Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2015) Volume 2, Issue 3
Arbuscular mycorrhizal fungi support the absorption of mineral nutrients by their host plants in exchange for photosynthate. We evaluated the growth responses of 64 rice cultivars to root colonization by an arbuscular mycorrhizal fungus, Funneliformis mosseae and identified the most positively responsive cultivar as indica rice, ARC5955. We compared the changes of mineral contents of ARC5955 under colonization by the fungus with those of a nonresponsive japonica cultivar, Nipponbare. Colonization increased the contents of beneficial elements such as P and decreased those of deleterious elements such as Cu and Al in both ARC5955 and Nipponbare. However, only ARC5955 showed strong growth stimulation. These results may provide new insights for breeding and yield increase of rice in sustainable agriculture.
<Keywords: Arbuscular mycorrhiza; Funneliformis mosseae; Growth response; Mineral nutrient; Oryza sativa; Symbiosis
AM: Arbuscular Mycorrhiza; DW: Dry Weight; ICP-AES: Inductively Coupled Plasma Atomic Emission Spectroscopy; MGR: Mycorrhizal Growth Response; QTL: Quantitative Trait Locus
Rice (Oryza sativa L.) is not only the most important staple food crop in the world but also an excellent model monocotyledonous plant. Numerous genomic resources including, an accurate genome sequence [1], easy transformation [2], functional characterization of genes [3], and genome editing using the CRISPR/Cas9 system [4], have been developed. Rice is also a suitable material for investigating the molecular mechanisms of symbiosis with arbuscular mycorrhizal (AM) fungi, which colonize the roots of most land plant species [5-9]. However, the number of reports describing the growth response of rice plants to AM fungi is unexpectedly much smaller than those of other crops such as maize, sorghum, legumes, and vegetables [10,11]. The reason may be that rice plants are grown mainly in anoxic paddy fields, in which the presence of AM fungi is still a matter of debate [12-15]. In a laboratory experiment, the colonization of AM fungi decreased under flooding conditions [13]. In contrast, rice seedlings are colonized well under drained upland conditions [12,13,16]. Thus, if we find rice cultivars positively responsive and nonresponsive to AM fungi in well-drained soil and apply genomic methods such as quantitative trait locus (QTL) analyses [3] to two contrasting cultivars, we may be able to identify genes that govern the complex growth response to AM fungi. Wheat, maize, and pearl millet showed diverse growth responses to AM fungi in a number of cultivars [17-19]. In rice, Dhillion described differential responses to AM fungi depending on cultivar, but investigated only three cultivars. In the present study, under upland conditions, we extensively evaluated growth responses to an AM fungus in 64 rice cultivars, most from the NIAS Global Rice Core Collection [20]. We also determined mineral nutrient contents in the rice seedlings by inductively coupled plasma atomic emission spectroscopy (ICP-AES) and discuss factors causing growth stimulation by the AM fungus.
All cultivars used in this study are classified as O. sativa. Seeds of 69 cultivars from the NIAS Global Rice Core Collection [20] were obtained from the National Institute of Agrobiological Sciences, Tsukuba 305-8602, Japan. Seeds of Koshihikari, Sasanishiki, Akihikari, and Nona Bokra were purchased from the National Institute of Agrobiological Sciences based on the advise of Dr. Masahiro Yano. Seeds of Kinnanpu, IR64, IR24, and Akibare were gifts from Prof. Motoyuki Ashikari, Nagoya University. Fourty ml of Kanuma soil, weathered volcanic lapillus with superior drainage property, and appropriate amounts of deionized water were prepared in a cell of a plastic seed tray. A soil inoculant of F. mosseae (1.2 g cell-1; formerly Glomus mosseae ) was added to the cell as described previously [21,22]. The rice seeds were germinated as described [8], and then each seed was transplanted to each cell. The seedlings were grown in a glasshouse under natural light for 4 wk from July 28 to August 26, 2009. Alternatively, they were grown in an indoor cultivation room kept at 28°C under a 16 hr day/8 hr night cycle for 4 wk. The intensity of white fluorescent light was 204 μmol m-2 s-1. Watering was done as needed. In addition, during the growth period, the soil in a cell was supplemented twice with 20 ml each of 0.5x modified Hoagland nutrient solution. Unless otherwise stated, the solution contained 0.1 mM phosphate. Ungerminated seeds or insufficiently grown seedlings were omitted from the analyses.
¨After harvesting, the seedlings were divided into shoots and roots, dried at 80°C for more than 48 hr, and then their dry weights (DWs) were determined. Shoot mycorrhizal growth response (MGR) was calculated following the equation:
The dried tissues (30-60 mg) were ground with mortar and pestle. A portion of each ground sample was digested with HNO3 in a DAP-60K pressure vessel (Berghof GmbH, Eningen, Germany). The vessel was heated successively at 145°C for 5 min, at 190°C for 10 min, and at 100°C for 10 min, with a Speed Wave MWS-2 (Berghof GmbH). The contents of metal elements were then determined by ICP-AES (IRIS ICAP, Nippon Jarrell Ash, Tokyo, Japan). A solution containing 4% (w/v) NaOH and 3% (w/v) K2S2O8 was added to the other portion of ground sample. The sample was heated, as described above, neutralized with HCl, and then we determined the absorption at 220 nm using a spectrophotometer (Shimadzu, Kyoto, Japan). The nitrogen content was calculated in comparison with the absorption of standard nitrate solutions. A one-way ANOVA was used to analyze effects of AM colonization.
As shown in Figure 1, rice seedlings grown under natural sunlight exhibited various MGR to F. mosseae , from −4.4% to 118.9%, depending on cultivar. ARC5955, an indica rice, showed the highest MGR whereas Nipponbare, a popular japonica rice, showed a MGR of only 18.7%. We focused on these two cultivars. No fungal structure was observed in uninoculated roots. However, because MGR and root length colonization were not strongly correlated in our experiment and individual differences in root colonization were large (inset in Figure 1), we did not further examine the colonization. In contrast to F. mosseae , ARC5955 and Nipponbare showed little response to Rhizophagus irregularis (formerly Glomus intraradices), if any (unpublished data). It is well known that plant growth responses to AM fungi depend on the combination of plant and fungus [23].
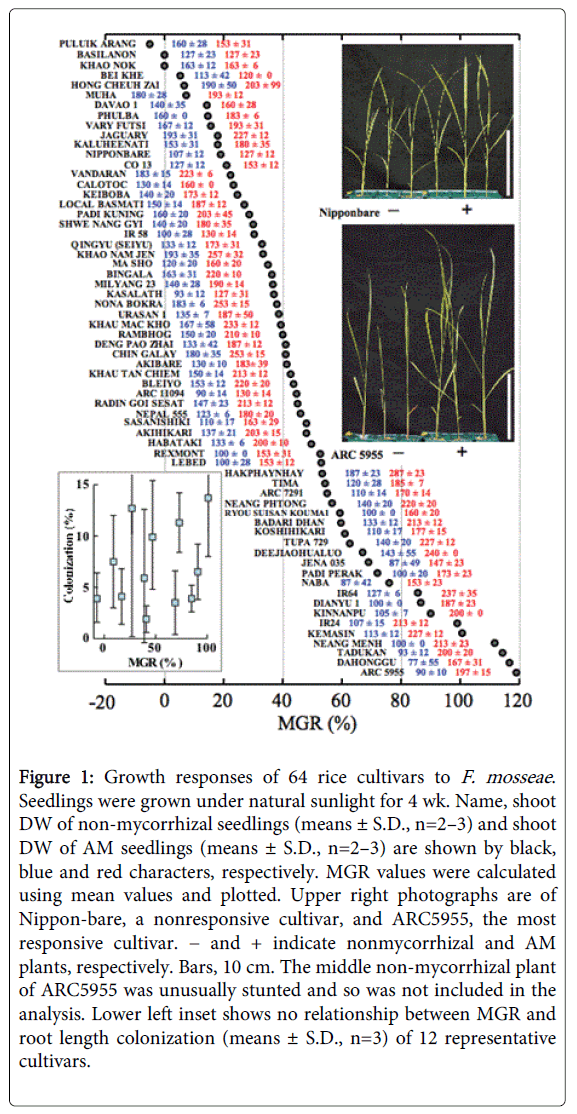
Figure 1: Growth responses of 64 rice cultivars to F. mosseae . Seedlings were grown under natural sunlight for 4 wk. Name, shoot DW of non-mycorrhizal seedlings (means ± S.D., n=2–3) and shoot DW of AM seedlings (means ± S.D., n=2–3) are shown by black, blue and red characters, respectively. MGR values were calculated using mean values and plotted. Upper right photographs are of Nippon-bare, a nonresponsive cultivar, and ARC5955, the most responsive cultivar. − and + indicate nonmycorrhizal and AM plants, respectively. Bars, 10 cm. The middle non-mycorrhizal plant of ARC5955 was unusually stunted and so was not included in the analysis. Lower left inset shows no relationship between MGR and root length colonization (means ± S.D., n=3) of 12 representative cultivars.
These results confirm the pioneering work of Dhillion, and show that rice is positively responsive to AM fungi under well-drained conditions and that colonization and growth response vary widely. Next, we assessed the reproducibility of the different growth responses of ARC5955 and Nipponbare under fluorescent light. As shown in Table 1, ARC5955 was again positively responsive, although the extent of growth stimulation was less than that under sunlight. This reduced response under low light intensity is similar to that of onion [24]. Nipponbare was again nonresponsive. These results enabled us to perform experiments repeatedly in a cultivation room.
| Cultivar | Portion | F. mosseaeDW (mg) | Shoot MGR (%) | |
|---|---|---|---|---|
| Nipponbare | Shoot | - | 110.4 ±10.2 | 15.0 |
| + | 127.0 ±6.7* | |||
| Root | - | 37.2 ±5.6 | ||
| + | 48.8 ±5.0** | |||
| ARC 5955 | Shoot | - | 89.8 ±21.1 | 57.3 |
| + | 141.3 ±34.0* | |||
| Root | - | 37.0 ±9.7 | ||
| + | 55.3 ±14.3 | |||
Table 1: Reproducibility of the growth responses of rice seedlings under fluorescent light at 28°C for 4 wk. DW values are means ±S.D. (n=4–5). ** and *, significance at p<0.01 and p<0.05, respectively.
Table 2 shows the mineral contents of ARC5955 and Nipponbare seedlings. It is noteworthy that colonization of F. mosseae increased P levels in either shoots or roots of both cultivars. The levels of N, S and Mg also tended to become higher in shoots and roots of both cultivars with colonization, but the differences were not significant.
| Cultivar | Portion | F.mosseae | N | P | K | Ca | Fe | S | Mg | Mn | Cu | Zn | B | Co | Al |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nippobare | Shoot | - | 10465 ± 5567 | 376 ± 17 | 17963 ± 1128 | 1560 ± 289 | 173 ± 33 | 1286 ± 162 | 1080 ± 190 | 189 ± 35 | 4.8 ± 1.8 | 47.2 ± 21.3 | 8.5 ± 7.9 | 0 | 1724 ± 392 |
| + | 17203 ± 8712 | 633 ± 203* | 15394 ± 5387 | 1484 ± 461 | 119 ± 48 | 1478 ± 419 | 1187 ± 408 | 98 ± 25 | 3.8 ± 2.6 | 22.0 ± 8.3* | 3.6 ± 0.9 | 0 | 1131 ± 543 | ||
| Root | - | 8681 ± 4904 | 357 ± 13 | 11249 ± 1590 | 2621 ± 1075 | 5989 ± 619 | 1795 ± 245 | 1093 ± 320 | 201 ± 38 | 15.5 ± 1.7 | 109.1 ± 98.3 | 2.6 ± 0.4 | 0.04 ± 0.01 | 56496 ± 8811 | |
| + | 11198 ± 6977 | 615 ± 86*** | 11216 ± 1570 | 1600 ± 646 | 4407 ± 706** | 2076 ± 248 | 1345 ± 497 | 186 ± 82 | 8.9 ± 2.1*** | 26.1 ± 22.8 | 1.9 ± 0.2** | 3.56 ± 2.07** | 39070 ± 7085** | ||
| ARC5955 | Shoot | - | 8015 ± 4167 | 249 ± 66 | 12510 ± 2622 | 1951 ± 482 | 183 ± 42 | 1928 ± 171 | 1142 ± 243 | 159 ± 8 | 5.8 ± 3.8 | 32.4 ± 5.5 | 8.4 ± 3.2 | 0 | 1639 ± 569 |
| + | 14951 ± 16361 | 503 ± 41*** | 16850 ± 1900 | 2188 ± 407 | 151 ± 28 | 2354 ± 329 | 1497 ± 180 | 188 ± 70 | 3.5 ± 2.0 | 29.3 ± 6.8 | 7.3 ± 2.2 | 0.06 ± 0.13 | 1434 ± 428 | ||
| Root | - | 8355 ± 5737 | 373 ± 61 | 14776 ± 4600 | 2768 ± 933 | 3724 ± 308 | 3938 ± 308 | 1366 ± 477 | 256 ± 42 | 28.0 ± 2.3 | 111.4 ± 82.6 | 8.8 ± 2.6 | 0.09 ± 0.08 | 34309 ± 14362 | |
| + | 11911 ± 7963 | 620 ± 58** | 18571 ± 9692 | 2516 ± 1017 | 3500 ± 2233 | 4961 ± 2327 | 1857 ± 906 | 303 ± 144 | 18.0 ± 2.5*** | 64.7 ± 25.2 | 7.3 ± 1.9 | 5.46 ± 1.98** | 22864 ± 9800 |
Table 2: Mineral contents (μg/g DW) of rice seedlings grown under fluorescent light for 4 wk. − and + indicate non-mycorrhizal and AM plants, respectively. Mean values ± S.D. (n=4–5) are shown. ***, ** and *, significance at p<0.001, p<0.01 and p<0.05, respectively.
The levels of K and Mn appeared slightly higher with colonization in ARC5955 but not in Nipponbare, although again the difference was not significant. In contrast to the above elements, the levels of Fe, Cu, Zn, B and Al decreased with colonization. AM fungi mitigate the deleterious effects of Al [25]. Ca level did not change with colonization, nor did Si, Ni, Cd or Cr levels (data not shown). Overall, ARC5955 responded well to the increase of beneficial elements, P, N, S and Mg, and decrease of somewhat toxic elements, such as Cu and Al. In contrast, Nipponbare did not respond to changes in mineral nutrition levels. The reason for this difference is unknown at present. Co contents in AM roots increased significantly in both cultivars (Table 2), but it is also unknown whether this low level of Co affects rice growth.
To confirm the positive response of ARC5955 to increased P content, we grew ARC5955 in the presence of varying concentrations of phosphate in modified Hoagland solution. As shown in Figure 2, ARC5955 grew well in the presence of 2 mM phosphate, concurrently with the increase of P content in the entire plant. Colonization of F. mosseae mimicked the above growth stimulation, and also in-creased the P content. Thus, the positive growth response of ARC5955 to F. mosseae can be explained, at least in part, by the increase of P content in fungus colonization.
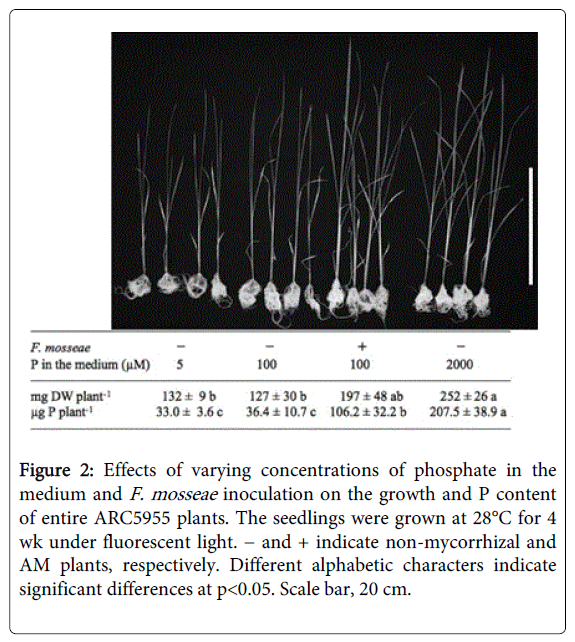
Figure 2: Effects of varying concentrations of phosphate in the medium and F. mosseae inoculation on the growth and P content of entire ARC5955 plants. The seedlings were grown at 28°C for 4 wk under fluorescent light. − and + indicate non-mycorrhizal and AM plants, respectively. Different alphabetic characters indicate significant differences at p<0.05. Scale bar, 20 cm.
AM fungi supply P [5,6,8], N [26], S [27], and other minerals to their host plants [23]. In the present study, we found that colonization of F. mosseae contributed increased contents of these beneficial elements and decreased contents of some deleterious elements. Curiously, these changes occured in both positively responsive and nonresponsive cultivars (Table 2). As pointed out by Smith et al., such differential growth response cannot be explained solely by a “C–P trade imbalance.” We need a new approach, such as QTL analysis, to clarify the mechanism of growth stimulation of rice by AM fungi. Such investigations may also lead to yield increase through breeding in sustainable agriculture with the aid of AM fungi.
We would like to thank the National Institute of Agrobiological Sciences and Dr. M. Ashikari for supplying the rice seeds, Drs. K. Yano and Y. Uenoyama for providing experimental equipment, Drs. T. Suzuki, M. Yano, M. Matsuoka, E. Minami and T. Mato for their advice and encouragement, and Enago (www.enago.jp) for the English language review.