalists?
Herbivores
Atractomorpha sinensis Bolivar is a ubiquitous generalist grasshopper native to China. It is distributed in all parts of China and feeds on a variety of vegetables and crops, including cabbage, radish, beans, eggplant, potato, corn, spinach, sweet potato, sugarcane, tobacco, hemp, cotton, rice, and wheat [29]. In field investigations, we found large populations of A. sinensis where A. philoxeroides was concentrated. Hymenia recurvalis (Fabricius) belongs to the Pyralidae family. It is also a ubiquitous generalist insect in China and is distributed across all parts of China, Korea, Japan, India, Sri Lanka, Australia, North America, and Africa. This species feeds on beet, sugarcane, tea, Amaranthus and Chenopodium [30]. The larvae of H. recurvalis chew the mesophyll tissue of vegetables and grasses, but they do not damage the venation. Zhuo-Kai et al. found that the larvae of H. recurvalis fed on A. philoxeroides in Thailand [31,32] also found that the larvae of H. recurvalis prefer plants of the Amaranthaceae family, including A. philoxeroides in China and suggested that H. recurvalis is a potential biocontrol agent for A. philoxeroides .
Agasicles hygrophila Selman and Vogt (Coleoptera: Chrysomelidae) is native to Argentina and Uruguay and is one of the specialist insect herbivores of A. philoxeroides . A. hygrophila was introduced to China in 1987 to control A. philoxeroides . Now, it is found in 16 provinces in central, eastern, and southern China. It can gnaw the leaves and stems of aquatic A. philoxeroides and successfully control its populations, but this herbivore cannot control the populations of A. philoxeroides in terrestrial habitats [33,34].
Field trials
From June 30 to July 3, 2011, various enemies of A. philoxeroides have not appeared yet, we set A. philoxeroides populations in eight 2 m × 2 m plots on Liangzi Island, China (30°001′–30°016′N, 114°032′– 114°043′E) and randomly selected 6-9 shoots of A. philoxeroides in each plot to determine net photosynthetic rate on the youngest fully expanded leaves (generally the second or third pair of leaves from the top) with a Li-6400 Portable Photosynthesis System (Li-Cor, USA). Under a photosynthetic photon flux density (PPFD) of red and blue LED light source at 1700 μmol m-2 s-1 between 11:00 and 14:00, the surrounding air temperature was 30-35°C and the relative humidity ranged 64-72%. Next to these the measured leaves were detached and leaf area was measured using a Li-3100 Area Meter (Li-Cor, USA) to calculate the light-saturated photosynthetic rate per unit leaf area (Pmax).
In August, most populations of A. philoxeroides were damaged by various insects. On August 14, after the dominating herbivore was distinguished, we randomly selected 5-6 shoots of A. philoxeroides damaged by natural insects in each plot and determined net photosynthetic rate on the second or third pair of damaged leaves from the top of each shoot, the surrounding air temperature was 29-34°C and the relative humidity ranged 60-67%. The area of the damaged leaf (Ad) used for the photosynthesis measurement was measured, Pmax was also calculated. To determine the original area of the damaged leaves before they were attacked (Ao), we traced the outlines of the damaged leaves on paper, cut out the leaf shapes and measured them using a Li-3100 Area Meter. The consumed leaf area (Ac) was calculated as: Ac=(Ao-Ad)/ Ao × 100%.
Controlled experiments
Controlled experiments were conducted in The National Field Station of Freshwater Ecosystem of Liangzi Lake, Hubei Province, China (30°50′-30°180′N, 114°210′-114°390′E). In April, 150 shoots of A. philoxeroides were collected from Liangzi Lake. These shoots were cultivated in circular basins with sandy sediment and 5 cm of water. One week later, 36 plants of similar height and weight (mean height 11.56 ± 1.15 cm; mean fresh weight 1.38 ± 0.185 g) were transferred to 18 aquaria (L × W × H=100 cm × 50 cm × 70 cm) filled with 15 cm of fine-textured, homogeneous sediment soil. Two plants were planted in each aquarium and all aquaria were placed on an outdoor cement platform. After 16 weeks, the two plants in each aquarium had formed a single population and all leaves were intact with no herbivore bite marks. The populations of A. philoxeroides were then exposed to the generalist and specialist herbivores. In the generalist treatment, we randomly selected six aquaria and placed twenty 2nd to 3rd instar larvae of A. sinensis and twenty 3rd instar larvae of H. recurvalis on the A. philoxeroides in each aquarium. In the specialist treatment, we randomly selected six aquaria and placed twenty 2nd instar larvae and twenty adults of A. hygrophila on the A. philoxeroides in each aquarium. The six remaining aquaria were controls. All herbivores were collected from populations of A. philoxeroides at the National Field Station of Freshwater Ecosystem of Liangzi Lake and starved for one day before the experiment. All aquaria were covered with a white nylon net (mesh size: 1 mm2) during the experiment.
Three weeks later, the net photosynthetic rate was determined for one of the second or third pairs of damaged leaves from the top at between 11:00 and 14:00 using a Li-6400 Portable Photosynthesis System (Li-Cor, USA) under a red and blue LED light source with a photosynthetic photon flux density (PPFD) of 1700 μmol m-2 s-1, a surrounding air temperature of 35-38°C and a relative humidity of 50-60%. The leaves used in the photosynthesis measurements were marked and used again at between 19:30 and 21:00 to determine the maximal quantum yield (Fv/Fm) using a DIVING-PAM (WALZ, Germany). Next, these leaves were detached; the leaf area was measured using a Li-3100 Area Meter (Li-Cor, USA) to calculate the Pmax. Then the leaves were dried at 70°C for more than 48 h and weighed to calculate the specific leaf area (SLA).
The next day, the herbivores were removed. All remaining second or third pairs of damaged leaves from the top were detached, dried at 70°C for more than 48 h, and stored at -20°C for chemical analyses. Finally after the number of shoots was determined, the plants were harvested. The leaves, stems and roots were separated, washed, and dried at 70°C for more than 48 h to determine the leaf, stem and root mass as well as the total biomass of the populations.
Leaf nitrogen concentration based on mass (Nmass) and leaf carbon concentration based on mass (Cmass) were quantified using a Euro EA 3000 elemental analyser (Euro Vector, Italy). The leaf polyphenolic concentration was measured using this [35]. The leaf lignin content was determined using the method described by the leaf cellulose [36] and total non-structural carbohydrate (TNC) content were determined using anthrone colourimetry [37].
Statistical analyses
All data were analysed using one-way ANOVA after a test for normality and homoscedasticity was conducted; then, Duncan’s tests were used to compare levels within factors for significance (P<0.05). All analyses were performed using SPSS 13.0 (SPSS Inc., Chicago, IL, USA).
The field trials
Among the five plots being analyzed, the populations of A. philoxeroides in plots 5, 6, 7, and 8 were grazed by A. hygrophila , only plot 4 was grazed by A. sinensis (Table 1). The consumed leaf areas (Ac) were different in these five plots, with the value in plot 4 being the smallest (Table 1). Pmax values were different among different populations, both before and after herbivory. Plot 4 had the least reduced Pmax among all five plots (Figure 1).
| Plot | Habitat | Location | Herbivore | Ac (%) (mean ± SD) |
|---|---|---|---|---|
| 1 | Pool | 30º15.673´N,114°33.514´E | No data | No data |
| 2 | Grassland | 30º15.641´N,114°33.567´E | No data | No data |
| 3 | Cropland | 30º15.798´N,114°33.521´E | No data | No data |
| 4 | Roadside | 30°15.862´N,114°33.538´E | A. sinensis | 11.28 ± 6.61 |
| 5 | Pond | 30°15.919´N,114º33.521´E | A. hygrophila | 24.75 ± 9.64 |
| 6 | Lake | 30°15.854´N,114º33.526´E | A. hygrophila | 16.12 ± 6.10 |
| 7 | Pond | 30°15.864´N,114º33.485´E | A. hygrophila | 22.48 ± 11.74 |
| 8 | Pond | 30°15.667´N,114º33.518´E | A. hygrophila | 19.54 ± 4.57 |
Table 1: Habitation, location, herbivore and consumed leaf area (Ac) of Alternanthera philoxeroides populations in the field trials.
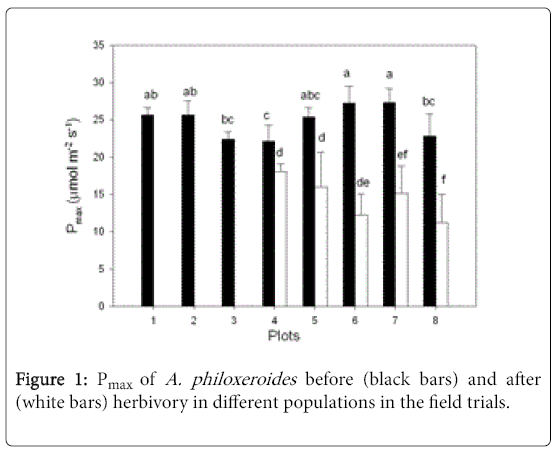
Figure 1: Pmax of A. philoxeroides before (black bars) and after (white bars) herbivory in different populations in the field trials.
The controlled experiments
Both the generalist and specialist herbivores decreased the leaf mass of A. philoxeroides. The specialists had a greater negative effect on the leaf mass than the generalists. Compared with control, leaf mass of A. philoxeroides in generalist and specialist treatments decreased by 42.7% and 84.0% respectively (Figure 2a). Neither the generalists nor the specialists influenced the root mass, stem mass, total biomass or shoot number in A. philoxeroides (Figure 2b-2f). The root and shoot ratio of A. philoxeroides significantly increased when the plants were attacked by generalist and specialist herbivores (Figure 2e).
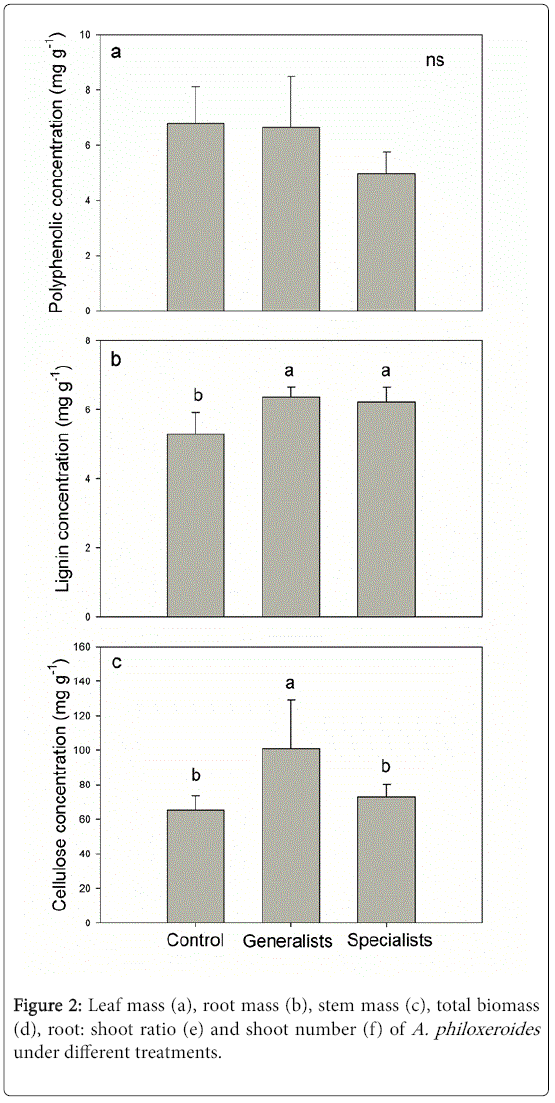
Figure 2: Leaf mass (a), root mass (b), stem mass (c), total biomass (d), root: shoot ratio (e) and shoot number (f) of A. philoxeroides under different treatments.
After herbivory by the generalists and specialists both the Pmax and Nmass of A. philoxeroides were significantly lower than those in the control; the specialists more greatly decreased Pmax and Nmass than the generalists (Figure 3a and 3d). Compared with control, Pmax of A. philoxeroides in general and specialist treatments decreased by 34.4% and 66.9% respectively. The Fv/Fm and leaf TNC content of A. philoxeroides were also significantly lower than those in the control population after herbivory by both generalists and specialists (Figure 3b and 3f). The generalists significantly decreased the SLA of A. philoxeroides , but the specialists had no influence on this parameter (Figure 3c). Neither the generalists nor the specialists influenced the leaf Cmass of A. philoxeroides (Figure 3e).
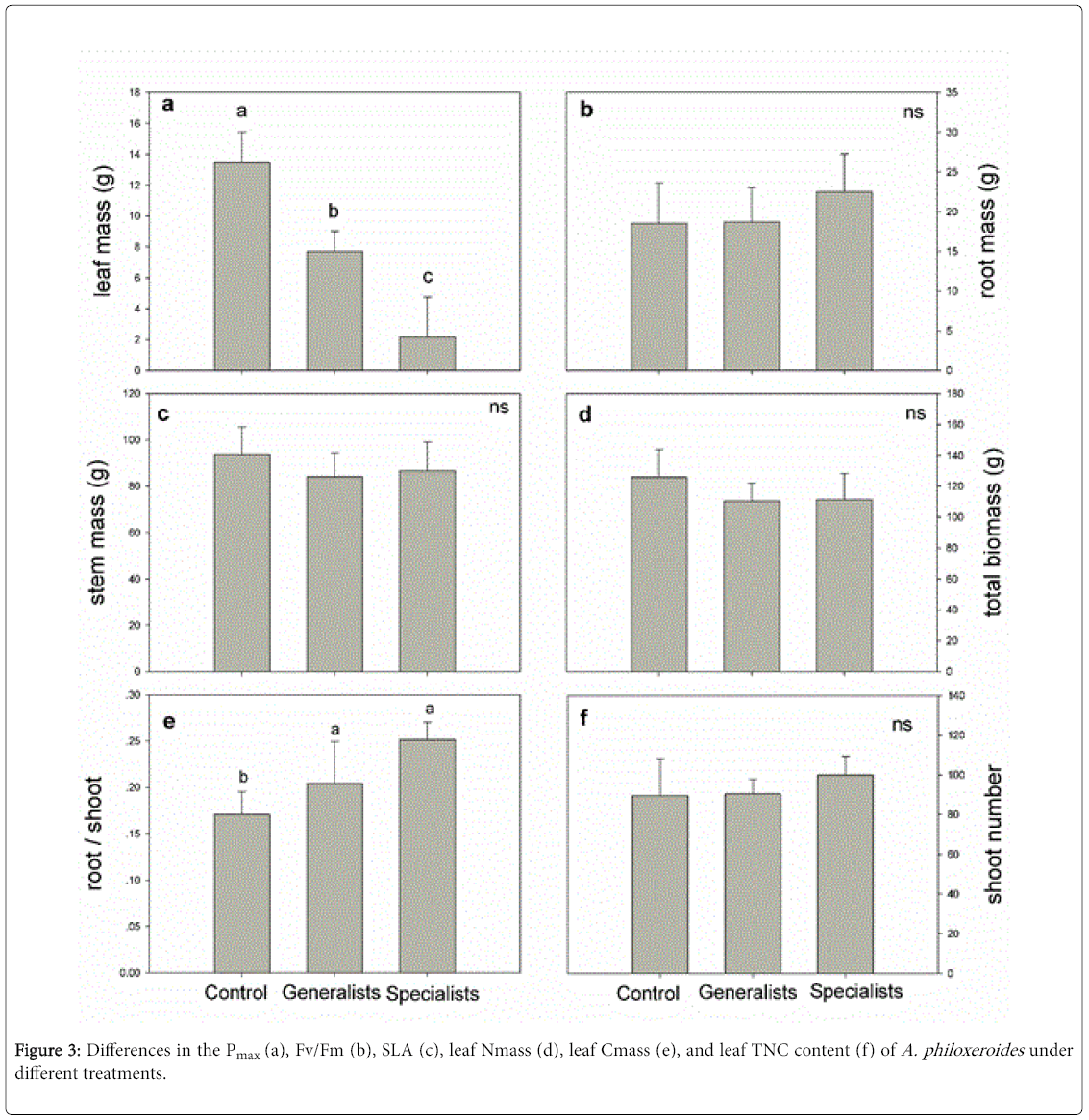
Figure 3: Differences in the Pmax (a), Fv/Fm (b), SLA (c), leaf Nmass (d), leaf Cmass (e), and leaf TNC content (f) of A. philoxeroides under different treatments.
The leaf polyphenolic concentration of A. philoxeroides did not increase to resist attack by the generalists or specialists (Figure 4a).
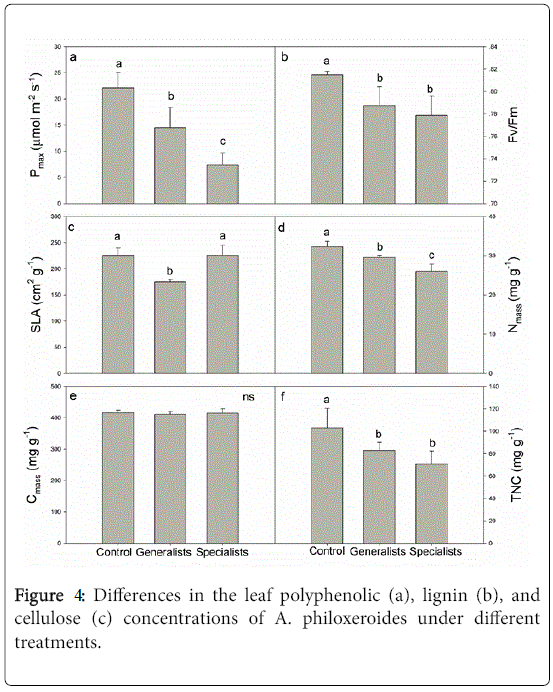
Figure 4: Differences in the leaf polyphenolic (a), lignin (b), and cellulose (c) concentrations of A. philoxeroides under different treatments.
When the generalists were present, the leaf lignin and cellulose concentrations of A. philoxeroides increased, but only the leaf lignin concentration increased in the presence of specialists (Figures 4b and 4c).
In our study, both the generalists and specialists decreased the photosynthetic rate, Fv/Fm, leaf nitrogen and TNC concentration in A. philoxeroides . Pmax is an important leaf trait in plants because the photosynthetic rate is related to various attributes of plant success, including growth, competitiveness, propagation, and resistance to herbivores or pathogens [38]. The insects feeding on leaves not only reduce the leaf area for photosynthesis and but also decrease photosynthesis in the remaining leaf tissues through four mechanisms: severed vasculature, altered sink demand, defence-induced autotoxicity, and defence-induced down-regulation of photosynthesis [16]. The leaf N content is positively related to photosynthesis and relative growth rates [38,39]. The loss of cytoplasm from wounds left by herbivores results in a decrease in leaf N concentration [14]. Moreover, leaf nutritional qualities may determine plant palatability, as herbivores prefer to feed on plants with higher N contents [40,41]. Studies have confirmed that leaf palatability is positively correlated with leaf N content [42,43]. Therefore, an intentional reduction in nitrogen concentrations after attack by herbivores has been suggested to be an anti-herbivore strategy used by plants [44-46] photosynthesis and a loss of cytoplasm can result in lower leaf TNC concentrations in A. philoxeroides .
The populations of A. philoxeroides suffered greater herbivory from specialists than generalists, which contributed to lower leaf mass in the specialist treatments. Leaf photosynthesis was negatively correlated with the degree of leaf damage [15,47]. In addition, studies have shown that after herbivory, the remaining leaf issue closer to the wound has a lower photosynthetic rate than leaf tissue farther away from the wound [15,16]. In our study, the larvae of the generalists, A. sinensis and H. recurvalis , are chewers and tended to gnaw all the leaf tissue in a given area. However, both the adults and larvae of the specialist, A. hygrophila are miners and left holes in random locations on the leaves. From the topology, the remaining leaf area closer to the wound caused by the specialists was larger than that caused by the generalists when both specialists and generalists consumed a given area leaf. As a result, photosynthesis in the remaining leaves after attack by specialists was lower than that after attack by generalists. Meanwhile, the larger wound area caused by the specialists resulted in more cytoplasm loss, which could also have contributed to the lower leaf N and TNC concentrations after herbivory by the specialists than after herbivory by the generalists.
Our results also reveal that A. philoxeroides responded more strongly to the generalists than to the specialists. Although the leaf polyphenolic concentration did not increase to resist the generalists or the specialists in this species, the leaf lignin and cellulose concentrations did increase to resist the generalists and only the leaf lignin increased in response to the specialists. Lignin and cellulose are major components of the plant cell wall. Elevated lignin and cellulose contents increase leaf toughness [48]. Moreover, most herbivores lack the ability to produce the enzymes necessary to decompose cellulose and they therefore cannot digest cellulose [49,50]. These components reduce the palatability of the plant. The specialist herbivore in this study, A. hygrophila is a monophagous beetle herbivore of A. philoxeroides . An increase in the leaf cellulose concentration cannot drive them to feed on other plants. Furthermore, induced defences are costly; they not only cost energy and materials but also affect other physiological processes [51]. A higher leaf cellulose content contributes to a lower SLA in plants, one of the main determinants of relative growth and photosynthetic rates [52,53]. Although both specialists and generalists reduce the leaf mass and physiological traits of A. philoxeroides , they have no influence on the population traits or fitness of A. philoxeroides . In our study, the leaf biomass of A. philoxeroides was found to make up only approximately 10% of the population biomass; stems and shoots constitute most of the biomass, but neither the generalists nor the specialists damaged the stems or roots, and the total biomass and shoot number were not influenced by herbivory. If herbivory treatment persists for a longer time, the decline of photosynthesis and leaf biomass will ultimately affect population performance. In addition, after A. hygrophila consumed all leaves, they will gnaw stems that contributes 75% of the total biomass [33,34]. However, A previous study found that herbivory has a significant and strong negative effect on individual performance and reduces the establishment of individual invaders but cannot completely prevent the further spread of invasive plants once they have successfully established [54]. The herbivory pressure from native generalist herbivores is not sufficient to limit the spread of exotics after they cross a density threshold; therefore, native herbivores fail to completely eradicate exotic plants [55]. The life histories and distributions of the specialist herbivores that are introduced from the original ranges of invasive plants are mismatched with those of the invasive plants in the invaded range [56] and they cannot remain present during the entire life history or throughout the entire range of the invasive plants. In the Yangtze River Basin, A. philoxeroides sprouts at the beginning of May, but the population of A. hygrophila does not emerge until the end of July. Invasive plants always have strong competitive abilities [52] due to their high relative growth and reproduction rates; therefore, A. philoxeroides can rapidly colonize an area before A. hygrophila is present. Moreover, A. philoxeroides shows strong compensatory ability by improving its photosynthetic rate after herbivory [57,58].
In summary, in its introduced range, the defoliation and physiological damage of A. philoxeroides populations caused by generalist herbivores was far lower than that caused by its co-evolved specialist herbivores and could defend against generalists better than specialists. Which further magnified the consequence of invasive plants suffered lower richness and abundance of natural enemy in introduced ranges. We conclude that compared with the exotic plants in original ranges, those in introduced areas, where specialists are absent, have less herbivory stress from the genelralists attack, which contributes to the better performance of exotic plants in introduced areas.
We thank Dr. Jin Yang, Chang Qian and Shuxia Fan for their valuable input and revisions of the manuscript. This study was supported by the National Natural Science Foundation of China (31500295).