Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2014) Volume 2, Issue 4
In the perennial biomass grass Miscanthus, the rhizome, a below-ground storage organ, is important for its nutrient-efficiency. Here, macro- and microelements in the rhizome and other organs were measured in different seasons. All nutrient concentrations were generally low, indicating high nutrient efficiency to build up biomass, and differed markedly between the organs, including root and rhizome. Active translocation from the shoots increased the rhizome N and P concentrations. After the growth period, most elements remained constant in the shoot. Although the rhizome and the root massively differed in their elemental concentrations, morphology and structure, their proteome was highly similar when analyzed by 2D-gel analysis. Only ~10% of the detected proteins differed, with stress-related proteins found more abundant in the rhizome and glycolysis-related proteins higher expressed in the root. The highly similar proteome of rhizomes and roots suggests major functional similarities in sub-surface organs, despite clear morphological, structural and nutritional differences.
Keywords: Rhizome; Macro and microelement; Nutrition; Remobilization; Rhizome and root proteomics; Proteome
Sustainable biomass production requires crop species that combine low fertilizer inputs with high dry biomass output, but ideally, all nutrients should stay in the field to be available for the next growing season. Furthermore, the environment should benefit from such an agronomic system. This ideotype appears closely matched by the perennial Miscanthus species [1]. Among several genotypes that have been tested across Europe, M. x giganteus has been repeatedly reported as one of the genotypes with best performance and highest yields [2]. Miscanthus x giganteus is a natural sterile hybrid of Miscanthus sinensis and Miscanthus sacchariflorus [3] and combines low N requirements with good agronomic properties and high biomass yields. Peak annual dry masses of up to 40 tons/ha were obtained in southern Europe and in the corn belt of the United States, although the harvested biomass after winter is typically lower, between 15-20 tons/ha [2,4]. In colder environments, it may last several years to reach the maximal output, but this is then stable for 15-20 years. In warmer environments the maximum yield potential is reached within 3 years [2]. After the first year, already about 20 % of the maximal harvestable biomass was obtained in Portugal, while only 4 % of the maximal biomass were obtained in England [2].
Perennial grasses such as M. x giganteus internally cycle or remobilize nutrients annually between above- and below-ground tissues. This seasonal variation of established stands of M. x giganteus appears to be of major importance for the high yields and the low input [5,6]. By use and recovery of 15N-labeled fertilizer, an internal recycling use of 81 % of the total plant N was recorded [7], but this includes recovery of N via the soil. Part of the very high nitrogen efficiency may also come from the interaction with nitrogen fixing bacterial endophytes, such as the associated Herbaspirillum frisingense, which has the genomic requirements to fix nitrogen. This bacterium lacks several factors that may contribute pathogenic characteristics that are found in other Herbaspirillum strains [8]. A strong decline of N (40 %) and P (40 %), but also K (25 %) concentrations from shoots was reported in field experiments in Ireland from late autumn to harvest in spring, which may derive from leaching by rain or active retranslocation to the sub-ground organs [6].
In the fields of established Miscanthus, the roots extended up to 180 cm depth and were for practical reasons mostly neglected in the mentioned studies. However, the total root dry weight was significant and increased from May (10.6 t ha−1) to November by about 30 % and then decreased again until March [9]. In 11-year old stands, only 15 % of the below ground dry mass was root, and 66 % was live rhizome (of 20,7 tons/ha) [10]. After full establishment of the field, less investment is put into rhizome development and the rhizome biomass increased only by 20 % annually.
It is well established that nitrogen fertilization has little impact on Miscanthus yields, especially when the long-term trials were started with high initial N in the soil [11]. However, Miscanthus nutrient efficiency appears also to apply to other major nutrients, such as P, a globally limited resource [12]. Less is known for the growth on soils with different P availability, how this affects other nutrients and whether nutrient remobilization is already relevant in the first year after planting, when the rhizome has just been established.
In this study, the concentrations of macro- and micro-nutrients in glasshouse-grown young M. x gigantheus were quantified in different plant organs. These included root, rhizome, stem and leaves at different physiological stages. Various sampling time points were taken during the year. Three questions were addressed: (1) how do macro- and microelement concentrations change throughout the first year after planting? (2) Is the internal nutrient cycling influenced by the soil? (3) Are the different elemental concentrations associated with different proteomes and functions of (storage) rhizomes and roots?.
M. x giganteus growth
Pot experiments were established in spring 2012 with four treatments (four harvest times see below) and three replications. Two 5 -10 cm rhizomes ofM. giganteus with at least 3 buds were planted into each pot. Each pot was filled with 4.5 kg commercial nutrient rich garden soil (soil characterization see Table 1). In separate experiments, seedlings with similar growth were transferred to 3 Rhizoboxes filled with 25 kg soil: sand (5:4) mix, to monitor root growth [13]. This soil originated from an organically managed tree plantation in southern Germany (Bavendorf, Table 1).
| pH (CaCl2) |
N mg/100g (CaCl2) |
P mg/100g (CAL) |
K mg/100g (CAL) |
Mg mg/100g |
|
|---|---|---|---|---|---|
| garden soil | 5.8 | 68 | 16.6 | 69.5 | 140 |
| sandy loam soil/sand mix | 6.4 | 15.1 | 3.5 | 11 | 14.4 |
Table 1: Macronutrient concentration of used soils.
The experiments were conducted under ambient conditions in a glasshouse at Hohenheim University in Stuttgart, Germany, from May 2012 to June 2013. Because there was no temperature control in the glasshouse, the plants were moved outside next to the glasshouse when the temperature was above 30°C. The planting pots were set up completely randomized. The water content of the soil was kept between 16 % to 22 % (w/w).
Sampling and element analysis
The samples were harvested at four dates, in August and November in year 2012 and April and June in year 2013. Roots, rhizome, stems, upper, middle and lower leaves were harvested separately. To get a representative sample, the samples from different organs were washed, cut into pieces and well mixed. About 10 g from each organ were sampled, dried for three days at 60 °C and ground to fine powder. The concentration of total N was determined using an elemental analyzer (HEKAtech, Wegberg, Germany). For the other macro- and microelements, 0.25 g tissue was ashed at 500°C in a muffle furnace for at least 4 hours. To obtain fully ashed sample, the samples were heated at 500°C for another one hour after some drops of 1:3 HNO3 were added. The ashed samples were dissolved in 1:3 HCl solution. This solution was then used to determine the concentration of K and Ca by a flamephotometer (Eppendorf Elex 6361) and Fe, Mn, Zn, Cu and Mg by an atomic-absorption-spectrophotometer (Thermo scientific iCE 3000). The concentration of total P was determined with the molybdatevanadate method with a spectrophotometer (Hitachi U-3300) at wavelength 436 nm [14].
Isolation and 2D separation ofM. giganteus protein from rhizome and root – Rhizome and root of M. x giganteus were harvested from glasshouse pot experiments in fall (November) and immediately frozen by liquid nitrogen. Total proteins were isolated via a modified protocol [15]. Approximately 4 g of frozen rhizome/root per biological replicate was ground in liquid nitrogen and homogenized in 8 ml fresh-made lysis buffer (0.7 M sucrose; 0.5 M Tris-HCl, pH 8.0; 50 mM EDTA; 0.1 M KCl; 1% DTT; protease inhibitor (Roche), 1 tablet for 50 ml lysis buffer). After 10 min incubation on ice, 8 ml water-saturated phenol (pH 8.0, SIGMA) was added to the sample and mixed on a shaker at 4 °C for 30 min. After centrifugation, the upper phenolic phase was removed into new tube and 5 volumes of 100 mM ammonium acetate in methanol were added to precipitate the protein. After overnight incubation at -20 °C, the protein was pelleted by centrifugation at 16000 g for 5 min at 4°C. The pellet was then washed three times by 80 % acetone and vacuum dried. The dried pellet was solubilized in 500 μl rehydration buffer (7 M urea, 2M thiourea, 2 % CHAPS, 1.25 % Biolyte3-10, 50 mM DTT, protease inhibitor and traces of bromphenole blue). The extracted protein was immediately stored in aliquots at -80°C.
The samples were rehydrated on 24cm Immobiline DryStrips pH 3-11 NL (GE Healthcare) with 1000 μg protein per biological replicate. Isoelectric focusing was carried out at 20 V overnight, followed by 150 V for 2 h, 300 V for 2 h, 300 V-1000 V gradient for 8 h, 1000 V-8000 V gradient for 3 h, 8000 V for 7 h, 8000 V – 50 V gradient to a final setting of 85,700 Vhs. The strips were equilibrated and separated on the basis of their molecular weight in 12 % SDS-page gels using EttanDaltsix equipment (GE Healthcare).
Detection of differential proteins between rhizome and root
After electrophoresis, proteins were stained with a modified Coomassie brilliant blue stain [16] for 72 h on an orbital shaker as previously described [15]. Stained gels were imaged using the Typhoon Trio+ Imaging System (GE Healthcare). The resulting gel image files were exported to Progenesis SameSpots (Non-linear Dynamics). Proteins were accepted as differentially accumulated between rhizome and root when they displayed a change of more than 2-fold and were significant by Student’s test at a significance level of 95% from 3 separate independent experiments for each organ.
Mass spectrometry analysis
Spots were cut from two-dimensional gels, in-gel-digested using trypsin (Roche, Germany) [17] and analyzed by Nano-LC-ESI-MS/ MS performed on an ACQUITY nano-UPLC system (Waters, USA) coupled to a LTQ-Orbitrap XL hybrid mass spectrometer (Thermo Fisher Scientific, Germany). Tryptic digests were concentrated and desalted on a precolumn (2 cm x 180 μm, Symmetry C18, 5 μm particle size, Waters, USA) and separated on a 25 cm x 75 μm BEH 130 C18 reversed phase column (1.7 μm particle size, Waters, USA). Gradient elution was performed from 1% acetonitrile (ACN) to 50% ACN in 0.1 % formic acid within 30 min. The LTQ-Orbitrap was operated under the control of XCalibur 2.0.7 software. Survey spectra (m/z = 250-1800) were detected in the Orbitrap at a resolution of 60.000 at m/z = 400. Data dependent tandem mass spectra were generated for the six most abundant peptide precursors in the linear ion trap. For all measurements using the Orbitrap detector, internal calibration was performed using lock-mass ions from ambient air as described [18]. The Mascot 2.3 (Matrix Science, UK) search engine was used for protein identification. Spectra were searched against the green plant subset of the NCBI protein sequence database downloaded as FASTA-formatted sequences from ftp://ftp.ncbi.nih.gov/blast/db/ FASTA/nr.gz. Search parameters specified trypsin as cleaving enzyme allowing three missed cleavages (cleavage before P was not allowed), a 5 ppm mass tolerance for peptide precursors and 0.6 Da tolerances for fragment ions. Carbamidomethylation of cysteine residues was set as fixed modification and methionine oxidation of proteins was allowed as a variable modification. The proteins were categorized with MERCATOR to perform over-representation analysis [19].
Comparision of transcriptome of rhizome vs. root
Transcritomic sequence raw data were downloaded from the Sequence Read Archive (SRA) in NCBI. The Miscanthus x giganteus fall rhizomes (Accession: SRX322129) and the root (Accession: SRX215392), were compared. The following steps i.e. Mapping, quantification of expressed transcripts, and transcript annotation were performed as described previously according to the description of our previous publication [20].
Nutrients in the rhizome and other organs in two different soils
The nutrient concentrations in the rhizome, root, stem, younger and older leaves of the plants were determined in the active shoot growth phase in the middle of June, where rhizome growth has not yet started [5,6]. A nutrient-rich garden soil and a fertilized sandy loam soil / sand mix with a high organic P fraction (derived from long-term organic farming) were chosen for comparison (Table 1).
Overall, macro- and micronutrient concentrations in plant organs differed strongly in both soils; despite that identical rhizome material was used. In plants grown on the sandy loam soil mix, the P concentrations in all tissues were consistently lower (Figure 1). By contrast, the concentrations of N, Fe, Mn and Cu were elevated in the sandy loam, compared with the garden soil, especially in the roots and the rhizome, but also in old leaf material for Fe and Mn. Furthermore, most nutrients were collectively detected at relative low levels, even in young plant tissue and despite sufficient nutrient supply.
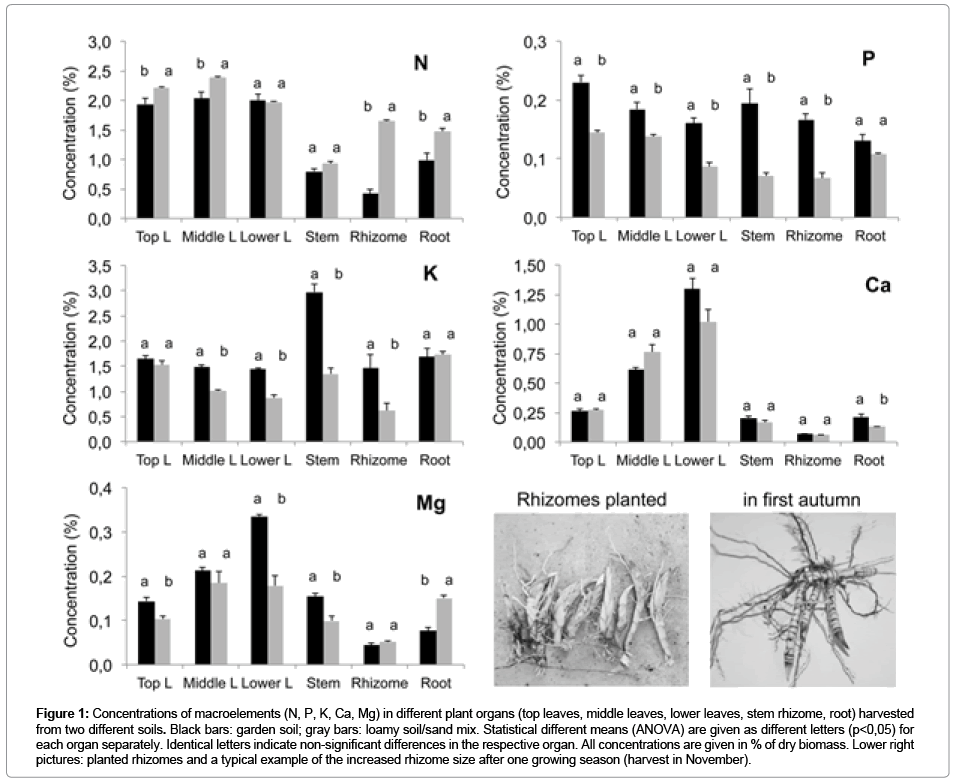
Figure 1: Concentrations of macroelements (N, P, K, Ca, Mg) in different plant organs (top leaves, middle leaves, lower leaves, stem rhizome, root) harvested from two different soils. Black bars: garden soil; gray bars: loamy soil/sand mix. Statistical different means (ANOVA) are given as different letters (p<0,05) for each organ separately. Identical letters indicate non-significant differences in the respective organ. All concentrations are given in % of dry biomass. Lower right pictures: planted rhizomes and a typical example of the increased rhizome size after one growing season (harvest in November).
Only the concentrations of N, Fe and Mn were significantly higher in rhizomes from the sandy loam compared with the other soil, while the concentrations of P and K were lower throughout the plant on that soil (Figure1,2). While the N concentration was high in all leaves, the concentration of P decreased with leaf age. By contrast, K was evenly concentrated in plant tissue grown on the garden soil, except for the doubled concentration in the stem. K was lower in the stem and the rhizome when grown on the sandy loam soil (Figure 1). The micronutrients, as well as Ca and Mg, were massively concentrated in older leaves and lowest in the rhizome (Figure 2).
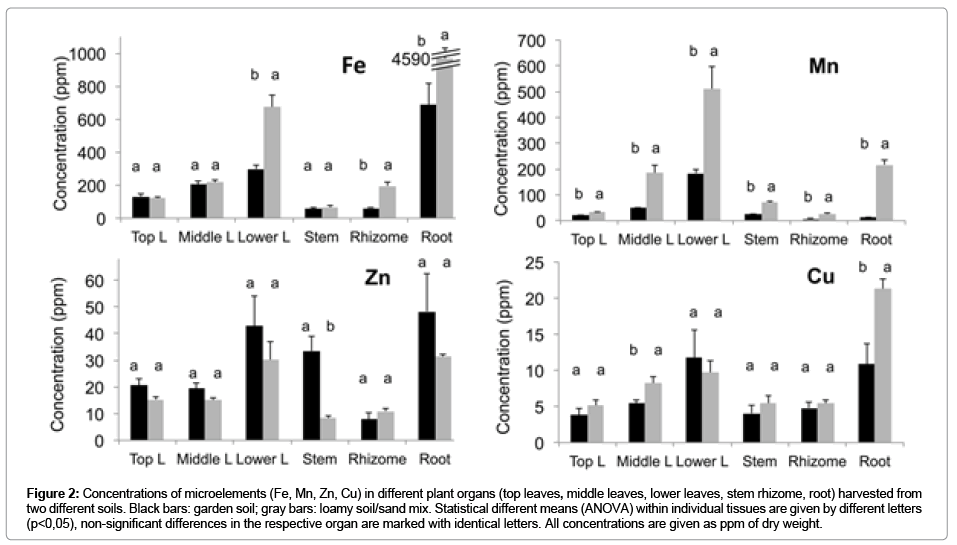
Figure 2: Concentrations of microelements (Fe, Mn, Zn, Cu) in different plant organs (top leaves, middle leaves, lower leaves, stem rhizome, root) harvested from two different soils. Black bars: garden soil; gray bars: loamy soil/sand mix. Statistical different means (ANOVA) within individual tissues are given by different letters (p<0,05), non-significant differences in the respective organ are marked with identical letters. All concentrations are given as ppm of dry weight.
Annual changes in nutrient concentrations in rhizome and root
The element concentrations were further measured in late summer (where shoot senescence had started, especially at the lower leaves), at the beginning and end of the winter, and again in spring. The rhizome mass had steeply increased in autumn, but critically depending on the number of initial buds per rhizome planted. An example of typical rhizomes planted in spring and those harvested in late autumn is given in Figure 1, showing a rough factor of 5-fold increase in rhizome dry weight. Because of heterogeneity among bud number in individual starting material, the biomass yields per rhizome differed largely and no meaningful biomass differences between the soil treatments were obtained.
Compared with the shoot, higher P and K were associated with the growing and storage phase of the rhizome (Figure 3). The concentrations of the mobile Mg was at the highest levels in rhizomes in August, those of the less mobile Mn lowest in June, while the root concentrations of Mg and Mn did not vary annually (Figures 3 and 4). The high rhizome concentrations of Mg and Mn were apparently maintained by uptake from the soil, rather than by nutrient remobilization from the shoot. Fe, Zn and Cu concentrations were much larger in roots in summer and spring (Figure 4), suggesting that the roots were actively acquiring these micronutrients in photosynthetically active plants. Notable massive decreases in the elemental concentrations of shoots were measured at the end of the growing season for N and P, with a concomitant increase in these concentrations in the rhizomes, while no changes in the rhizome concentrations occurred for other mobile elements (K, Mg), compared to the late summer sampling date (Figure 3). The metal micronutrients were apparently not remobilized from the shoots to the rhizome (Figure 4).
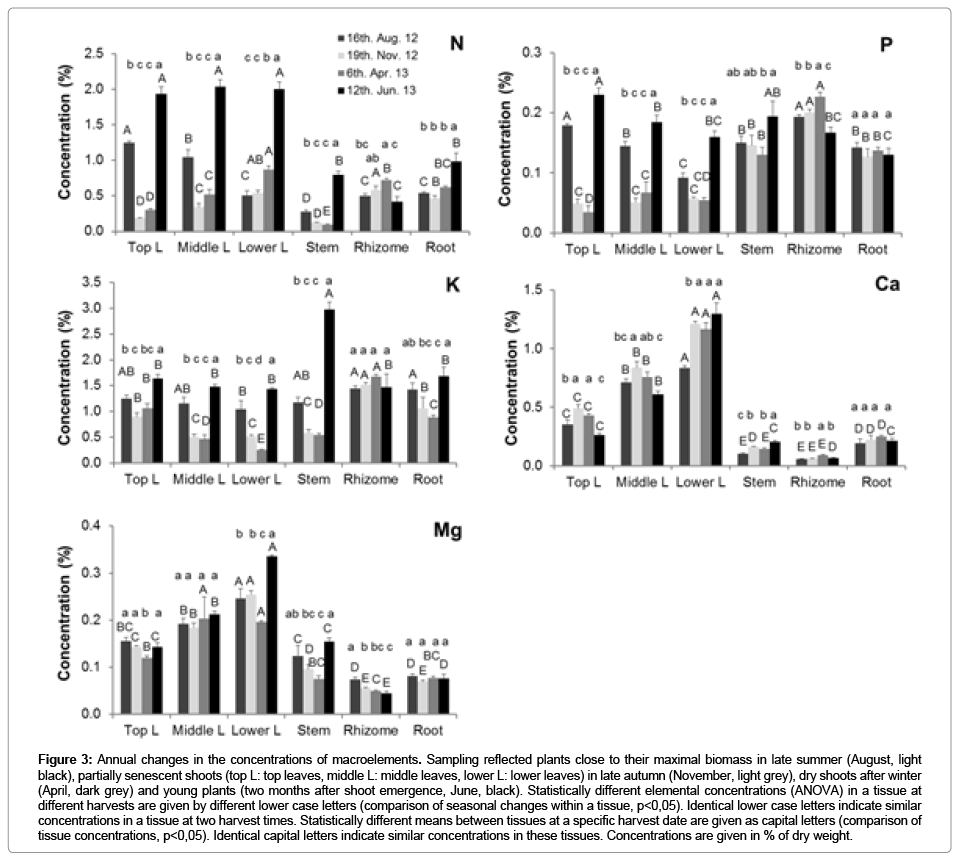
Figure 3: Annual changes in the concentrations of macroelements. Sampling reflected plants close to their maximal biomass in late summer (August, light black), partially senescent shoots (top L: top leaves, middle L: middle leaves, lower L: lower leaves) in late autumn (November, light grey), dry shoots after winter (April, dark grey) and young plants (two months after shoot emergence, June, black). Statistically different elemental concentrations (ANOVA) in a tissue at different harvests are given by different lower case letters (comparison of seasonal changes within a tissue, p<0,05). Identical lower case letters indicate similar concentrations in a tissue at two harvest times. Statistically different means between tissues at a specific harvest date are given as capital letters (comparison of tissue concentrations, p<0,05). Identical capital letters indicate similar concentrations in these tissues. Concentrations are given in % of dry weight.
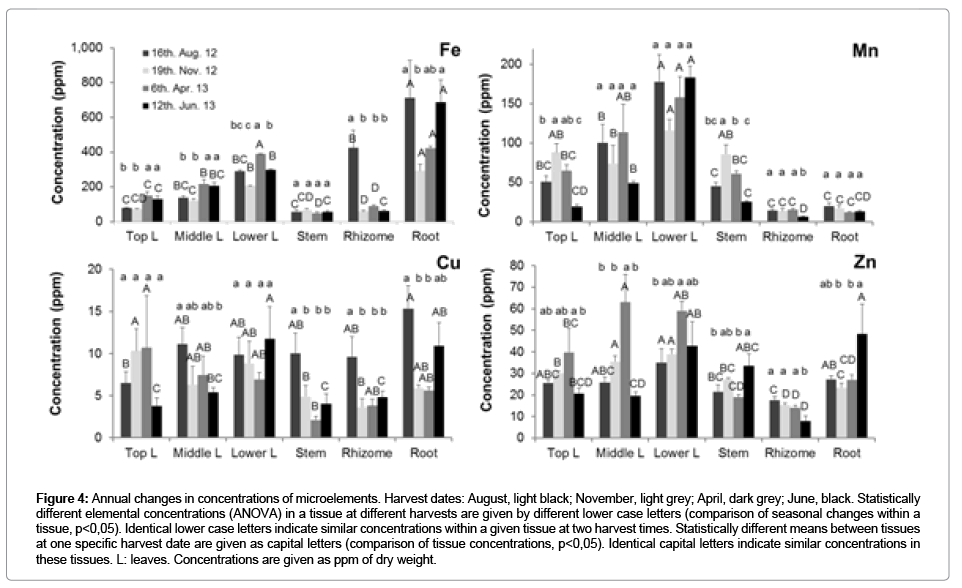
Figure 4: Annual changes in concentrations of microelements. Harvest dates: August, light black; November, light grey; April, dark grey; June, black. Statistically different elemental concentrations (ANOVA) in a tissue at different harvests are given by different lower case letters (comparison of seasonal changes within a tissue, p<0,05). Identical lower case letters indicate similar concentrations within a given tissue at two harvest times. Statistically different means between tissues at one specific harvest date are given as capital letters (comparison of tissue concentrations, p<0,05). Identical capital letters indicate similar concentrations in these tissues. L: leaves. Concentrations are given as ppm of dry weight.
The concentration of the macro elements N, P, K and Mg, but not Ca, in the shoots was higher in summer than in winter (in dead material), reflecting the mobility of these elements (Figure 3). The little mobile elements Fe, Zn, Cu and especially Mn, which are mostly transported via the xylem, had highest concentrations in old leaves (Figure 4).
Fall rhizome and root proteome
The fall rhizome and root proteomes were then isolated and compared. Despite both organs share the same sub-ground habitat, their structure, morphology, elemental composition and cellular components differ strongly. We considered the possibility that the rhizome might contain specific storage proteins or differential expression of metabolic pathways involved in nutrient storage. Surprisingly, only about 10% of the total protein spots identified from 2D-gel separation of the soluble proteome differed significantly (54 spots). The differentially expressed proteins were excised from the gel, but only 38 spots (70%) could be unambiguously identified by multiple peptides by Nano-LCESI- MS/MS (28 among 38 spots in Fig. 5). For the remaining spots, where multiple proteins were found within the same spots in similar quantities, a unique identification was not possible, so that these spots were not further considered (16 spots). For the identified differential proteins, we checked whether these are compatible with transcriptional differences in publically available RNA-seq sequencing data of fall root/ rhizomes (NCBI SRX215392/SRX322129). This comparison of protein and transcriptional differences revealed a large overlap between root and rhizome preference. For 28 out of 38 spots differences in the protein levels were accompanied by transcriptional differences in the same direction. However, the magnitude of the differences in the protein and transcript abundance were not accurately matching, as expected. These differentially expressed proteins are given in Table 2. The annotation and categorization of the proteins preferentially found in rhizomes indicated that major categories with large numbers of significantly changed proteins were “protein” (17 %), “glycolysis” (17 %) and “stressrelated” (12 %) categories (Figure 6). Over-representation analysis indicated that the category of glycolysis was over-represented in roots, whereas the category of stress-related proteins was over-represented in rhizomes (Table 2). This may reflect the different functions of these underground organs in active nutrient acquisition or storage.
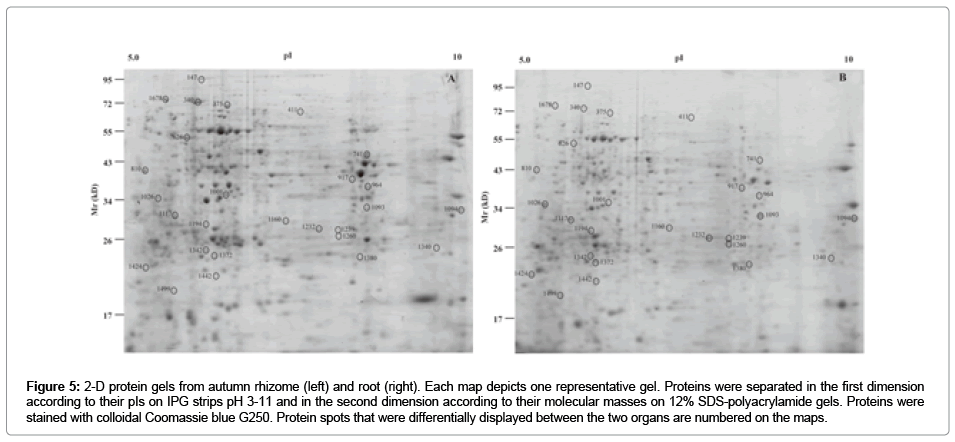
Figure 5: 2-D protein gels from autumn rhizome (left) and root (right). Each map depicts one representative gel. Proteins were separated in the first dimension according to their pIs on IPG strips pH 3-11 and in the second dimension according to their molecular masses on 12% SDS-polyacrylamide gels. Proteins were stained with colloidal Coomassie blue G250. Protein spots that were differentially displayed between the two organs are numbered on the maps.
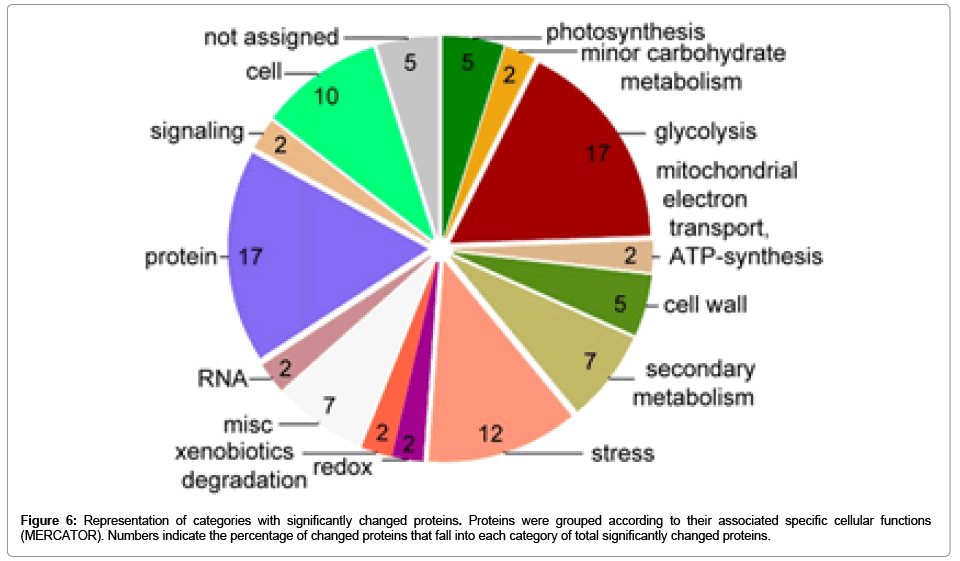
Figure 6: Representation of categories with significantly changed proteins. Proteins were grouped according to their associated specific cellular functions (MERCATOR). Numbers indicate the percentage of changed proteins that fall into each category of total significantly changed proteins.
| Protein No. | Fold ration Rhizome/Root |
Number of unique peptide | Sequence coverage% | Mr gel | pI gel | Function NCBI.nr | Accession | Fold ration Rhizome/Root |
Transcriptome | |
|---|---|---|---|---|---|---|---|---|---|---|
| 375 | 2.4 | 11 | 17 | 72 | 5.8 | heat shock 70 kDa protein [Zea mays] | gi|226500540 | 1.49 | Locus_1820_Transcript_1/1 | |
| 2.38 | Locus_4846_Transcript_1/1 | |||||||||
| 411 | 2.2 | 19 | 30 | 67 | 6.6 | TPA: putative TCP-1/cpn60 chaperonin family protein [Zea mays] | gi|414865473 | 2.56 | Locus_2370_Transcript_1/1 | |
| 1.45 | Locus_3751_Transcript_1/1 | |||||||||
| 626 | 2.0 | 6 | 17 | 50 | 5.5 | alpha-tubulin [Miscanthusfloridulus] | gi|37936230 | 1.88 | Locus_877_Transcript_1/2 | |
| 1001 | 2.3 | 2 | 7 | 35 | 5.8 | glyoxylase1 [Zea mays] | gi|162461576 | 1.89 | Locus_12232_Transcript_1/1 | |
| 1.61 | Locus_264_Transcript_1/1 | |||||||||
| 1678 | 3.1 | 12 | 24 | 72 | 5.3 | stromal heat shock protein [Zea mays] | gi|413916527 | 2.38 | Locus_4846_Transcript_1/1 | |
| 1.59 | Locus_225_Transcript_1/1 | |||||||||
| 741-2 | 2.1 | 5 | 12 | 46 | 7.6 | GTP-binding protein PTD004 [Zea mays] | gi|226528389 | 1.47 | Locus_3211_Transcript_1/1 | |
| 917 | 0.48 | 9 | 20 | 38 | 7.2 | PREDICTED: LOW QUALITY PROTEIN: probable fructose-bisphosphatealdolase 3, chloroplastic-like [Setariaitalica] | gi|514774567 | 0.57 | Locus_280_Transcript_1/1 | |
| Protein No. | Fold ration Rhizome/Root |
Number of unique peptide | Sequence coverage% | Mr gel | pI gel | Function NCBI.nr | Accession | Fold ration Rhizome/Root |
Transcriptome | |
| 1093 | 0.38 | 31 | 7.7 | G3PC_ANTMA RecName: Full=Glyceraldehyde-3-phosphate dehydrogenase, cytosolic | gi|120666 | 0.06 | Locus_8003_Transcript_1/1 | |||
| 1117 | 0.38 | 4 | 10 | 31 | 5.5 | PER1_MAIZE RecName: Full=Peroxidase 1; AltName: Full=Plasma membrane-bound peroxidase 1; Short=pmPOX1; Flags: Precursor | gi|221272350 | 0.007 | Locus_2763_Transcript_1/1 | |
| 0.005 | Locus_3033_Transcript_1/1 | |||||||||
| 1194 | 0.43 | 2 | 5 | 29 | 5.7 | TPA: aspartate aminotransferase isoform 1 [Zea mays] | gi|414885728 | 0.64 | Locus_11214_Transcript_1/1 | |
| 0.46 | Locus_8279_Transcript_1/1 | |||||||||
| 1194-1 | 0.43 | 4 | 12 | 29 | 5.7 | fructose-bisphosphatealdolase, cytoplasmic isozyme [Zea mays] | gi|162462282 | 0.23 | Locus_1412_Transcript_2/3 | |
| 0.08 | Locus_3730_Transcript_1/1 | |||||||||
| 1239 | 0.29 | 4 | 32.5 | 28 | 7.0 | glyceraldehyde 3-phosphate dehydrogenase, putative [Ricinuscommunis] | gi|255591569 | 0.07 | Locus_8003_Transcript_1/1 | |
| 1260 | 0.32 | 3 | 5 | 26 | 7.0 | G3PC1_MAIZE RecName: Full=Glyceraldehyde-3-phosphate dehydrogenase 1, | gi|120670 | 0.68 | Locus_14511_Transcript_1/1 | |
| Protein No. | Fold ration Rhizome/Root |
Number of unique peptide | Sequence coverage% | Mr gel | pI gel | Function NCBI.nr | Accession | Fold ration Rhizome/Root |
Transcriptome | |
| Cytosolic [Zea mays] | ||||||||||
| 1340 | 0.34 | 3 | 7 | 24 | 9.5 | Rab GDP dissociation inhibitor alpha [Zea mays] | gi|308081801 | 0.66 | Locus_1616_Transcript_1/1 | |
| 0.35 | Locus_275_Transcript_1/1 | |||||||||
| 1342 | 0.5 | 3 | 15 | 24 | 5.7 | PREDICTED: chalcone--flavononeisomerase-like [Setariaitalica] | gi|514812950 | 0.31 | Locus_518_Transcript_1/1 | |
| 0.18 | Locus_9893_Transcript_1/1 | |||||||||
| 1372 | 0.43 | 5 | 30 | 23 | 5.8 | actin [Pisumsativum] | gi|1666232 | 0.05 | Locus_10197_Transcript_1/1 | |
| 0.36 | Locus_10784_Transcript_1/1 | |||||||||
| 1380 | 0.45 | 3 | 11 | 23 | 7.4 | PREDICTED: phosphomannomutase-like [Setariaitalica] | gi|514741978 | 0.20 | Locus_609_Transcript_1/1 | |
| 1424 | 0.50 | 5 | 18 | 22 | 5.2 | proteasome subunit alpha type 5 [Zea mays] | gi|226499840 | 0.72 | Locus_12192_Transcript_1/1 | |
| 0.44 | Locus_1984_Transcript_1/1 | |||||||||
| 1499 | 0.37 | 5 | 23 | 20 | 5.4 | glutathione S-transferase I [Zea mays] | gi|168489 | 0.01 | Locus_1377_Transcript_1/1 | |
| 0.14 | Locus_796_Transcript_1/3 | |||||||||
| 1688 | 0.53 | 6 | 21 | 42 | 6.3 | Pectinesterase-2 precursor, putative [Ricinuscommunis] | gi|255581396 | 0.02 | Locus_4445_Transcript_1/1 | |
| 1342-1 | 0.50 | 3 | 11 | 25 | 5.7 | inositol monophosphatase 3 | gi|226493279 | 0.66 | Locus_2052_Transcript_1/1 | |
| Protein No. | Fold ration Rhizome/Root |
Number of unique peptide | Sequence coverage% | Mr gel | pI gel | Function NCBI.nr | Accession | Fold ration Rhizome/Root |
Transcriptome | |
| [Zea mays] | ||||||||||
| 1342-2 | 0.50 | 2 | 9 | 25 | 5.7 | beta 4 subunit of 20S proteasome [Oryzasativa Japonica Group] | gi|8671508 | 0.24 | Locus_25116_Transcript_1/1 | |
| 0.73 | Locus_8657_Transcript_1/1 | |||||||||
| 1380-2 | 0.45 | 2 | 9 | 23 | 7.2 | PREDICTED: gamma carbonic anhydrase 1, mitochondrial-like [Oryzabrachyantha] | gi|573912739 | 0.27 | Locus_5069_Transcript_1/1 | |
| 1442-1 | 0.45 | 4 | 21 | 22 | 5.7 | triosephosphateisomerase, cytosolic [Zea mays] | gi|195608720 | 0.62 | Locus_163_Transcript_1/1 | |
| 0.35 | Locus_3024_Transcript_1/1 | |||||||||
| 917-1 | 0.48 | 6 | 24 | 38 | 7.2 | PREDICTED: peroxidase 1-like [Setariaitalica] | gi|514748421 | 0.01 | Locus_18234_Transcript_1/1 | |
| 0.01 | Locus_4228_Transcript_1/1 | |||||||||
| 1026 | 0.45 | 4 | 13 | 35 | 5.3 | PREDICTED: probable protein phosphatase 2C 10-like [Setariaitalica] | gi|514709867 | 1.0 | Locus_1921_Transcript_1/1 | |
| Locus_2843_Transcript_1/1 | ||||||||||
| 1160 | 0.5 | 5 | 16 | 29 | 6.4 | PREDICTED: probable L-ascorbate peroxidase 7, chloroplastic-like [Setariaitalica] | gi|514801505 | 0.8 | Locus_3408_Transcript_1/1 | |
| 810 | 0.37 | 13 | 16 | 43 | 5.1 | AF304164_1 keratin 1 [Homo sapiens] | gi|11935049 | 1.12 | Locus_16527_Transcript_1/1 | |
| Protein No. | Fold ration Rhizome/Root |
Number of unique peptide | Sequence coverage% | Mr gel | pI gel | Function NCBI.nr | Accession | Fold ration Rhizome/Root |
Transcriptome | |
| 1094 | 0.45 | 3 | 10 | 31 | 5.7 | TPA: RNA and export factor-binding protein 2 [Zea mays] | gi|414588875 | 0.97 | Locus_14032_Transcript_1/1 | |
| 0.45 | 2 | 6 | 31 | 5.7 | PREDICTED: 40S ribosomal protein S2-4-like [Oryzabrachyantha] | gi|573926466 | 1.04 | Locus_5433_Transcript_1/1 | ||
| 0.90 | Locus_6655_Transcript_1/1 | |||||||||
| 1232 | 0.43 | 5 | 40 | 28 | 6.8 | glyceraldehyde-3-phosphate dehydrogenase, partial [Taliparititiliaceum] | gi|83701244 | 1.01 | Locus_292_Transcript_1/1 | |
| 1.25 | Locus_476_Transcript_1/1 | |||||||||
| 741-2 | 2.10 | 5 | 12 | 46 | 7.6 | GTP-binding protein PTD004 [Zea mays] | gi|226528389 | 0.68 | Locus_3211_Transcript_1/1 | |
| 0.81 | Locus_5053_Transcript_1/1 | |||||||||
| 964-2 | 2.50 | 2 | 7 | 37 | 7.8 | PREDICTED: eukaryotic translation initiation factor 2 subunit alpha-like [Setariaitalica] | gi|514820378 | 1.04 | Locus_10188_Transcript_1/1 | |
| 1.39 | Locus_11037_Transcript_1/1 | |||||||||
| 147 | 3.0 | 25 | 33 | 96 | 5.7 | Cell division cycle protein 48, putative, expressed [Oryzasativa Japonica Group] | gi|110289141 | 1.26 | Locus_7934_Transcript_1/1 | |
| 340 | 2.8 | 11 | 20 | 73 | 5.6 | PREDICTED: probable mediator of RNA polymerase II transcription subunit 37c-like [Setariaitalica] | gi|514753044 | 1.27 | Locus_2197_Transcript_1/1 | |
Table 2: Differentially expressed proteins in autumn rhizomes and roots.
Overall, the nutrient concentrations determined here were similar or even slightly lower compared with previous analyses in field trials. When compared with other crops, such as the distantly related sorghum or maize, most elements were close to or below the deficiency threshold in those other crops [21], confirming that Miscanthus is highly nutrient efficient in establishing biomass. In pot trials with two different soils, large differences in the concentrations of several macroand micronutrients were recorded in young Miscanthus plants (Figures 1 and 2).
The lower P concentrations in the plants grown on the sandy loam/ sand mix soil with lower plant available P were correlated with higher concentrations of Fe and Mn and Cu (especially in the roots, Figure 2). This likely is explained by the typical physiological root adaptations of many plants to low P, which involves rhizosphere acidification, organic acid secretion and other factors, which also help to mobilize Fe, Mn and Cu from the soil [22]. The different macro-nutrient allocation and different N, P, K levels in established above-ground and below-ground tissue had been observed earlier on 4 different soils, with variation in the yield and season [23]. While that study measured similar macronutrient concentrations in rhizomes and roots, another study suggested that the major nutrients were lower in the roots, compared with the rhizome [6]. In the young plants investigated here, a clear tendency towards similar or higher nutrient concentrations in the roots was seen (except for P) (Figures 2 and 3), but this also depended on the type of soil. A likely explanation is that older roots with more dead fractions have lower element concentrations.
Rhizomes were identified as significant nutrient storage organs in winter. In previous field trials on established 3-year-old Miscanthus plants, N translocation from the rhizome to the shoot was about 34%, for P 58%, K 28% and Mg 44% [5], which accounts for 21 % N, 36 % P, 14 % K and 27 % Mg of the shoot elements content. In another study, only 9 %, 18 % and 7 % of the shoot N, P, K, respectively, were derived from the rhizomes [6]. By contrast, re-mobilization from the shoots to the rhizome was even more effective, with the rhizome nutrient content increasing between September and March. About half of the rhizome N and P content was derived from the shoot and almost one third of the K and Mg content [5]. In our experiment with one year old young plants, leaf and stem concentrations of N, P, K and Mg (old leaf and stem) were also decreased after summer (Figure 3). However, the concentrations of the phloem immobile Ca and all the measured microelements in the shoot (leaf and stem) were not decreased in winter. The Fe and Zn concentrations were even higher in the winter/ fall leaves, when comparing to that in spring and summer (Figure 4). This suggested that remobilization of these microelements from the shoots is poor. The nutrient remobilization from the rhizome to the shoot can be illustrated from the last two sampling dates after winter (Figures 3 and 4). Compared to the winter element concentrations in the rhizome, the concentrations of N decreased by 42%, P by 27%, Ca by 33% and those of Zn and Mn by 44% and 58%, respectively. This suggests that these nutrients are effectively mobilized from the rhizome to the shoot. All other element concentrations, however, were not significantly different from the previous sampling point. Notably, the concentrations of N, K and Zn, as well as Fe and Cu, increased during the active growth of the roots, suggesting active investment into the uptake of these nutrients from the soil to further supply the shoot. All the above analysis of nutrient remobilization indicates that nutrient concentrations differ in the first year and after several years of establishment [24]. Despite that total element concentrations differed between soils and nutrient availabilities, the nutrient cycling appeared to be little affected by the soil.
In a field trail, the concentrations of N, P (and Mg and K) had dropped by one order of magnitude in fall and decreased further during winter [5]. By contrast, no decrease in these nutrients in winter was measured with plants kept in the glasshouse (Figure 3). Because the plants were not exposed to rainfall, the above results clearly indicate that the previously observed massive decrease of the mobile N, P, K in the established field sites in winter is mostly due to leaching by rainfall, but not due to active export of the nutrients from the shoots to the below-ground organs. Furthermore, the distribution of the little mobile Fe, Zn, Cu and Mn is compatible with their movement within the xylem stream (Figure 4).
The function of rhizomes requires apparently relatively low nutrient concentrations, as these were almost always lower than in stem or in all other tissues (except for K). Different elemental concentrations in the roots and rhizomes and their annual fluctuation were contrasted by similar proteomes of these underground organs. The 2D-separated soluble proteome of rhizome and roots had a surprising overlap in almost 500 protein spots. Only about 10% of the protein spots were differentially expressed, suggesting a marked overlap in the belowground organ protein set and overlapping basic metabolic functions. As expected, the soluble proteome from rhizomes and roots had little overlap with the proteome of field-grown adult Miscanthus leaves [20]. Differential protein expression in the root and rhizome were accompanied and confirmed for most proteins by differential transcript abundance from RNA-seq experiments.
The preferential identification of stress-related proteins in the rhizome proteome may be related to its structure and storage function. As a major function of the rhizome is to provide a nutrient-storing over-wintering organ, metabolic activity is likely suppressed and rhizome cells are desiccated to increase freezing tolerance. By contrast, the more metabolically active root tissue e.g. higher abundance of the glycolysis-related metabolism may be explained by the importance and high energetic demand for shoot-derived assimilates to maintain root function and nutrient acquisition (Table 2). Despite the differences and dynamics in the elemental profiles, no proteins with direct relation to their storage and remobilization capacity for nutrients were identified, although large differences in element concentrations occurred between organs and during the year.
We thank Elke Dachtler and Helene Ochott for help with nutrient analyses. We also thank the Life Science Center (University Hohenheim), especially to Dr. Jens Pfannstiel and Berit Wuertz, for the mass spectrometry analysis.