Anatomy & Physiology: Current Research
Open Access
ISSN: 2161-0940
ISSN: 2161-0940
Research Article - (2018) Volume 8, Issue 1
To investigate a role of proteoglycan (PG) on developing mouse mandibular condylar cartilage, effects of β-D-xyloside, which can inhibit normal PG synthesis, was confirmed in organ culture. Anlagen of condylar cartilage at E14.0 were taken and cultured for 6 days with or without β-D-xyloside. Adding β-D-xyloside remarkably reduced condylar cartilage formation, and formed cartilage showed more fibrocartilaginous characteristics. These results indicated that normal PGsynthesis is required for forming intact condylar cartilage. Thickened perichondrium, however, was compensatory formed around cartilage matrix, and consequently outline of mandibular condyle was considerably maintained. This phenomenon might be derived from characteristics of condylar cartilage as secondary cartilage, and condylar cartilage might have ability to manage to execute its own morphogenesis utilizing materials available.
Keywords: Mandibular condylar cartilage; Proteoglycan; Origin, β-D-xyloside
The mandibular condylar cartilage is classified as secondary cartilage in embryology. Secondary cartilage has several definitions, and most narrow definition is that it arises from the periostea of membrane after (secondary to) bone formation [1,2]. We have demonstrated that newly formed condylar cartilage is derived from alkaline phosphatase-positive periosteum-like tissue, and supported this definition in mice [3,4], rats [4], and humans [4,5]. In addition, mandibular condylar cartilage differs from primary cartilage in the expression pattern of collagen types [6,7], indicating this cartilage has fibrocartilaginous characteristics. Meanwhile, large chondroitin sulfate proteoglycan (CSPG) named aggrecan is a main non-collagenous component of cartilage, and it produces specific nature of cartilage such as elasticity in tracheal wall and resistance to mechanical force in articular cartilage [8]. Thus, we hypothesized that normal PG (aggrecan) synthesis is required for intact morphogenesis of condylar cartilage. To confirm it, we accepted organ culture of developing mandibular condylar cartilage [9,10] with or without β-D-xyloside , which could block normal PG synthesis by inhibiting the incorporation of CS chains into core protein [11].
Animals
Three pregnant Institute of Cancer Research (ICR) mice at E14.0 were obtained from Sankyo laboratories (Tokyo, Japan). The animaluse protocol conformed to the National Institutes of Health guidebook and was reviewed and approved by the Screening Committee for Animal Research of the Tokyo Medical and Dental University (No. 0120394A).
Organ culture experiments and metabolic labeling with [35S]
Organ culture of condylar cartilage was according to Ogawa [9]. In short, explants of separated anlagen of condylar cartilage at E14. 0 were cultured on 0.1-mm Omnipore filters (Nihon Millipore, Tokyo, Japan) in Trowell-type organ culture containing BGJB medium with ascorbic acid (50 μM) and 1% (w/v) penicillin-streptomycin for 6 days in a humidified atmosphere of 5% CO2 in air at 37°C. In experimental groups, β-D-Xyloside (4-Methyumbelliferyl-β-D-xylopyranoside; Sigma, Saint Louis, MO, USA) was added to the culture media to a final concentration of 2mM. To confirm effects of β-D-Xyloside , [35S] Na2SO4 (100 μCi/ml) was added to culture media of explants at 6 days, and cultured for further 24 h under the same culture conditions. We analyzed three different dishes for each explant.
Extraction of proteoglycan and gel filtration
After labeling with [35S], the incubated medium was removed, and solid guanidine HCl and CHAPS were added to make the solutions 4 M in guanidine HCl and 0.5% (w/v) CHAPS (medium extracts). Explants were extracted with 4 M guanidine HCl, 50 mM sodium acetate (pH 6.0), containing 2% (w/v) Triton X-100 and 1% protease inhibitors (Sigma-Aldrich, Tokyo, Japan) at 4°C for 24 h (explant extracts). Both the medium and explant extracts were subjected to Sephadex G-50 columns (GE Healthcare, Buckinghamshire, UK) and eluted with 4 M guanidine HCI, 50 mM sodium acetate (pH 6.0), containing 0.5% (w/v) Triton X-100 to remove unincorporated radioactive precursors. The radioactivity in the collected void volume fractions was counted with a liquid scintillation counter and the total amount of [35S]-labeled macromolecules were estimated. [35S]-labeled macromolecules from Sephadex G-50 columns were then injected into prepacked Superose 6 columns (GE Healthcare) and eluted with 4 M guanidine HCl, 50 mM sodium acetate (pH 6.0), containing 0.5% (w/v) Triton X-100. Fractions (0.4 ml each) were collected and counted radioactivity.
Presenting graves were made with Excel 2010 (Windows) and all data were presented as mean ± standard error (SE). Standard significance was determined using one-way analysis of variance, followed by Mann-Whitney U-test. A P-value 0.05 was considered to indicate statistical significance.
Tissue preparation
After 6 days of culture, explants were fixed with 4% paraformaldehyde (0.1M phosphate buffer, pH7.4) at 4°C for 24 h, and processed for embedding in paraffin. Sections (5 μm) were made for general histology, in situ hybridization and immunohistochemistry. Other specimens were subjected to skeletal staining with alcian blue to confirm cartilage formation.
Digoxigenin-labelled RNA probes and in situ hybridization
The digoxigenin-labeled RNA probes used for versican, aggrecan and collagen types I and II were reported in our previous studies [12,13]. In situ hybridization was performed as previously described [13]. Briefly, deparaffinized sections were washed in TE (10 mM Tris- HCl, 1 mM EDTA, pH8.0), and then the sections were treated with 5 μg/ml protease K (Sigma Chemical, St Louis, MO, USA) for 15 min at 37°C. After washing in phosphate buffered-saline (PBS), the sections were treated with 0.2 M HCl for 10 min and then acetylated in 0.25% acetic anhydride in 0.1 M acetylation buffer (pH 8.0), dehydrated with a series of ethanol (70% to 100%), and air dried. The hybridization solution was applied to each section. Sense probes were used as negative controls. Hybridization was performed overnight at 50°C in a moist chamber. Each slide was washed in 2 × SSC containing 50% formamide at 60°C for 30 min. After washing in TNE (10 mM Tris- HCl, 1 mM, EDTA, 500 mM NaCl, pH7.6) at 37°C for 10 min, each slide was treated with 50 μg/ml RNAase in TNE at 37°C for 30 min, and washed in TNE. Subsequently, the slides were washed twice with 2 × SSC and 0.2 × SSC at 50°C for 20 min each. The DIG-labelled probes were detected using a Nucleic Acid Detection Kit (Roche Diagnostics) according to the manufacturer’s instructions.
Immunohistochemistry
A rabbit polyclonal antibody against versican - α domain was from GE Healthcare. Monoclonal antibodies against aggrecan (12/21/1C6, anti-reduced, alkylated aggrecan core protein) were obtained from Developmental Studies Hybridoma Bank (Iowa City, IA, USA). Rabbit polyclonal antibodies against collagen types I and II were from LSL (Tokyo, Japan). A HISTOFINE SAB kit (Nichirei, Tokyo, Japan) and a MOM kit (Vector Laboratories, Burlingame, CA, USA) were used for immunohistochemistry. After deparaffinization, sections were digested with testicular hyaluronidase (Sigma, St Louis, MO, USA; 25 mg/ml) for 60 min at 37°C. Sections for aggrecan were further immersed in mouse IgG blocking reagent in the MOM kit. All sections were then reacted with primary antibodies diluted with PBS containing 1% bovine serum albumin (1:100 for versican, 1:50 for aggrecan, 1:500 for collagen types I and II). The streptavidin–biotin method was then applied to the sections using the HISTOFINE SAB kit, as previously described [13]. Finally, sections were treated with 3-amino-9- ethylcarbazole (Nichirei, Tokyo, Japan) to reveal any reaction, and were examined by light microscopy after counterstaining with hematoxylin. Negative controls were incubated with normal anti-rabbit or antimouse IgG instead of the primary antibodies.
In the control groups after 6 days of culture, [35S]-labeled macromolecules were released both into explants and culture media and more than 80 % of them were into explants. Meanwhile, ~90% of them were released into culture media in β-D-xyloside treated experimental groups, although no standard significances were recognized between groups (Figure 1A). This may be due to small number of samples (n=3). Superose 6 chromatography indicated that in the control groups, a large peak (peak 1) eluted at the position of void volume was observed both in the medium and the explant extracts (Figure 1B). Peaks 1corresponds to aggrecan-type large proteoglycan [14]. In the experimental groups, peak 1 disappeared in the medium extracts and reduced markedly in the explant extracts. A new peak (peak 2) of smaller molecular weight was observed in the explant extracts. Peak 2 corresponds to β-D-xyloside-induced abnormal glycosaminoglycan chain [11]. These results confirmed that β-D-xyloside blocked normal PG synthesis.
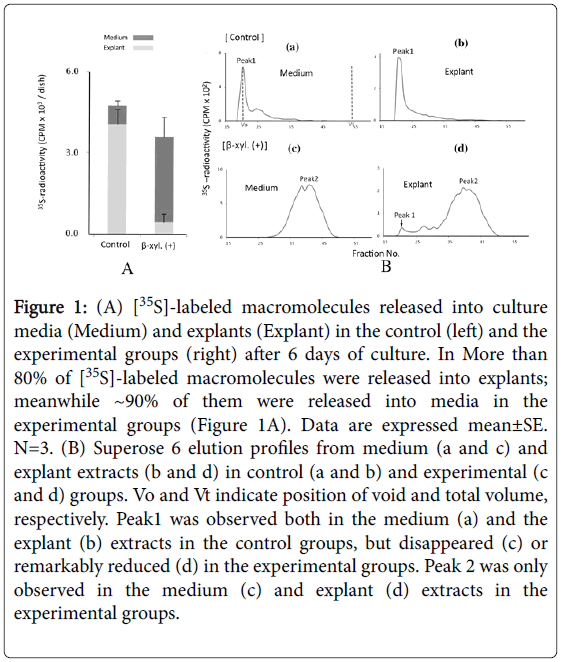
Figure 1: (A) [35S]-labeled macromolecules released into culture media (Medium) and explants (Explant) in the control (left) and the experimental groups (right) after 6 days of culture. In More than 80% of [35S]-labeled macromolecules were released into explants; meanwhile ~90% of them were released into media in the experimental groups (Figure 1A). Data are expressed mean±SE. N=3. (B) Superose 6 elution profiles from medium (a and c) and explant extracts (b and d) in control (a and b) and experimental (c and d) groups. Vo and Vt indicate position of void and total volume, respectively. Peak1 was observed both in the medium (a) and the explant (b) extracts in the control groups, but disappeared (c) or remarkably reduced (d) in the experimental groups. Peak 2 was only observed in the medium (c) and explant (d) extracts in the experimental groups.
Skeletal staining indicated that the condylar cartilage was clearly formed in the control groups, but its formation was hardly recognized in the experimental groups (Figure 2a and 2b).
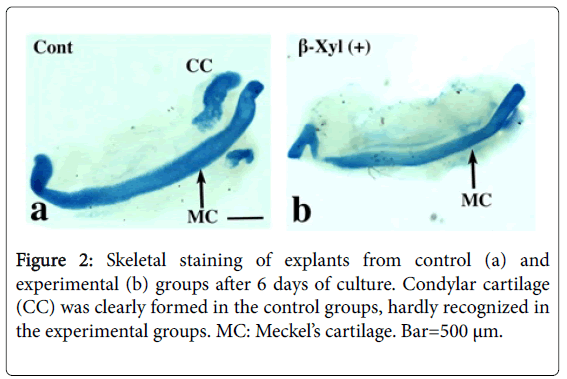
Figure 2: Skeletal staining of explants from control (a) and experimental (b) groups after 6 days of culture. Condylar cartilage (CC) was clearly formed in the control groups, hardly recognized in the experimental groups. MC: Meckel’s cartilage. Bar=500 μm.
Histological sections showed that oval-shaped condylar cartilage with thin perichondrium was clearly formed in the control groups. Cartilage matrix showed tissue metachromasia by toluidibe blue (Figure 3a) or stained with alcian blue (Figure 3b). In the experimental groups, condylar cartilage showing tissue metachromasia (Figure 3c) or stained with alcian blue (Figure 3d) was recognized, but cartilage matrix just occupied small area in the center and surrounded by thickened perichondrium (Figure 3c and 3d). Collagen type I mRNA and its immunoreactivity were mainly expressed in the perichondrium in the control groups (Figure 3e and 3f), but mainly within the cartilage matrix in the experimental groups (Figure 3g and 3h). Collagen type II mRNA and its immunoreactivity were expressed within the cartilage matrix, but expression areas in the experimental groups (Figure 3i and 3j) were much narrower than those in the control groups (Figure 3k and 3l). Aggrecan mRMA and its immunoreactivity showed similar expression patterns to those of collagen type II (Figure 3m-3p). Versican mRNA and its immunoreactivity were mainly expressed in the perichondrium, and the expression area of immunoreactivity in the experimental groups (Figure 3s and 3t) was wider than that in the control groups (Figure 3q and 3r).
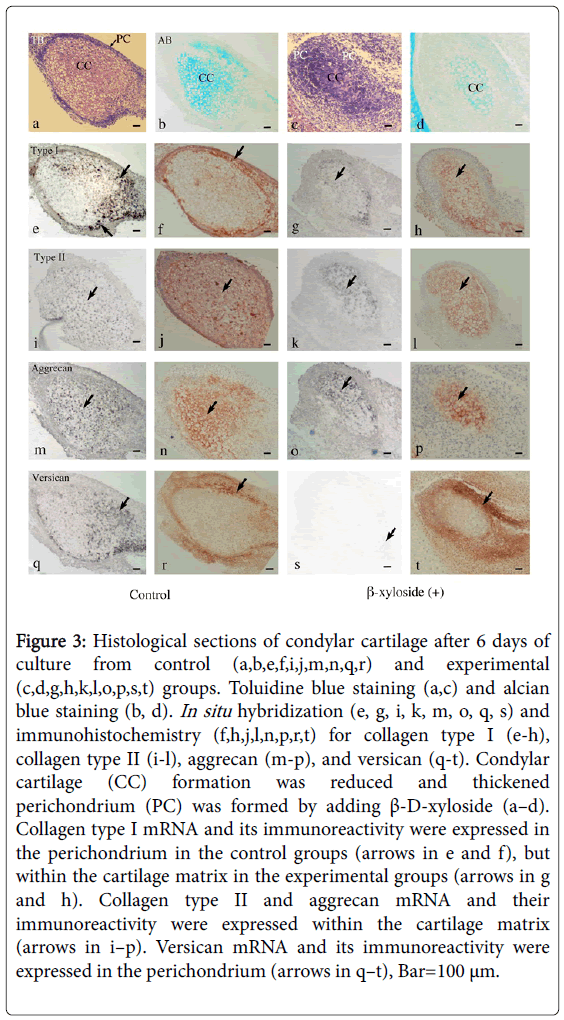
Figure 3: Histological sections of condylar cartilage after 6 days of culture from control (a,b,e,f,i,j,m,n,q,r) and experimental (c,d,g,h,k,l,o,p,s,t) groups. Toluidine blue staining (a,c) and alcian blue staining (b, d). In situ hybridization (e, g, i, k, m, o, q, s) and immunohistochemistry (f,h,j,l,n,p,r,t) for collagen type I (e-h), collagen type II (i-l), aggrecan (m-p), and versican (q-t). Condylar cartilage (CC) formation was reduced and thickened perichondrium (PC) was formed by adding β-D-xyloside (a–d). Collagen type I mRNA and its immunoreactivity were expressed in the perichondrium in the control groups (arrows in e and f), but within the cartilage matrix in the experimental groups (arrows in g and h). Collagen type II and aggrecan mRNA and their immunoreactivity were expressed within the cartilage matrix (arrows in i–p). Versican mRNA and its immunoreactivity were expressed in the perichondrium (arrows in q–t), Bar=100 μm.
In cartilage organ/cell culture systems, β-D-xyloside added into the culture media is rapidly cooperated into systems of CS synthesis, and abnormal CS chains are synthesized and secreted into the culture media [11]. Utilizing this nature, β-D-xyloside has been used to investigate functions of PGs in organogenesis such as salivary gland [15], lung [16], skeletal muscle [17], and tooth germ [18,19]. These studies indicate that normal organogenesis is impaired by adding β-Dxyloside. In normal mouse embryo, anlagen of mandibular condylar cartilage were recognized as mesenchymal condensation at E14.0, and newly formed condylar cartilage appears at E14.5-15.0 [4,7,12]. Previous organ cultures using condylar anlagen at E14.0 indicated that condylar cartilage forms by 6 days of culture [9,10], corresponding to present results in the control groups. In the experimental groups with β-D-xyloside, condylar cartilage formation was hardly detected by skeletal staining with alcian blue, indicating normal PG synthesis is required to form intact condylar cartilage. However, in histological sections, condylar cartilage formation was recognized, although its area was much smaller than that in the control groups. Profile of Superose 6 gel filtration indicated that peak 1 (corresponding to aggrecan-type large PG) was slightly recognized even in the experimental groups (Figure 1B), small amount of aggrecan molecules with normal GAG chains might be utilized for cartilage matrix. In addition, aggrecan and collagen type II mRNA expression was considerably maintained in the experimental groups, impaired formation of GAG chains rather than protein synthesis was responsible for the impaired formation of condylar cartilage. Type I collagen immunoreactivity was widely detected within the condylar cartilage in the experimental groups, indicating this cartilage has more fibrocartilaginous characteristics. Furthermore, the thickened perichondrium was compensatory formed around the cartilage matrix, and consequently the oval-shaped outline of condyle was considerably maintained in the experimental groups. The mandibular condylar cartilage is classified as the secondary cartilage and arises from the periostea of mandibular bone [3,4], and has fibrocartilaginous characteristics [6,7]. These natures might explain more fibrocartilaginous characteristics and the compensatory formation of thickened perichondrium to maintain the outline of condyle. These results further implied that, even under severe conditions lacking in PGs, mandibular condylar cartilage has ability to manage to execute its own morphogenesis using materials available.
Versican is non-cartilaginous large CSPG, and its normal formation may be blocked by β-D-xyloside. Strong versican immunoreactivity, however, was detected in the thickened perichondrium in the experimental groups, indicating, different from aggrecan, synthesis of versican may be enhanced. Quantitative analysis of versican /aggrecan synthesis is required to clarify this interesting topic in the future.
This work was supported by Grant-in-Aid for Scientific Research (No. 22592044 and 24792277) from Ministry of Education, Culture, Sports, Science, and Technology of Japan.